
Widely Targeted Metabolomics Reveals Metabolite Diversity in Jalapeño and Serrano Chile Peppers (Capsicum annuum L.)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Collection of Fruit Samples
2.2. UPLC-MS/MS Based Widely Targeted Metabolomics Approach
2.3. Qualitative and Quantitative Analyses of Metabolites
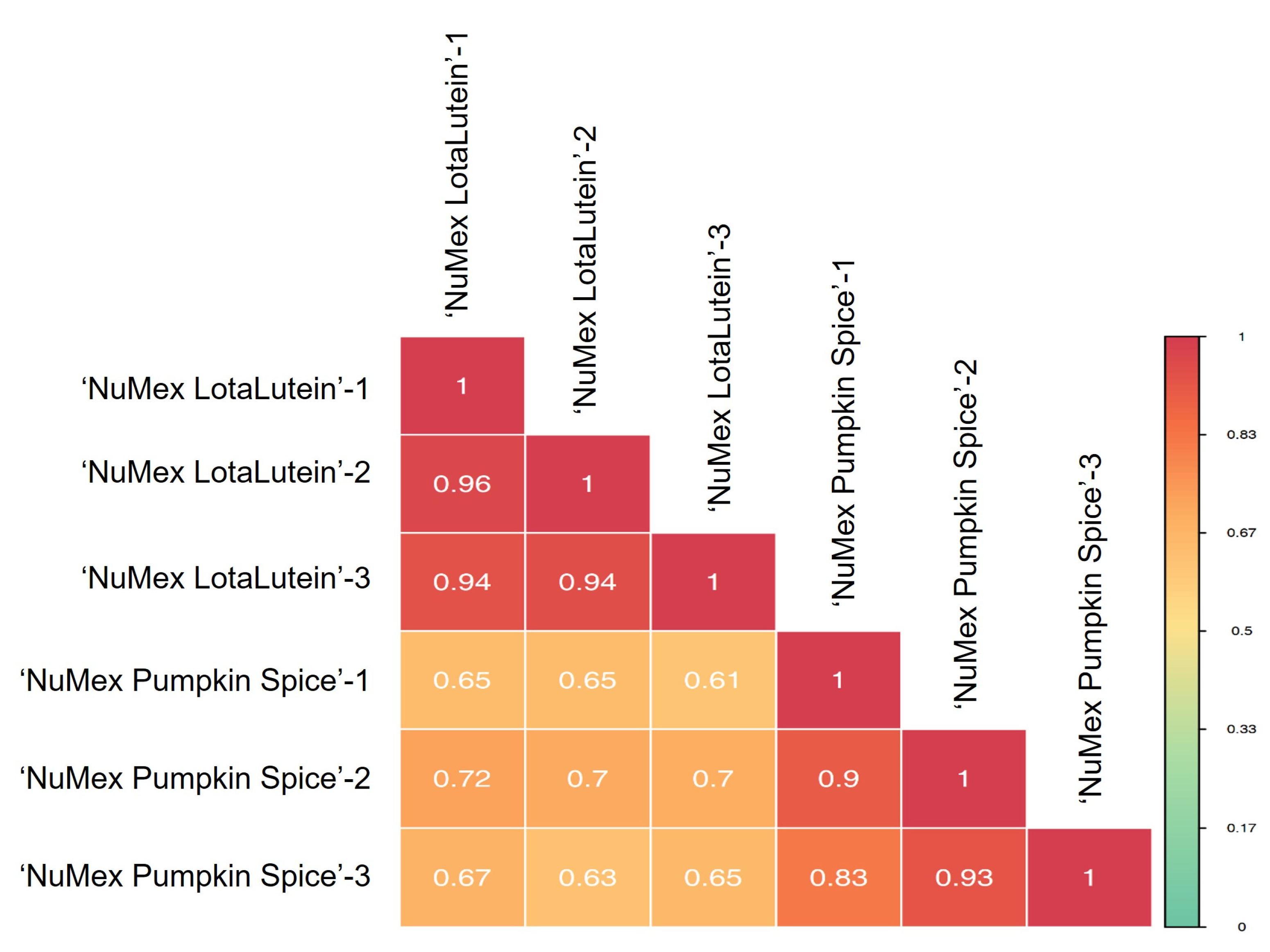
2.4. Sample Quality Control and Correlation Analysis
2.5. Analysis of Principal Components and Discriminant Analysis by Orthogonal Partial Least Squares
2.6. Identification and Functional Annotation of Differentially Expressed Metabolites
3. Results
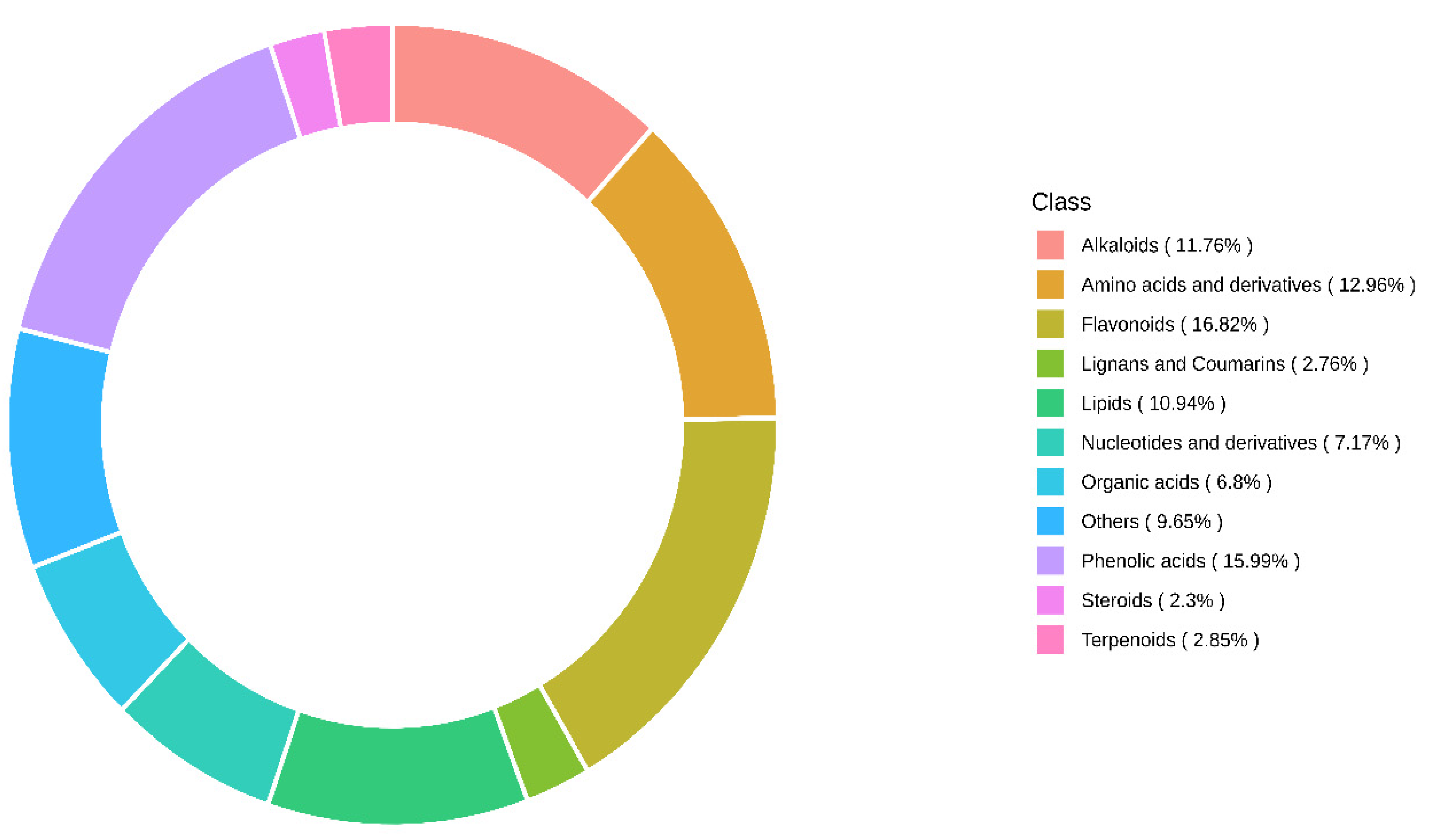
3.1. Fruit Sample Collection and Metabolite Identification
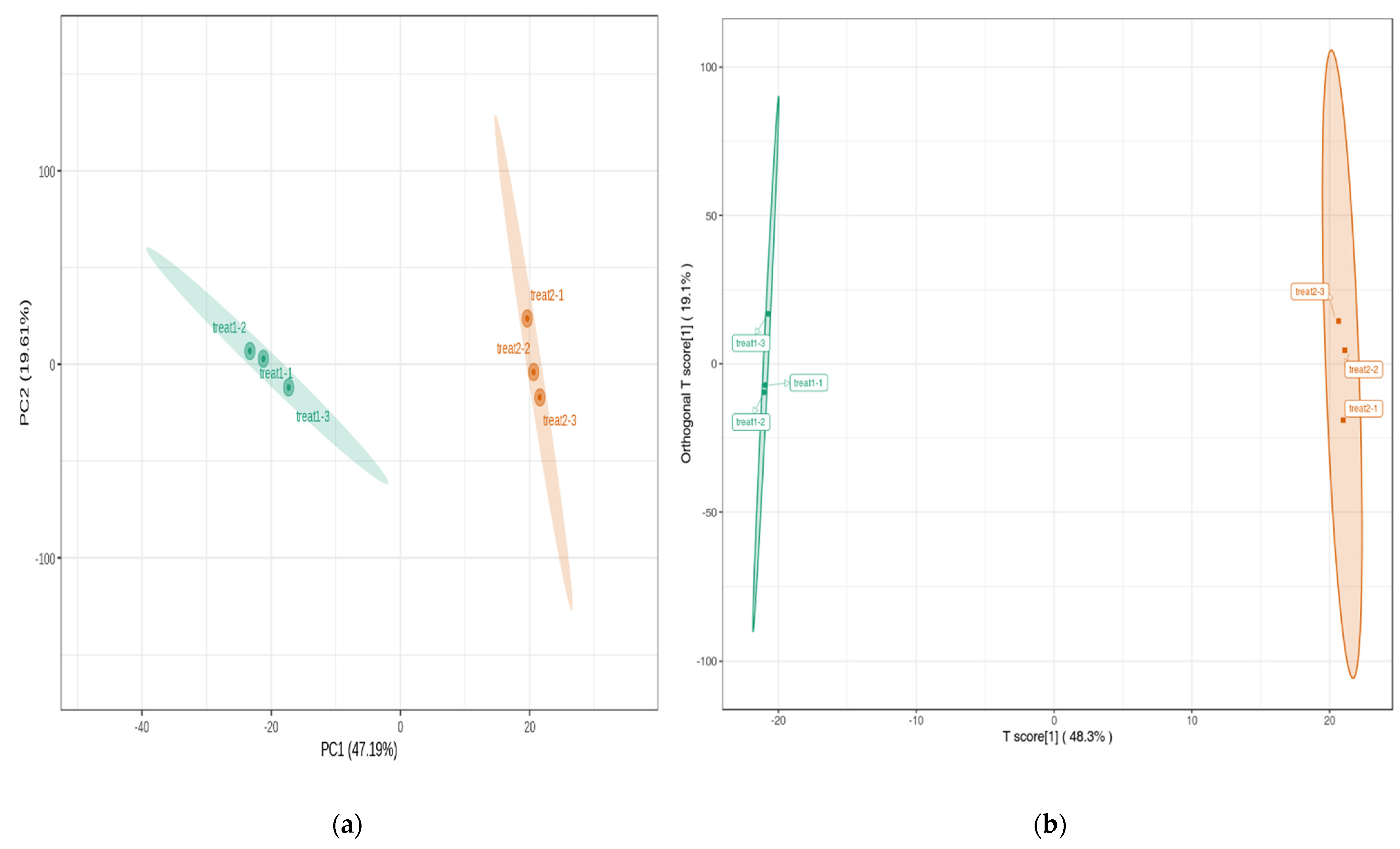
3.2. Analysis of Principal Components and Orthogonal Partial Least Squares
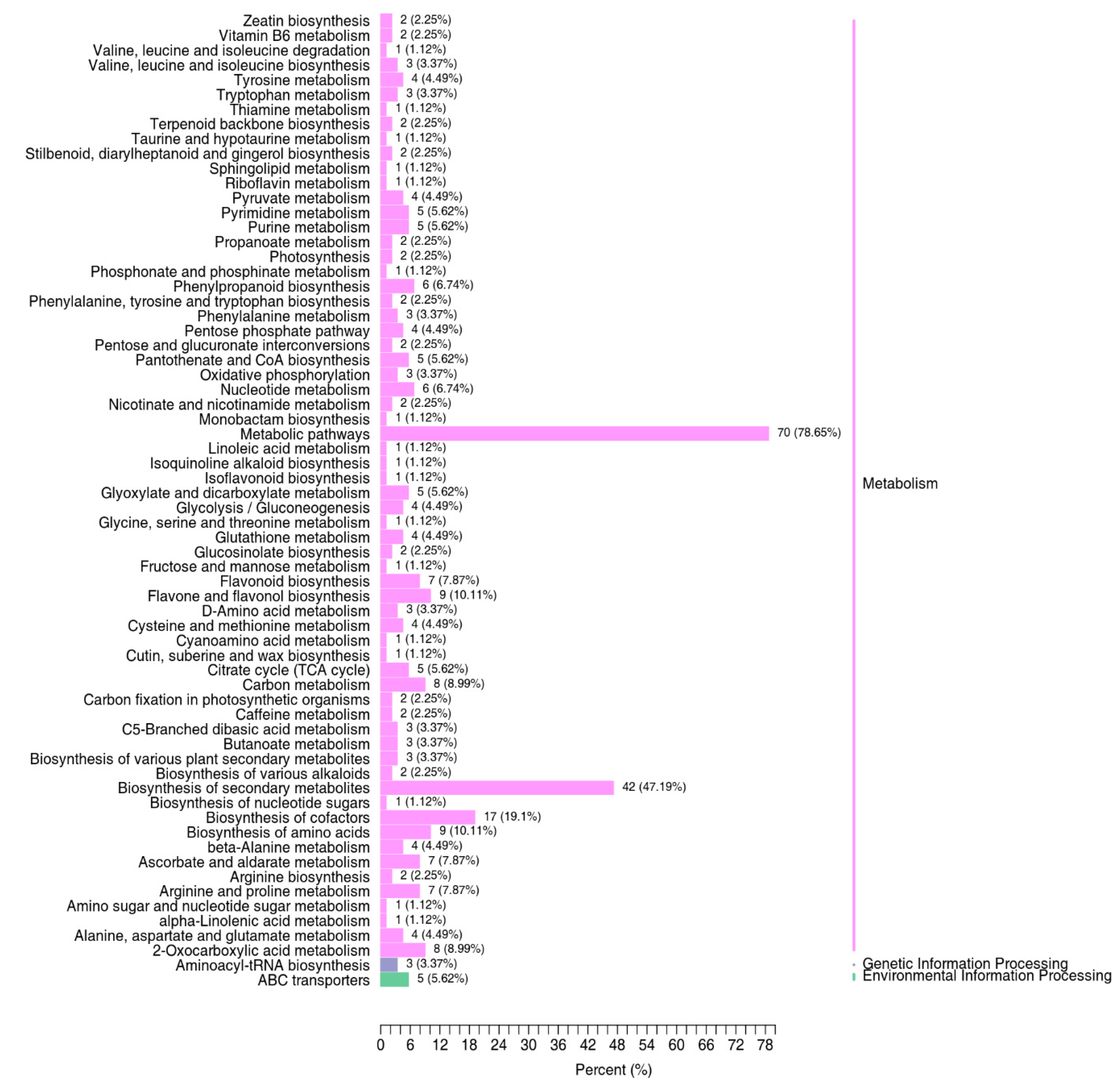
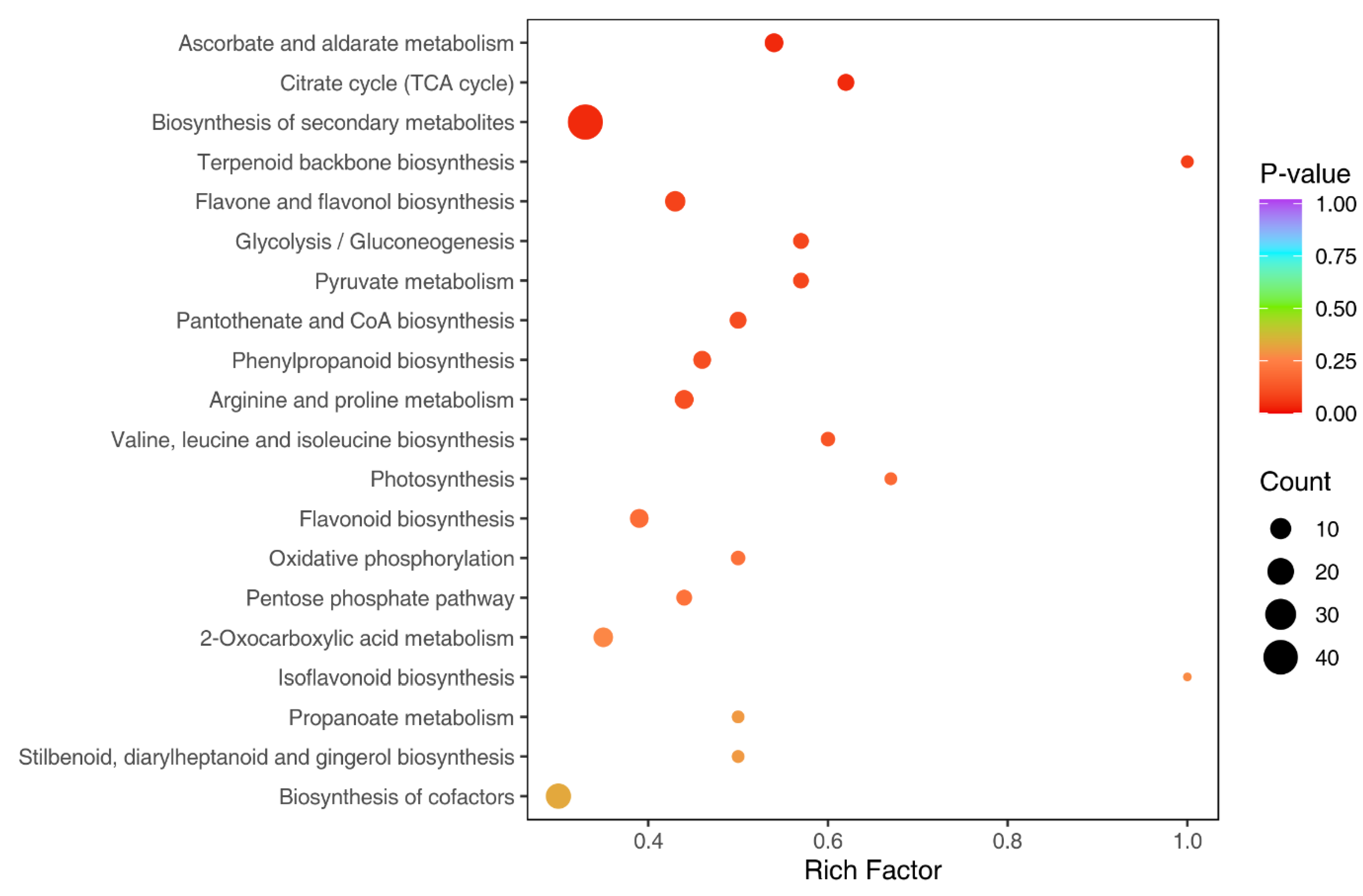
3.3. Identification and Functional Annotation of Differentially Expressed Metabolites
4. Discussion
4.1. Metabolite Profiles Successfully Discriminate Jalapeño and Serrano Type Chile Peppers
4.2. Differentially Expressed Metabolites Have Implications on Human Health and Nutrition
4.3. Integrating Metabolomics with Genomics Can Direct Breeding and Selection Decisions for Nutritional Quality Trait Improvement in Chile Pepper
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martínez-Ispizua, E.; Martínez-Cuenca, M.-R.; Marsal, J.I.; Díez, M.J.; Soler, S.; Valcárcel, J.V.; Calatayud, Á. Bioactive Compounds and Antioxidant Capacity of Valencian Pepper Landraces. Molecules 2021, 26, 1031. [Google Scholar] [CrossRef]
- Olatunji, T.L.; Afolayan, A.J. The suitability of chili pepper (Capsicum annuum L.) for alleviating human micronutrient dietary deficiencies: A review. Food Sci. Nutr. 2018, 6, 2239–2251. [Google Scholar] [CrossRef] [Green Version]
- Clark, R.; Lee, S.-H. Anticancer properties of capsaicin against human cancer. Anticancer Res. 2016, 36, 837–843. [Google Scholar]
- Batiha, G.E.-S.; Alqahtani, A.; Ojo, O.A.; Shaheen, H.M.; Wasef, L.; Elzeiny, M.; Ismail, M.; Shalaby, M.; Murata, T.; Zaragoza-Bastida, A. Biological properties, bioactive constituents, and pharmacokinetics of some Capsicum spp. and capsaicinoids. Int. J. Mol. Sci. 2020, 21, 5179. [Google Scholar] [CrossRef]
