
Chemical Characterization and Leishmanicidal Activity In Vitro and In Silico of Natural Products Obtained from Leaves of Vernonanthura brasiliana (L.) H. Rob (Asteraceae)
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of Crude Extracts and Fractions
2.3. Liquid Chromatography-Mass Spectrometry Analyses (LC-MS)
2.4. In Silico Studies and Molecular Docking
2.5. Parasites
2.6. Activity against Promastigote Forms
2.7. Fluorescence Microscopy
2.8. Determination of Drug Interactions
2.9. Cytotoxicity Assay in RAW 264.7 Cells
2.10. Toxicity in Tenebrio Molitor Larvae
2.11. Statistical Analyses
3. Results
3.1. Chromatography and Identification of Compounds
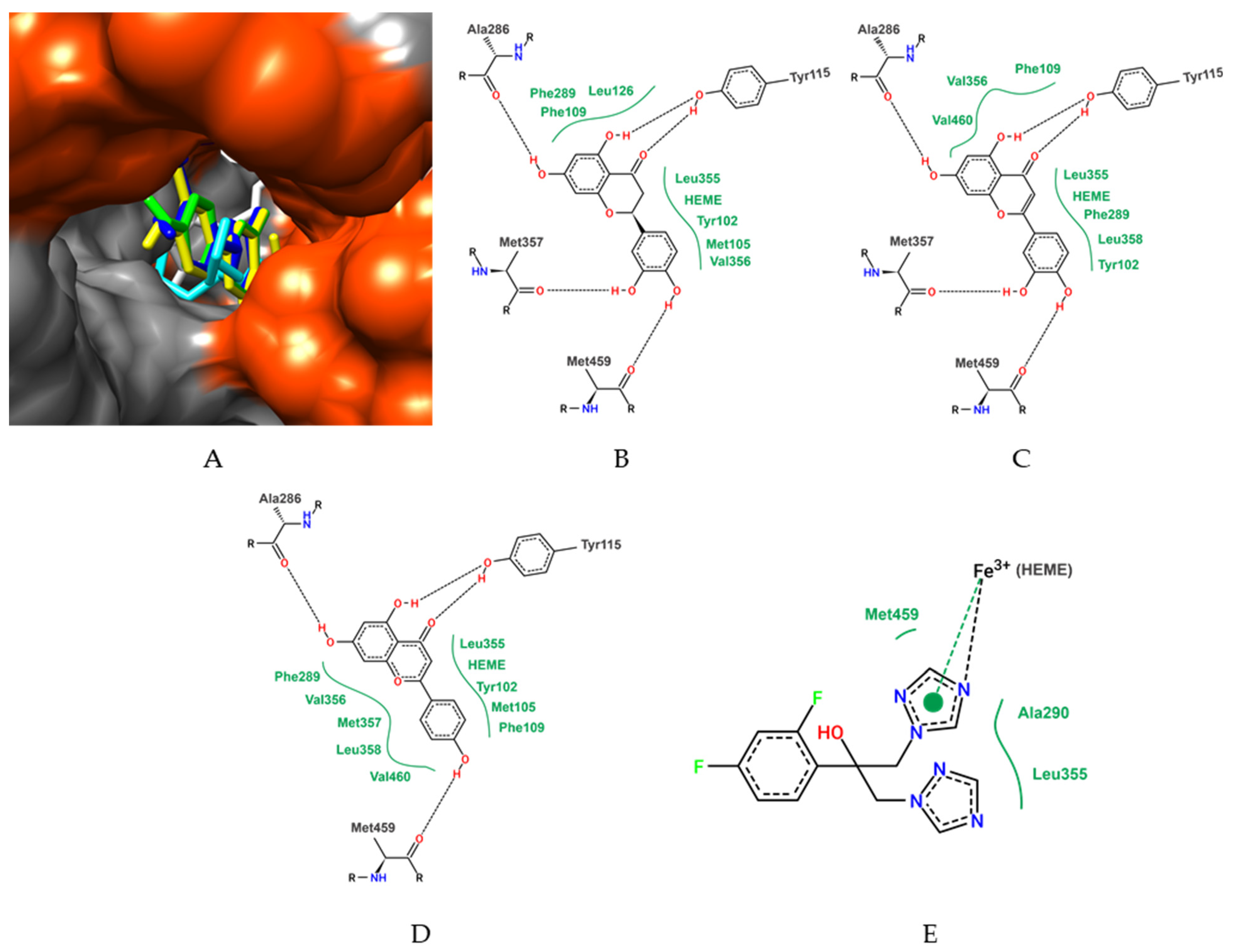
3.2. In Silico Studies and Molecular Docking
3.3. Leishmanicidal Activity
3.4. Determination of Drug Interactions
3.5. In Vitro Cytotoxicity
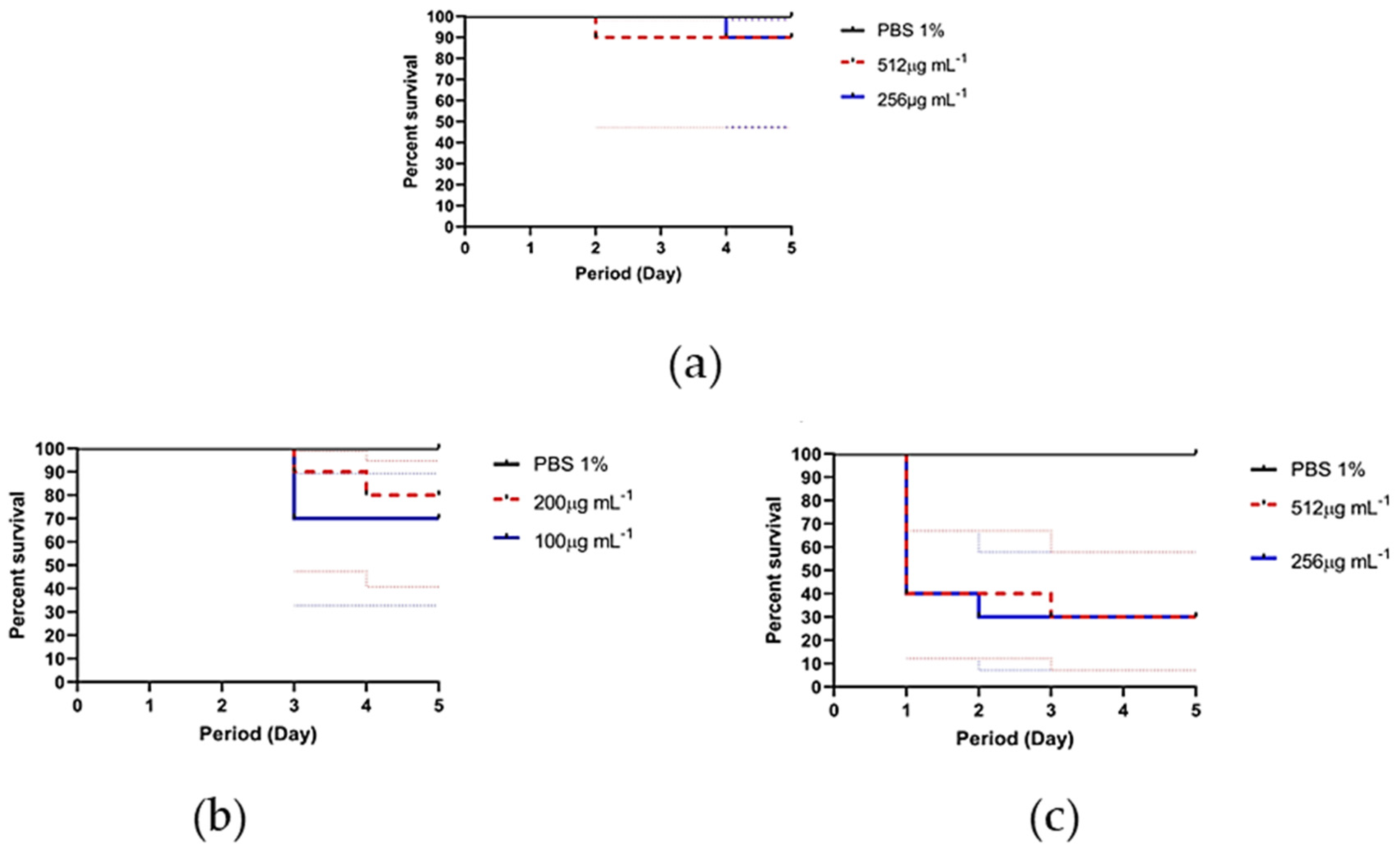
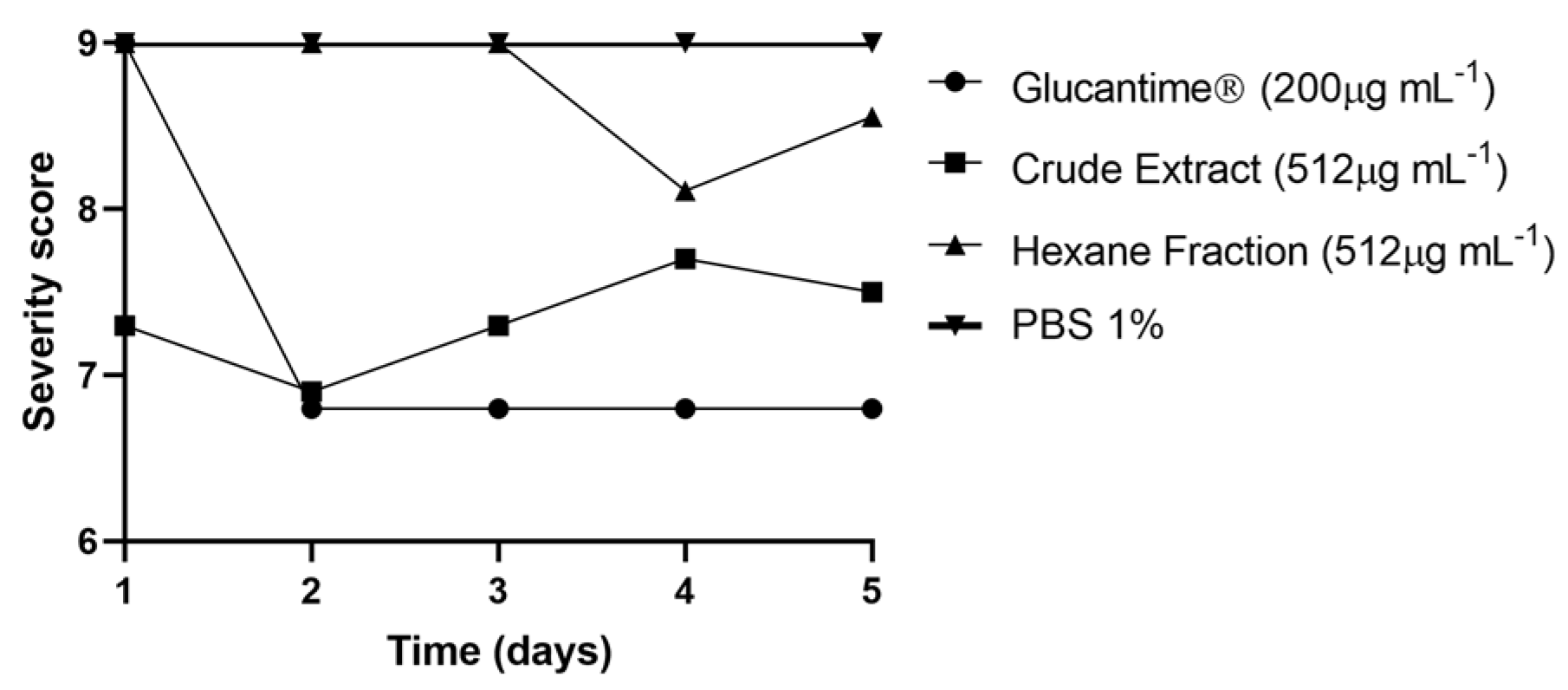
3.6. In Vivo Toxicity against Tenebrio Molitor
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mashayekhi-Ghoyonlo, V.; Kiafar, B.; Rohani, M.; Esmaeili, H.; Erfanian-Taghvaee, M.R. Correlation between Socioeconomic Status and Clinical Course in Patients with Cutaneous Leishmaniasis. J. Cutan. Med. Surg. 2015, 19, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M. Who Leishmaniasis Control the WHO Leishmaniasis Control Team Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef]
- Jennings, Y.L.; de Souza, A.A.A.; Ishikawa, E.A.; Shaw, J.; Lainson, R.; Silveira, F. Phenotypic characterization ofLeishmania spp. causing cutaneous leishmaniasis in the lower Amazon region, western Pará state, Brazil, reveals a putative hybrid parasite,Leishmania(Viannia)guyanensis × Leishmania(Viannia)shawi shawi. Parasite 2014, 21, 39. [Google Scholar] [CrossRef] [PubMed]
- Silveira, F.T.; Lainson, R.; Corbett, C.E.P. Clinical and immunopathological spectrum of American cutaneous leishmaniasis with special reference to the disease in Amazonian Brazil: A review. Mem. Inst. Oswaldo Cruz. 2004, 99, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Cock, I.; Selesho, M.; Van Vuuren, S. A review of the traditional use of southern African medicinal plants for the treatment of selected parasite infections affecting humans. J. Ethnopharmacol. 2018, 220, 250–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, L.; Almeida, J.; Macêdo, R.; Barbosa-Filho, J. A review of natural products with antileishmanial activity. Phytomedicine 2005, 12, 514–535. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Mishra, B.B.; Bajpai, S.; Singh, R.K.; Tiwari, V.K. Natural product based leads to fight against leishmaniasis. Bioorg. Med. Chem. 2013, 22, 18–45. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, L. Chinese herbal medicine: Fighting SARS-CoV-2 infection on all fronts. J. Ethnopharmacol. 2021, 270, 113869. [Google Scholar] [CrossRef]
