
Transcriptome and Metabolome Analysis Reveal the Flavonoid Biosynthesis Mechanism of Abelmoschus manihot L. at Different Anthesis Stages
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Determination of Total Flavonoid Content
2.3. Vitexin Rhamnoside, Hyperin, Rutin, and Quercetin Content Measurement in HSK
2.4. Antioxidant Activity of Total Flavonoids from HSK
2.5. Metabolome Detection and Analysis by LC-MS/MS
2.6. Transcriptome Detection and Analysis in HSK
2.7. RT-PCR Detection
3. Results and Discussion
3.1. Phenotype and Total Flavonoids Measurement in HSK
3.2. Several Main Flavonoids Detection in HSK Flowers
3.3. Antioxidant Activity Analysis of Total Flavonoids in HSK
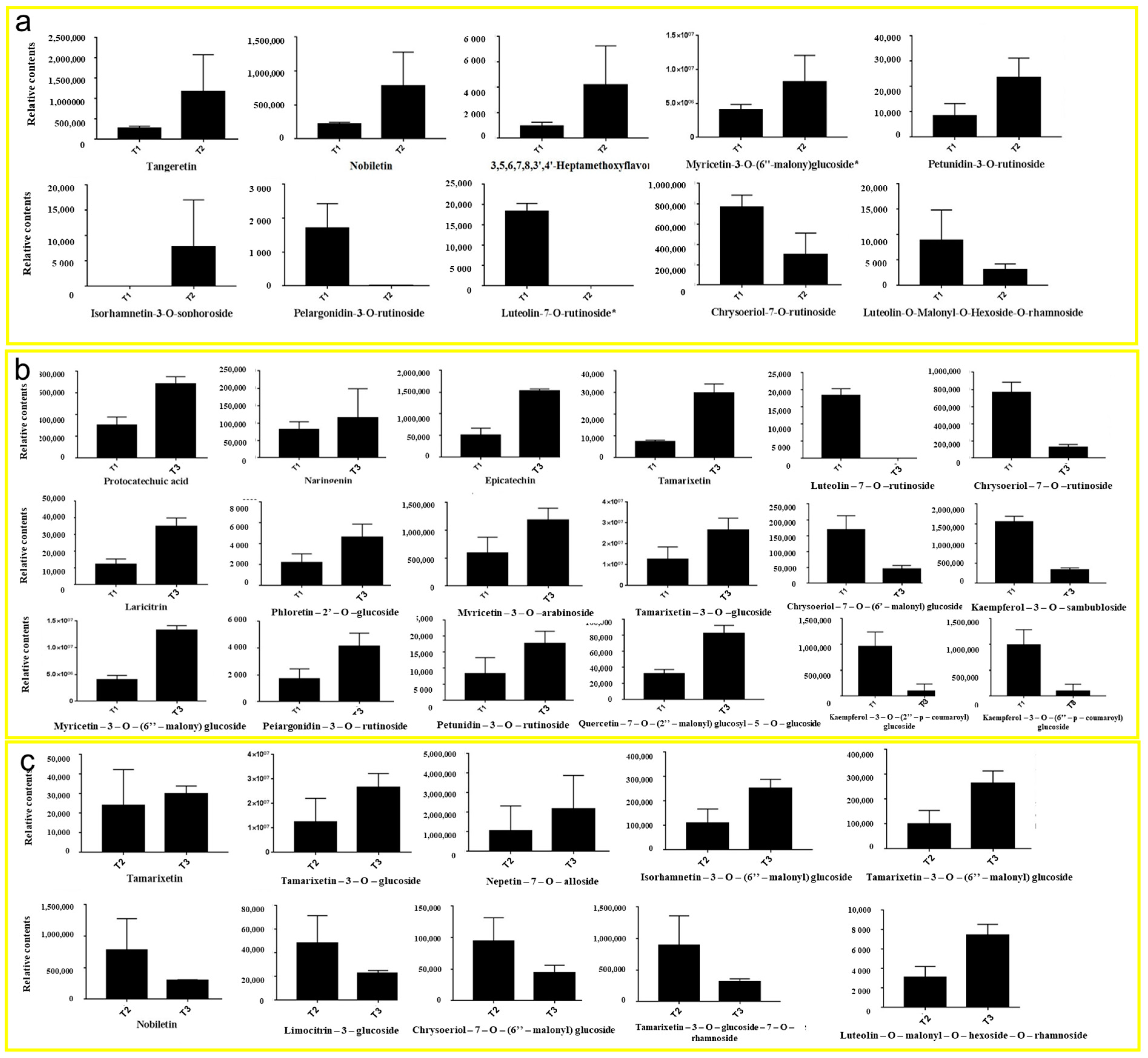
3.4. Metabolome Analysis of Flavonoids in HSK
3.5. Comparison of DAMs in HSK Flowers at Different Developmental Stages
3.6. Transcriptome Analysis in HSK at Different Flower Stages
3.7. KEGG Analysis of Differentially Expressed Genes in HSK
3.8. RT-PCR Validation
3.9. Combined Analysis of Transcriptome and Metabolomic Profiles
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, M.J.; Hung, K.Y.; Li, L. Research Progress on Chemical Constituents and Biological Activity of Hibiscus manihot L. J. Anhui Agric. Sci. 2019, 47, 5–19. [Google Scholar]
- Ge, J.X. Multifunctional Plants—Hibiseu manihot L. Spec. Econ. Anim. Plants 2005, 3, 38. [Google Scholar]
- Tang, J.R. Endangered Rare Plant Hibiseu manihot L. Sci. Technol. Sichuan Agric. 2005, 12, 12. [Google Scholar]
- Wu, M.Q. Rare Plants—Hibiseu manihot L. Contemp. Hortic. 2008, 10, 16–17. [Google Scholar]
- Yang, Z.; Tang, H.; Shao, Q.; Bilia, A.R.; Wang, Y.; Zhao, X. Enrichment and Purification of the Bioactive Flavonoids from Flower of Abelmoschus manihot (L.) Medic Using Macroporous Resins. Molecules 2018, 23, 2649. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.H.; Miao, M.S. Analysis of modern research and comprehensive utilization of Annual Herbaceous. Acta Chin. Med. 2016, 31, 1966–1968. [Google Scholar]
- Song, C.P. Exploration of new development model of whole industry chain of Hibiseu manihot L. Agric. Technol. Equip. 2020, 9, 23–26. [Google Scholar]
- Li, J.; Wang, Y.F.; Rong, J.D.; Zhan, S.H.; Wang, Z.H. Nutritional value and antioxidant activity of Abelmoschus manihot. Chin. J. Trop. Crops 2019, 40, 1354–1358. [Google Scholar]