- Materska, M.; Perucka, I. Antioxidant activity of the main phenolic compounds isolated from hot pepper fruit (Capsicum annuum L.). J. Agric. Food Chem. 2005, 53, 1750–1756. [Google Scholar] [CrossRef]
- Sadef, Y.; Javed, T.; Javed, R.; Mahmood, A.; Alwahibi, M.S.; Elshikh, M.S.; AbdelGawwa, M.R.; Alhaji, J.H.; Rasheed, R.A. Nutritional status, antioxidant activity and total phenolic content of different fruits and vegetables’ peels. PLoS ONE 2022, 17, e0265566. [Google Scholar] [CrossRef]
- Lutz, M.; Hernández, J.; Henríquez, C. Phenolic content and antioxidant capacity in fresh and dry fruits and vegetables grown in Chile. CyTA-J. Food 2015, 13, 541–547. [Google Scholar]
- Taofiq, O.; González-Paramás, A.M.; Barreiro, M.F.; Ferreira, I.C.F.R. Hydroxycinnamic acids and their derivatives: Cosmeceutical significance, challenges and future perspectives, a review. Molecules 2017, 22, 281. [Google Scholar] [CrossRef]
- Kantar, M.B.; Anderson, J.E.; Lucht, S.A.; Mercer, K.; Bernau, V.; Case, K.A.; Le, N.C.; Frederiksen, M.K.; DeKeyser, H.C.; Wong, Z.-Z. Vitamin variation in Capsicum spp. provides opportunities to improve nutritional value of human diets. PLoS ONE 2016, 11, e0161464. [Google Scholar] [CrossRef]
- Luo, J. Metabolite-based genome-wide association studies in plants. Curr. Opin. Plant Biol. 2015, 24, 31–38. [Google Scholar] [CrossRef]
- Fiehn, O.; Kloska, S.; Altmann, T. Integrated studies on plant biology using multiparallel techniques. Curr. Opin. Biotechnol. 2001, 12, 82–86. [Google Scholar] [CrossRef]
- Taiti, C.; Costa, C.; Migliori, C.A.; Comparini, D.; Figorilli, S.; Mancuso, S. Correlation between volatile compounds and spiciness in domesticated and wild fresh chili peppers. Food Bioprocess Technol. 2019, 12, 1366–1380. [Google Scholar] [CrossRef]
- Barrajon-Catalan, E.; Álvarez-Martínez, F.J.; Borras, F.; Perez, D.; Herrero, N.; Ruiz, J.J.; Micol, V. Metabolomic analysis of the effects of a commercial complex biostimulant on pepper crops. Food Chem. 2020, 310, 125818. [Google Scholar] [CrossRef]