- Abdallah, H.; El-Halawany, A.; Sirwi, A.; El-Araby, A.; Mohamed, G.; Ibrahim, S.; Koshak, A.; Asfour, H.; Awan, Z.; Elfaky, M.A. Repurposing of Some Natural Product Isolates as SARS-CoV-2 Main Protease Inhibitors via In Vitro Cell Free and Cell-Based Antiviral Assessments and Molecular Modeling Approaches. Pharmaceuticals 2021, 14, 213. [Google Scholar] [CrossRef]
- Huo, J.-L.; Fu, W.-J.; Liu, Z.-H.; Lu, N.; Jia, X.-Q.; Liu, Z.-S. Research advance of natural products in tumor immunotherapy. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Z. Natural Products, Alone or in Combination with FDA-Approved Drugs, to Treat COVID-19 and Lung Cancer. Biomedicines 2021, 9, 689. [Google Scholar] [CrossRef]
- Neto, R.N.M.; Setúbal, R.F.B.; Higino, T.M.M.; Castro, M.C.; Da Silva, L.C.N.; Aliança, A.S.D.S. Asteraceae Plants as Sources of Compounds Against Leishmaniasis and Chagas Disease. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef]
- Hameed, H.; King, E.; Doleckova, K.; Bartholomew, B.; Hollinshead, J.; Mbye, H.; Ullah, I.; Walker, K.; Van Veelen, M.; Abou-Akkada, S.; et al. Temperate Zone Plant Natural Products—A Novel Resource for Activity against Tropical Parasitic Diseases. Pharmaceuticals 2021, 14, 227. [Google Scholar] [CrossRef]
- de Sousa, D.F.; de Araújo, M.F.M.; de Mello, V.D.; Damasceno, M.M.C.; de Freitas, R.W.J.F. Cost-Effectiveness of Passion Fruit Albedo versus Turmeric in the Glycemic and Lipaemic Control of People with Type 2 Diabetes: Randomized Clinical Trial. J. Am. Coll. Nutr. 2020, 40, 679–688. [Google Scholar] [CrossRef]
- Sereno, A.B.; Pinto, C.D.; Andrade, F.A.; da Silva, M.A.B.; Garcia, A.C.; Krüger, C.C.H.; Reason, I.J.D.M. Effects of okra (Abelmoschus esculentus (L.) Moench) on glycemic markers in animal models of diabetes: A systematic review. J. Ethnopharmacol. 2022. [Google Scholar] [CrossRef]
- Li, S.-Z.; Zeng, S.-L.; Liu, E.-H. Anti-obesity natural products and gut microbiota. Food Res. Int. 2021, 151, 110819. [Google Scholar] [CrossRef]
- de Almeida, A.M.; Fonseca, C.R.; Prado, P.I.; Almeida-Neto, M.; Diniz, S.; Kubota, U.; Braun, M.R.; Raimundo, R.L.G.; dos Anjos, L.A.; Mendonça, T.G.; et al. Diversidade e ocorrência de Asteraceae em cerrados de São Paulo. Biota Neotrop. 2005, 5, 27–43. [Google Scholar] [CrossRef] [Green Version]
- De Mesquita, M.L.; Desrivot, J.; Bories, C.; Fournet, A.; De Paula, J.E.; Grellier, P.; Espindola, L. Antileishmanial and trypanocidal activity of Brazilian Cerrado plants. Mem. Inst. Oswaldo Cruz. 2005, 100, 783–787. [Google Scholar] [CrossRef]
- Nishimuta, H.; Rossi, A.; Yamashita, O.; Pena, G.; Santos, P.; Giustina, L.; Rossi, F. Leaf and Root Allelopathic Potential of the Vernonanthura brasiliana. Planta Daninha 2019, 37. [Google Scholar] [CrossRef]
- Toyang, N.J.; Verpoorte, R. A review of the medicinal potentials of plants of the genus Vernonia (Asteraceae). J. Ethnopharmacol. 2013, 146, 681–723. [Google Scholar] [CrossRef]
- Adedapo, A.A.; Aremu, O.J.; Oyagbemi, A. Anti-oxidant, anti-inflammatory and antinociceptive properties of the acetone leaf extract of vernonia amygdalina in some laboratory animals. Adv. Pharm. Bull. 2014, 4, 591. [Google Scholar] [CrossRef]
- Cáceres, A.L.; Flores-Giubi, M.E.; Romero-Rodríguez, M.C.; Alvarenga, N.L. In vitro anthelmintic activity and chemical composition of methanol extracts and fractions of Croton paraguayensis and Vernonia brasiliana against Eisenia fetida. Asian Pac. J. Trop. Dis. 2017, 7, 71–74. [Google Scholar] [CrossRef]
- de Arias, A.R.; Ferro, E.; Inchausti, A.; Ascurra, M.; Acosta, N.; Rodriguez, E.; Fournet, A. Mutagenicity, insecticidal and trypanocidal activity of some Paraguayan Asteraceae. J. Ethnopharmacol. 1995, 45, 35–41. [Google Scholar] [CrossRef]
- Rocha, M.F.G.; De Aguiar, F.L.N.; Brilhante, R.S.N.; Cordeiro, R.D.A.; Teixeira, C.E.C.; Castelo-Branco, D.D.S.C.M.; Paiva, M.D.A.N.; Zeferino, J.P.O.; Mafezoli, J.; Sampaio, C.M.D.S.; et al. Extratos de Moringa oleifera e Vernonia sp. sobre Candida albicans e Microsporum canis isolados de cães e gatos e análise da toxicidade em Artemia sp. Ciência Rural 2011, 41, 1807–1812. [Google Scholar] [CrossRef]
- da Silva, V.D.; Almeida-Souza, F.; Teles, A.M.; Neto, P.A.; Mondego-Oliveira, R.; Filho, N.E.M.; Taniwaki, N.N.; Abreu-Silva, A.L.; Calabrese, K.D.S.; Filho, V.E.M. Chemical composition of Ocimum canum Sims. essential oil and the antimicrobial, antiprotozoal and ultrastructural alterations it induces in Leishmania amazonensis promastigotes. Ind. Crop. Prod. 2018, 119, 201–208. [Google Scholar] [CrossRef]
- Maia, A.I.V.; Torres, M.C.M.; Pessoa, O.D.L.; De Menezes, J.E.S.A.; Costa, S.M.O.; Nogueira, V.L.R.; Melo, V.M.M.; De Souza, E.B.; Cavalcante, M.G.B.; Albuquerque, M.R.J.R. Óleos essenciais das folhas de Vernonia Remotiflora e Vernonia Brasiliana: Composição química e atividade biológica. Quim. Nova 2010, 33, 584–586. [Google Scholar] [CrossRef] [Green Version]
- Abreu, P.M.; Martins, E.S.; Kayser, O.; Bindseil, K.-U.; Siems, K.; Seemann, A.; Frevert, J. Antimicrobial, antitumor and antileishmania screening of medicinal plants from Guinea-Bissau. Phytomedicine 1999, 6, 187–195. [Google Scholar] [CrossRef]
- Mondêgo-Oliveira, R.; Sousa, J.C.D.S.; Moragas-Tellis, C.J.; de Souza, P.V.R.; Chagas, M.D.S.D.S.; Behrens, M.D.; Hardoim, D.D.J.; Taniwaki, N.N.; Chometon, T.Q.; Bertho, A.L.; et al. Vernonia brasiliana (L.) Druce induces ultrastructural changes and apoptosis-like death of Leishmania infantum promastigotes. Biomed. Pharmacother. 2021, 133, 111025. [Google Scholar] [CrossRef]