- Osorio, S.; Alba, R.; Nikoloski, Z.; Kochevenko, A.; Fernie, A.R.; Giovannoni, J.J. Integrative comparative analyses of transcript and metabolite profiles from pepper and tomato ripening and development stages uncovers species-specific patterns of network regulatory behavior. Plant Physiol. 2012, 159, 1713–1729. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Zhang, G.; Yang, T.; Yang, S.; Aisimutuola, P.; Wang, B.; Li, N.; Wang, J.; Yu, Q. Biochemical variances through metabolomic profile analysis of Jacq. during fruit development. Folia Hortic. 2021, 33, 17–26. [Google Scholar] [CrossRef]
- Feng, X.; Yu, Q.; Li, B.; Kan, J. Comparative analysis of carotenoids and metabolite characteristics in discolored red pepper and normal red pepper based on non-targeted metabolomics. LWT 2022, 153, 112398. [Google Scholar] [CrossRef]
- Cervantes-Hernández, F.; Ochoa-Alejo, N.; Martínez, O.; Ordaz-Ortiz, J.J. Metabolomic Analysis Identifies Differences Between Wild and Domesticated Chili Pepper Fruits During Development (Capsicum annuum L.). Front. Plant Sci. 2022, 13, 893055. [Google Scholar] [CrossRef]
- Zhou, Y.; Shao, L.; Zhu, J.; Li, H.; Duan, H. Comparative analysis of tuberous root metabolites between cultivated and wild varieties of Rehmannia glutinosa by widely targeted metabolomics. Sci. Rep. 2021, 11, 11460. [Google Scholar] [CrossRef]
- Adamski, J.; Suhre, K. Metabolomics platforms for genome wide association studies—Linking the genome to the metabolome. Curr. Opin. Biotechnol. 2013, 24, 39–47. [Google Scholar] [CrossRef]
- Sawada, Y.; Akiyama, K.; Sakata, A.; Kuwahara, A.; Otsuki, H.; Sakurai, T.; Saito, K.; Hirai, M.Y. Widely targeted metabolomics based on large-scale MS/MS data for elucidating metabolite accumulation patterns in plants. Plant Cell Physiol. 2009, 50, 37–47. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- Bosland, P.W.; Coon, D. ‘NuMex Lemon Spice’,‘NuMex Pumpkin Spice’, and ‘NuMex Orange Spice’Jalapenos. HortScience 2015, 50, 1104–1105. [Google Scholar] [CrossRef] [Green Version]
- Guzman, I.; Coon, D.; Vargas, K.; Bosland, P.W. NuMex LotaLutein, a Lutein-rich Serrano Pepper. HortScience 2020, 55, 2052–2055. [Google Scholar] [CrossRef]
- Fraga, C.G.; Clowers, B.H.; Moore, R.J.; Zink, E.M. Signature-discovery approach for sample matching of a nerve-agent precursor using liquid chromatography− mass spectrometry, XCMS, and chemometrics. Anal. Chem. 2010, 82, 4165–4173. [Google Scholar] [CrossRef]
- Bujak, R.; Daghir-Wojtkowiak, E.; Kaliszan, R.; Markuszewski, M.J. PLS-based and regularization-based methods for the selection of relevant variables in non-targeted metabolomics data. Front. Mol. Biosci. 2016, 3, 35. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Singh, D.P.; Bisen, M.S.; Shukla, R.; Prabha, R.; Maurya, S.; Reddy, Y.S.; Singh, P.M.; Rai, N.; Chaubey, T.; Chaturvedi, K.K. Metabolomics-Driven Mining of Metabolite Resources: Applications and Prospects for Improving Vegetable Crops. Int. J. Mol. Sci. 2022, 23, 12062. [Google Scholar] [CrossRef]
- Fiehn, O.; Kopka, J.; Dörmann, P.; Altmann, T.; Trethewey, R.N.; Willmitzer, L. Metabolite profiling for plant functional genomics. Nat. Biotechnol. 2000, 18, 1157–1161. [Google Scholar] [CrossRef]
- Ward, J.L.; Harris, C.; Lewis, J.; Beale, M.H. Assessment of 1H NMR spectroscopy and multivariate analysis as a technique for metabolite fingerprinting of Arabidopsis thaliana. Phytochemistry 2003, 62, 949–957. [Google Scholar] [CrossRef]
- Tohge, T.; Nishiyama, Y.; Hirai, M.Y.; Yano, M.; Nakajima, J.; Awazuhara, M.; Inoue, E.; Takahashi, H.; Goodenowe, D.B.; Kitayama, M. Functional genomics by integrated analysis of metabolome and transcriptome of Arabidopsis plants over-expressing an MYB transcription factor. Plant J. 2005, 42, 218–235. [Google Scholar] [CrossRef]
- Litvinov, D.Y.; Karlov, G.I.; Divashuk, M.G. Metabolomics for Crop Breeding: General Considerations. Genes 2021, 12, 1602. [Google Scholar] [CrossRef]
- Kim, E.-H.; Lee, K.M.; Lee, S.-Y.; Kil, M.; Kwon, O.-H.; Lee, S.-G.; Lee, S.-K.; Ryu, T.-H.; Oh, S.-W.; Park, S.-Y. Influence of genetic and environmental factors on the contents of carotenoids and phenolic acids in red pepper fruits (Capsicum annuum L.). Appl. Biol. Chem. 2021, 64, 1–11. [Google Scholar] [CrossRef]
- Lozada, D.N.; Bhatta, M.; Coon, D.; Bosland, P.W. Single Nucleotide Polymorphisms Reveal Genetic Diversity in New Mexican Chile Peppers (Capsicum spp.). BMC Genom. 2021, 22, 356. [Google Scholar] [CrossRef]