- Dennington, R.; Keith, T.A.; Millam, J.M. GaussView5. 2016. Available online: https://www.scirp.org/(S(vtj3fa45qm1ean45vvffcz55))/reference/ReferencesPapers.aspx?ReferenceID=1958990 (accessed on 1 December 2022).
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 09. 2016. Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkozje))/reference/referencespapers.aspx?referenceid=1989943 (accessed on 1 December 2022).
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.J.O.; Calado, G.P.; Fróes, Y.N.; de Araújo, S.A.; França, L.M.; Paes, A.M.D.A.; de Morais, S.V.; da Rocha, C.Q.; Vasconcelos, C.C. Plant Metabolites as SARS-CoV-2 Inhibitors Candidates: In Silico and In Vitro Studies. Pharmaceuticals 2022, 15, 1045. [Google Scholar] [CrossRef]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell Viability Assays. Assay Guid. Man. 2016. Available online: https://www.ncbi.nlm.nih.gov/books/NBK144065/ (accessed on 1 December 2022).
- Fivelman, Q.L.; Adagu, I.S.; Warhurst, D.C. Modified Fixed-Ratio Isobologram Method for Studying In Vitro Interactions between Atovaquone and Proguanil or Dihydroartemisinin against Drug-Resistant Strains of Plasmodium falciparum. Antimicrob. Agents Chemother. 2004, 48, 4097–4102. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Colasso, A.H.M.; Barros, T.F.; Figueiredo, I.F.D.S.; Junior, A.R.C.; Fernandes, E.S.; Uchoa, M.R.B.; da Silva, L.C.N. The latex of Euphorbia tirucalli inhibits staphyloxanthin production and protects Tenebrio molitor larvae against Staphylococcus aureus infection. Nat. Prod. Res. 2019, 34, 3536–3539. [Google Scholar] [CrossRef]
- Gobbo-Neto, L.; Lopes, N.P. Plantas Medicinais: Fatores de Influência No Conteúdo de Metabólitos Secundários. Quim. Nova 2007, 30, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Fugita, J.M.S.; Pereira, T.B.C.; Banzato, T.C.; Kitajima, E.W.; Souto, E.R.; Bedendo, I.P. Two distinct 16SrIII phytoplasma subgroups are associated with shoot proliferation in Vernonia brasiliana, a wild species inhabiting the Brazilian savanna. Trop. Plant Pathol. 2017, 42, 298–303. [Google Scholar] [CrossRef]
- Tuck, S.; Patel, H.; Safi, E.; Robinson, C. Lanosterol 14 alpha-demethylase (P45014DM): Effects of P45014DM inhibitors on sterol biosynthesis downstream of lanosterol. J. Lipid Res. 1991, 32, 893–902. [Google Scholar] [CrossRef]
- Sen, R.; Chatterjee, M. Plant derived therapeutics for the treatment of Leishmaniasis. Phytomedicine 2011, 18, 1056–1069. [Google Scholar] [CrossRef]
- Santos, G.C.d.O.; Vasconcelos, C.C.; Lopes, A.J.O.; do, S. de Sousa Cartágenes, M.; Filho, A.K.D.B.; do Nascimento, F.R.F.; Ramos, R.M.; Pires, E.R.R.B.; de Andrade, M.S.; Rocha, F.M.G.; et al. Candida Infections and Therapeutic Strategies: Mechanisms of Action for Traditional and Alternative Agents. Front. Microbiol. 2018, 9, 1351. [Google Scholar]
- Hargrove, T.Y.; Friggeri, L.; Wawrzak, Z.; Qi, A.; Hoekstra, W.J.; Schotzinger, R.J.; York, J.D.; Guengerich, F.P.; Lepesheva, G.I. Structural analyses of Candida albicans sterol 14α-demethylase complexed with azole drugs address the molecular basis of azole-mediated inhibition of fungal sterol biosynthesis. J. Biol. Chem. 2017, 292, 6728–6743. [Google Scholar] [CrossRef] [Green Version]
- Taran, M.; Mohebali, M.; Esmaeli, J. In Vivo Efficacy of Gum Obtained Pistacia Atlantica in Experimental Treatment of Cutaneous Leishmaniasis. Iran. J. Public Heal. 2010, 39, 36–41. [Google Scholar]
- Ahmed, Z.B.; Yousfi, M.; Viaene, J.; Dejaegher, B.; Demeyer, K.; Heyden, Y. Vander Four Pistacia Atlantica Subspecies (Atlantica, Cabulica, Kurdica and Mutica): A Review of Their Botany, Ethnobotany, Phytochemistry and Pharmacology. J. Ethnopharmacol. 2021, 265, 113329. [Google Scholar] [CrossRef]
- Blainski, A.; Gionco, B.; Oliveira, A.G.; Andrade, G.; Scarminio, I.S.; Silva, D.B.; Lopes, N.P.; Mello, J.C. Antibacterial activity of Limonium brasiliense (Baicuru) against multidrug-resistant bacteria using a statistical mixture design. J. Ethnopharmacol. 2017, 198, 313–323. [Google Scholar] [CrossRef]
- Salem, M.M.; Werbovetz, K.A. Antiprotozoal Compounds from Psorothamnus polydenius. J. Nat. Prod. 2004, 68, 108–111. [Google Scholar] [CrossRef]
- Sandjo, L.P.; de Moraes, M.H.; Kuete, V.; Kamdoum, B.C.; Ngadjui, B.T.; Steindel, M. Individual and combined antiparasitic effect of six plant metabolites against Leishmania amazonensis and Trypanosoma cruzi. Bioorg. Med. Chem. Lett. 2016, 26, 1772–1775. [Google Scholar] [CrossRef] [PubMed]
- DAS, B.B.; Sen, N.; Roy, A.; Dasgupta, S.B.; Ganguly, A.; Mohanta, B.C.; Dinda, B.; Majumder, H.K. Differential induction of Leishmania donovani bi-subunit topoisomerase I-DNA cleavage complex by selected flavones and camptothecin: Activity of flavones against camptothecin-resistant topoisomerase I. Nucleic Acids Res. 2006, 34, 1121–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittra, B.; Saha, A.; Chowdhury, A.R.; Pal, C.; Mandal, S.; Mukhopadhyay, S.; Bandyopadhyay, S.; Majumder, H.K. Luteolin, an Abundant Dietary Component is a Potent Anti-leishmanial Agent that Acts by Inducing Topoisomerase II-mediated Kinetoplast DNA Cleavage Leading to Apoptosis. Mol. Med. 2000, 6, 527–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manjolin, L.C.; dos Reis, M.B.G.; Maquiaveli, C.D.C.; Santos-Filho, O.A.; da Silva, E.R. Dietary flavonoids fisetin, luteolin and their derived compounds inhibit arginase, a central enzyme in Leishmania (Leishmania) amazonensis infection. Food Chem. 2013, 141, 2253–2262. [Google Scholar] [CrossRef] [Green Version]