- Mi, S.; Yu, W.; Li, J.; Liu, M.; Sang, Y.; Wang, X. Characterization and discrimination of chilli peppers based on multi-element and non-targeted metabolomics analysis. LWT 2020, 131, 109742. [Google Scholar] [CrossRef]
- Espichán, F.; Rojas, R.; Quispe, F.; Cabanac, G.; Marti, G. Metabolomic characterization of 5 native Peruvian chili peppers (Capsicum spp.) as a tool for species discrimination. Food Chem. 2022, 386, 132704. [Google Scholar] [CrossRef]
- Florentino-Ramos, E.; Villa-Ruano, N.; Hidalgo-Martínez, D.; Ramírez-Meraz, M.; Méndez-Aguilar, R.; Velásquez-Valle, R.; Zepeda-Vallejo, L.G.; Pérez-Hernández, N.; Becerra-Martínez, E. 1H NMR-based fingerprinting of eleven Mexican Capsicum annuum cultivars. Food Res. Int. 2019, 121, 12–19. [Google Scholar] [CrossRef]
- Villa-Ruano, N.; Ramírez-Meraz, M.; Méndez-Aguilar, R.; Zepeda-Vallejo, L.G.; Álvarez-Bravo, A.; Pérez-Hernández, N.; Becerra-Martínez, E. 1H NMR-based metabolomics profiling of ten new races from Capsicum annuum cv. serrano produced in Mexico. Food Res. Int. 2019, 119, 785–792. [Google Scholar] [CrossRef]
- Lozada, D.N.; Coon, D.L.; Guzmán, I.; Bosland, P.W. Heat profiles of ‘superhot’and New Mexican type chile peppers (Capsicum spp.). Sci. Hortic. 2021, 283, 110088. [Google Scholar] [CrossRef]
- Carvalho Lemos, V.; Reimer, J.J.; Wormit, A. Color for life: Biosynthesis and distribution of phenolic compounds in pepper (Capsicum annuum). Agriculture 2019, 9, 81. [Google Scholar] [CrossRef] [Green Version]
- Kumar, O.A.; Rao, S.A.; Tata, S.S. Phenolics quantification in some genotypes of Capsicum annuum L. J. Phytol. 2010, 2, 87–90. [Google Scholar]
- Guzman, I.; Vargas, K.; Chacon, F.; McKenzie, C.; Bosland, P.W. Health-promoting carotenoids and phenolics in 31 Capsicum accessions. HortScience 2021, 56, 36–41. [Google Scholar] [CrossRef]
- De Sa Mendes, N.; Santos, M.C.P.; Santos, M.C.B.; Cameron, L.C.; Ferreira, M.S.L.; Goncalves, E.C.B.A. Characterization of pepper (Capsicum baccatum)—A potential functional ingredient. LWT 2019, 112, 108209. [Google Scholar] [CrossRef]