- Albayrak, S.; Aksoy, A.; Sağdiç, O.; Budak, Ü. Phenolic compounds and antioxidant and antimicrobial properties of Helichrysum species collected from eastern Anatolia, Turkey. Turk. J. Biol. 2010, 34, 463–473. [Google Scholar] [CrossRef]
- Morikawa, T.; Ninomiya, K.; Akaki, J.; Kakihara, N.; Kuramoto, H.; Matsumoto, Y.; Hayakawa, T.; Muraoka, O.; Wang, L.-B.; Wu, L.-J.; et al. Dipeptidyl peptidase-IV inhibitory activity of dimeric dihydrochalcone glycosides from flowers of Helichrysum arenarium. J. Nat. Med. 2015, 69, 494–506. [Google Scholar] [CrossRef] [Green Version]
- Tabatabaei, S.M.; Farimani, M.M.; Nejad-Ebrahimi, S.; Salehi, P. Phytochemical study of Tanacetum sonbolii aerial parts and the antiprotozoal activity of its components. Biointerface Res. Appl. Chem. 2020, 19, 77–83. [Google Scholar] [CrossRef]
- Czinner, E.; Hagymási, K.; Blázovics, A.; Kéry, A.; Szőke, E.; Lemberkovics, E. In vitro antioxidant properties of Helichrysum arenarium (L.) Moench. J. Ethnopharmacol. 2000, 73, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Kefi, S.; Essid, R.; Mkadmini, K.; Kefi, A.; Haddada, F.M.; Tabbene, O.; Limam, F. Phytochemical investigation and biological activities of Echium arenarium (Guss) extracts. Microb. Pathog. 2018, 118, 202–210. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, D.P.; de Almeida, L.; Marques, M.J.; de Carvalho, R.R.; Dias, A.L.T.; da Silva, G.A.; de Pádua, R.M.; Braga, F.C.; da Silva, M.A. Exploring the bioactivity potential of Leonotis nepetifolia: Phytochemical composition, antimicrobial and antileishmanial activities of extracts from different anatomical parts. Nat. Prod. Res. 2019, 35, 3120–3125. [Google Scholar] [CrossRef]
- Fonseca-Silva, F.; Canto-Cavalheiro, M.M.; Menna-Barreto, R.F.S.; Almeida-Amaral, E.E. Effect of Apigenin on Leishmania amazonensis Is Associated with Reactive Oxygen Species Production Followed by Mitochondrial Dysfunction. J. Nat. Prod. 2015, 78, 880–884. [Google Scholar] [CrossRef]
- Naddaf, N.; Haddad, S. Apigenin effect against Leishmania tropica amastigotes in vitro. J. Parasit. Dis. 2020, 44, 574–578. [Google Scholar] [CrossRef]
- Cruz, E.D.M.; da Silva, E.R.; Maquiaveli, C.D.C.; Alves, E.S.S.; Lucon, J.F.; dos Reis, M.B.G.; de Toledo, C.E.M.; Cruz, F.G.; Vannier-Santos, M.A. Leishmanicidal activity of Cecropia pachystachya flavonoids: Arginase inhibition and altered mitochondrial DNA arrangement. Phytochemistry 2013, 89, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Emiliano, Y.S.S.; Almeida-Amaral, E.E. Efficacy of Apigenin and Miltefosine Combination Therapy against Experimental Cutaneous Leishmaniasis. J. Nat. Prod. 2018, 81, 1910–1913. [Google Scholar] [CrossRef]
- Gonçalves-Oliveira, L.F.; Souza-Silva, F.; Côrtes, L.M.D.C.; Veloso, L.B.; Pereira, B.A.S.; Cysne-Finkelstein, L.; Lechuga, G.C.; Bourguignon, S.C.; Almeida-Souza, F.; Calabrese, K.D.S.; et al. The combination therapy of meglumine antimoniate and oxiranes (epoxy-α-lapachone and epoxymethyl-lawsone) enhance the leishmanicidal effect in mice infected by Leishmania (Leishmania) amazonensis. Int. J. Parasitol. Drugs Drug Resist. 2019, 10, 101–108. [Google Scholar] [CrossRef]
- Pastor, J.; García, M.; Steinbauer, S.; Setzer, W.N.; Scull, R.; Gille, L.; Monzote, L. Combinations of ascaridole, carvacrol, and caryophyllene oxide against Leishmania. Acta Trop. 2015, 145, 31–38. [Google Scholar] [CrossRef]
- dos Santos, D.B.; Lemos, J.A.; Miranda, S.E.M.; Di Filippo, L.D.; Duarte, J.L.; Ferreira, L.A.M.; Barros, A.L.B.; Oliveira, A.E.M.F.M. Current Applications of Plant-Based Drug Delivery Nano Systems for Leishmaniasis Treatment. Pharmaceutics 2022, 14, 2339. [Google Scholar] [CrossRef] [PubMed]
- Mehrizi, T.Z.; Khamesipour, A.; Ardestani, M.S.; Shahmabadi, H.E.; Hoseini, M.H.M.; Mosaffa, N.; Ramezani, A. Comparative analysis between four model nanoformulations of amphotericin B-chitosan, amphotericin B-dendrimer, betulinic acid-chitosan and betulinic acid-dendrimer for treatment of Leishmania major: Real-time PCR assay plus. Int. J. Nanomed. 2019, ume 14, 7593–7607. [Google Scholar] [CrossRef] [Green Version]
- Mehrizi, T.Z.; Ardestani, M.S.; Hoseini, M.H.M.; Khamesipour, A.; Mosaffa, N.; Ramezani, A. Novel Nanosized Chitosan-Betulinic Acid Against Resistant Leishmania Major and First Clinical Observation of such parasite in Kidney. Sci. Rep. 2018, 8, 1–19. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Retention Time (min) | [M-H] - | MS n Ion m/z (-) | Proposed Substance |
|---|---|---|---|---|
| 1 | 16.2 | 353; 191 | 353 | chlorogenic acid or 5-o-caffeoylquinic acid |
| 2 | 16.7 | 353; 289; 203 | 515 | 3,5-dicapheoylquinic acid |
| 3 | 22.2 | 287; 151 | 463 | eriodictyol-7-o-glucuronide |
| 4 | 23.4 | 375; 191 | 537 | hydroxylariciresinol-hexoside |
| 5 | 24.0 | 149 | 177 | esculetin |
| 6 | 25.4 | 285 | 923/461 | luteolin-7-o-glucuronide |
| 7 | 27.2 | 151 | 575/287 | eriodictyol |
| 8 | 28.9 | 275; 231 | 293 | octadecadienoic acid |
| 9 | 30.9 | 285 | 571 | luteolin |
| 10 | 32.9 | 269; 151 | 539/269 | apigenin |
| 11 | 33.1 | 284 | 299 | hispidulin |
| 12 | 35.5 | 299 | 599 | chrysoeriol |
| 13 | 37.7 | 298; 283 | 313 | 3’,4’-dimethoxy luteolin |
| 14 | 41.6 | 171 | 293 | gingerol |
| Leishmania infantum CYP51 | |
|---|---|
| Ligand | ΔGbind (kcal/mol) |
| eriodictyol | −9.0 |
| luteolin | −8.7 |
| apigenin | −8.6 |
| hispidulin | −8.5 |
| chlorogenic acid | −8.1 |
| chrysoeriol | −8.0 |
| 3’,4’-dimethoxyluteolin | −8.0 |
| gingerol | −7.3 |
| luteolin-7-o-glucuronide | −7.2 |
| eriodictiol-7-o-glucuronide | −7.1 |
| esculetin | −6.9 |
| quinic acid | −6.9 |
| hydroxylariciresinol-hexoside | −6.8 |