- Lee, Y.; Howard, L.R.; Villalon, B. Flavonoids and antioxidant activity of fresh pepper (Capsicum annuum) cultivars. J. Food Sci. 1995, 60, 473–476. [Google Scholar] [CrossRef]
- Chen, L.; Kang, Y.-H. Anti-inflammatory and antioxidant activities of red pepper (Capsicum annuum L.) stalk extracts: Comparison of pericarp and placenta extracts. J. Funct. Foods 2013, 5, 1724–1731. [Google Scholar] [CrossRef]
- Mokhtar, M.; Soukup, J.; Donato, P.; Cacciola, F.; Dugo, P.; Riazi, A.; Jandera, P.; Mondello, L. Determination of the polyphenolic content of a Capsicum annuum L. extract by liquid chromatography coupled to photodiode array and mass spectrometry detection and evaluation of its biological activity. J. Sep. Sci. 2015, 38, 171–178. [Google Scholar] [CrossRef]
- Jeong, W.Y.; Jin, J.S.; Cho, Y.A.; Lee, J.H.; Park, S.; Jeong, S.W.; Kim, Y.; Lim, C.; El-Aty, A.M.A.; Kim, G. Determination of polyphenols in three Capsicum annuum L. (bell pepper) varieties using high-performance liquid chromatography-tandem mass spectrometry: Their contribution to overall antioxidant and anticancer activity. J. Sep. Sci. 2011, 34, 2967–2974. [Google Scholar] [CrossRef]
- Rodrıguez-Concepción, M.; Boronat, A. Elucidation of the methylerythritol phosphate pathway for isoprenoid biosynthesis in bacteria and plastids. A metabolic milestone achieved through genomics. Plant Physiol. 2002, 130, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, M.; Vamvaka, E.; Torrado, A.; Jahns, P.; Dann, M.; Rosenhammer, L.; Aziba, A.; Leister, D.; Rühle, T. Introduction of the carotenoid biosynthesis α-branch into Synechocystis sp. PCC 6803 for lutein production. Front. Plant Sci. 2021, 12, 699424. [Google Scholar] [CrossRef]
- Zafar, J.; Aqeel, A.; Shah, F.I.; Ehsan, N.; Gohar, U.F.; Moga, M.A.; Festila, D.; Ciurea, C.; Irimie, M.; Chicea, R. Biochemical and Immunological implications of Lutein and Zeaxanthin. Int. J. Mol. Sci. 2021, 22, 10910. [Google Scholar] [CrossRef]
- Sakurai, N. Recent applications of metabolomics in plant breeding. Breed. Sci. 2022, 72, 56–65. [Google Scholar] [CrossRef]