| Fluconazole | −8.1 |
| Natural Products | 24 h | 48 h | 72 h | |||
|---|---|---|---|---|---|---|
| IC50 ± SD (µg·mL−1) | R² | IC50 ± SD (µg·mL−1) | R² | IC50 ± SD (µg·mL−1) | R² | |
| EBVb | 24.63 ± 0.375 a,1 | 0.970 | 24.31 ± 0.625 a,1 | 0.904 | 19.04 ± 0.750 b,1 | 0.880 |
| FHVb | 5.76 ± 0.250 a,2 | 0.972 | 12.44 ± 0.875 b,2 | 0.941 | 22.53 ± 0.125 c,2 | 0.876 |
| FAEVb | 21.78 ± 0.500 a,3 | 0.947 | 28.24 ± 0.750 b,3 | 0.953 | 54.89 ± 0.375 c,3 | 0.879 |
| FMVb | 60.63 ± 0.250 a,4 | 0.940 | 353.3 ± 1.00 b,4 | 0.948 | 60.13 ± 0.125 c,4 | 0.862 |
| Glucantime® | >500 | ND | >500 | ND | >500 | ND |
| Amphotericin-B | 0.1644 ± 0.500 a | 0.996 | 0.5388 ± 0.250 a | 0.991 | 0.7793 ± 0.375 a | 0.991 |
| Combination Ratio | Combined Drugs | FICI | |||||
|---|---|---|---|---|---|---|---|
| IC50 µg·mL−1 | FIC50 | ∑FIC50 | |||||
| FHVb | Anf-B | FHVb | Anf-B | FHVb | Anf-B | ||
| 5 | 0 | 160 | 0 | 7.101 | 0 | 7.101 | 23,691 |
| 4 | 1 | 80 | 0.25 | 3550 | 0.320 | 3.870 | |
| 3 | 2 | 40 | 0.5 | 1775 | 0.641 | 2416 | |
| 2 | 3 | 20 | 1 | 0.887 | 1283 | 2.170 | |
| 1 | 4 | 10 | 2 | 0.443 | 2567 | 3.010 | |
| 0 | 5 | 0 | 4 | 0 | 5134 | 5124 | |
| Natural Compounds | CC50 (µg·mL−1) | 95% CI (µg·mL−1) | R² | SI |
|---|---|---|---|---|
| EBVb | <8 | ND | ND | ND |
| FHVb | 314.8 | 90.23–106.6 | 0.964 | 25.30 |
| FAEVb | 32.4 | 95.60–147.4 | 0.979 | 1, 14 |
| DMSO (40µg·mL−1) | <1 | ND | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fróes, Y.N.; Araújo, J.G.N.; Gonçalves, J.R.d.S.; Oliveira, M.d.J.M.G.d.; Everton, G.O.; Filho, V.E.M.; Silva, M.R.C.; Silva, L.D.M.; Silva, L.A.; Neto, L.G.L.; et al. Chemical Characterization and Leishmanicidal Activity In Vitro and In Silico of Natural Products Obtained from Leaves of Vernonanthura brasiliana (L.) H. Rob (Asteraceae). Metabolites 2023, 13, 285. https://doi.org/10.3390/metabo13020285
Fróes YN, Araújo JGN, Gonçalves JRdS, Oliveira MdJMGd, Everton GO, Filho VEM, Silva MRC, Silva LDM, Silva LA, Neto LGL, et al. Chemical Characterization and Leishmanicidal Activity In Vitro and In Silico of Natural Products Obtained from Leaves of Vernonanthura brasiliana (L.) H. Rob (Asteraceae). Metabolites. 2023; 13(2):285. https://doi.org/10.3390/metabo13020285
Chicago/Turabian StyleFróes, Yuri Nascimento, João Guilherme Nantes Araújo, Joyce Resende dos Santos Gonçalves, Milena de Jesus Marinho Garcia de Oliveira, Gustavo Oliveira Everton, Victor Elias Mouchrek Filho, Maria Raimunda Chagas Silva, Luís Douglas Miranda Silva, Lucilene Amorim Silva, Lídio Gonçalves Lima Neto, and et al. 2023. "Chemical Characterization and Leishmanicidal Activity In Vitro and In Silico of Natural Products Obtained from Leaves of Vernonanthura brasiliana (L.) H. Rob (Asteraceae)" Metabolites 13, no. 2: 285. https://doi.org/10.3390/metabo13020285