- Lozada, D.N.; Bosland, P.; Barchenger, D.W.; Haghshenas-Jaryani, M.; Sanogo, S.; Walker, S. Chile Pepper (Capsicum) Breeding and Improvement in the “Multi-Omics” Era. Front. Plant Sci. 2022, 13, 879182. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Class | VIP 1 | Fold Change (FC) | log2FC 2 |
|---|---|---|---|---|
| 1-Phenylethanol | Phenolic acids | 1.44 | 269,754.80 | 18.04 |
| Dicaffeoylshikimic acid | Phenolic acids | 1.44 | 86,594.42 | 16.40 |
| 2-Methoxycinnamic acid | Phenolic acids | 1.44 | 10,264.53 | 13.33 |
| 1,3-O-Dicaffeoylquinic Acid (Cynarin) | Phenolic acids | 1.42 | 266.71 | 8.06 |
| 4-O-(4′-O-alpha-D-Glucopyranosyl)caffeoylquinic acid | Phenolic acids | 1.43 | 182.93 | 7.52 |
| Chlorogenic acid methyl ester | Phenolic acids | 1.44 | 103.94 | 6.70 |
| Cyanidin-3-O-(6′′-O-caffeoyl-2′′-O-xylosyl)glucoside | Flavonoids | 1.43 | 82.45 | 6.37 |
| Tri-O-galloyl Methyl gallate | Phenolic acids | 1.41 | 82.20 | 6.36 |
| Luteolin-7-O-Sophoroside-5-O-arabinoside | Flavonoids | 1.43 | 74.07 | 6.21 |
| 5-O-Feruloyl quinic acid glucoside | Phenolic acids | 1.37 | 73.70 | 6.20 |
| 3,6-Di-O-caffeoyl glucose | Phenolic acids | 1.44 | 9.18 × 10−6 | −16.73 |
| Rosmarinic acid methyl ester | Phenolic acids | 1.44 | 2.37 × 10−5 | −15.36 |
| 2,3-Dimethoxybenzaldehyde | Phenolic acids | 1.44 | 3.2 × 10−5 | −14.93 |
| Quercetin-3-O-(2′′-O-rhamnosyl)rutinoside-7-O-glucoside | Flavonoids | 1.44 | 2.5 × 10−4 | −11.96 |
| Quercetin-3-O-(6′′-O-acetyl)glucosyl-(1→3)-Galactoside | Flavonoids | 1.41 | 0.016 | −6.01 |
| p-Coumaroylagmatine | Alkaloids | 1.43 | 0.020 | −5.64 |
| Quercetin-7-O-(6′′-malonyl)glucosyl-5-O-glucoside | Flavonoids | 1.42 | 0.023 | −5.43 |
| Quercetin-3-O-rutinoside-7-O-rhamnoside | Flavonoids | 1.40 | 0.0251 | −5.31 |
| Quercetin-3-O-xylosyl(1→2)glucosyl(1→2)glucoside | Flavonoids | 1.40 | 0.0253 | −5.30 |
| Quercetin-3-O-(6′′-O-malonyl)glucosyl-5-O-glucoside | Flavonoids | 1.38 | 0.031 | −5.00 |
| Metabolite | Class | Variable Importance in Projection (VIP) Value | Fold Change (FC) | log2FC | KEGG Orthology (KO) Index | Annotation |
|---|---|---|---|---|---|---|
| Chlorogenic acid (3-O-Caffeoylquinic acid) | Phenolic acids | 1.43 | 20.46 | 4.35 | KO00940; KO00941; KO00945; KO01110 | Phenylpropanoid biosynthesis; flavonoid biosynthesis; stilbenoid, diarylheptanoid and gingerol biosynthesis; metabolic pathways |
| 3,7-Di-O-methylquercetin | Flavonoids | 1.34 | 11.21 | 3.49 | KO00944; KO01110 | Flavone and flavonol biosynthesis; metabolic pathways |
| Phenol | Phenolic acids | 1.42 | 11.01 | 3.46 | KO00350; KO01100 | Tyrosine metabolism; metabolic pathways |
| ATP; Adenosine 5′-Triphosphate | Nucleotides and derivatives | 1.42 | 8.39 | 3.07 | KO00190; KO00195; KO00230; KO00908 KO01100; KO01110; KO01232; KO01240 | Oxidative phosphorylation; photosynthesis; purine metabolism; zeatin biosynthesis; metabolic pathways; nucleotide metabolism; biosynthesis of cofactors |
| 2-Furoic acid | Organic acids | 1.38 | 5.77 | 2.53 | KO01100 | Metabolic pathways |
| Kaempferol-3-O-rutinoside(Nicotiflorin) | Flavonoids | 1.18 | 5.61 | 2.49 | KO00944; KO01110 | Flavone and flavonol biosynthesis; metabolic pathways |
| 2-Oxoheptanedionic acid | Organic acids | 1.37 | 5.55 | 2.47 | KO01100; KO01210; KO01240 | Metabolic pathways; 2-oxocarboxylic acid metabolism; biosynthesis of cofactors |
| Kaempferol-3-O-rhamnoside (Afzelin)(Kaempferin) | Flavonoids | 1.31 | 4.92 | 2.30 | KO00944 | Flavone and flavonol biosynthesis |
| Luteolin-7-O-glucuronide | Flavonoids | 1.26 | 4.76 | 2.25 | KO00944 | Flavone and flavonol biosynthesis |
| 3-O-Methylquercetin | Flavonoids | 1.40 | 4.56 | 2.19 | KO00944; KO01110 | Flavone and flavonol biosynthesis; metabolic pathways |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozada, D.N.; Pulicherla, S.R.; Holguin, F.O. Widely Targeted Metabolomics Reveals Metabolite Diversity in Jalapeño and Serrano Chile Peppers (Capsicum annuum L.). Metabolites 2023, 13, 288. https://doi.org/10.3390/metabo13020288
Lozada DN, Pulicherla SR, Holguin FO. Widely Targeted Metabolomics Reveals Metabolite Diversity in Jalapeño and Serrano Chile Peppers (Capsicum annuum L.). Metabolites. 2023; 13(2):288. https://doi.org/10.3390/metabo13020288
Chicago/Turabian StyleLozada, Dennis N., Sahithi Reddy Pulicherla, and Francisco Omar Holguin. 2023. "Widely Targeted Metabolomics Reveals Metabolite Diversity in Jalapeño and Serrano Chile Peppers (Capsicum annuum L.)" Metabolites 13, no. 2: 288. https://doi.org/10.3390/metabo13020288