- Pan, X.; Du, L.; Tao, J.; Jiang, S.; Qian, D.; Duan, J. Dynamic changes of flavonoids in Abelmoschus manihot different organs at different growth periods by UPLC-MS/MS. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1059, 21–26. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, X.; Chen, Y.; Gao, J.; Shi, Q.; Tian, L.; Cao, L. Combined Metabolome and Transcriptome Analyses Reveal the Flavonoids Changes and Biosynthesis Mechanisms in Different Organs of Hibiseu manihot L. Front. Plant Sci. 2022, 13, 817378. [Google Scholar] [CrossRef]
- Mahmoudi, R.; Nosratpour, S. Teucrium polium L. essential oil: Phytochemiacl component and antioxidant properties. Int. Food Res. J. 2013, 20, 1697–1701. [Google Scholar]
- Lu, D.; Jia, R.B. Research progress on of Chinese medicinal material Aurea Helianthus. Chin. J. Drug Eval. 2015, 32, 90–92. [Google Scholar]
- Xu, Z.J.; Huang, S.B.; Dai, T.L.; Zhang, F.; Zhang, X. Optimization of ultrasonic extraction of flavonoids from phyllostachyspubescens leaves by response surface methodology. J. Wuzhou Univ. 2020, 30, 8–15. [Google Scholar]
- Wang, L.; Wu, S.; Wang, M.; Li, X.Z.; Wang, D.; Wu, Z.H.; Gao, H. Optimization of Extraction Parameters of Flavonoids from Ablemoschus manihot Using Response Surface Methodology. Mod. Chin. Med. 2016, 18, 907–912. [Google Scholar]
- Wang, S.N.; Song, Z.J.; Huang, Y.J. Optimization of ultrasonic assisted extraction of total flavonoids from platycladus orientalis leaves by response surface methodology. Food Res. Dev. 2020, 41, 88–93. [Google Scholar]
- Guo, H.M.; Zhu, H.S.; Guo, J.X. Comparative study on extraction and detection methods of total flavonoids of alfalfa at different growth stages. Chin. J. Grassl. 2020, 42, 141–146. [Google Scholar]
- Shen, G.Y.; Tan, W. Study on the determination of flavonoids from plantain and its oxidation resistance. Cereals Oils 2020, 33, 95–97. [Google Scholar]
- Zhu, C.H.; Tang, J.M.; Wei, X. Study on the Total Flavonoids Content Determination of the Lepionurus sylvestris BL. and Its Antioxidant Activity. Food Res. Dev. 2019, 40, 168–175. [Google Scholar]
- Xiong, S.L.; Ma, N.; Liao, T.T. Antioxidant activity of flavone with different solvent extracts from Premna microphylla Turcz leaves. Sci. Technol. Food Ind. 2017, 38, 19–24. [Google Scholar]
- Liao, Y.Y.; Wang, Y.X.; Guo, X.N. Study on Extraction and Antioxidant of Total Flavonoids from Loquat Leaves. J. Northwest Minzu Univ. (Nat. Sci. Ed. ) 2020, 41, 60–64. [Google Scholar]
- Wojtunik-Kulesza, K.A. Approach to Optimization of FRAP Methodology for Studies Based on Selected Monoterpenes. Molecules 2020, 25, 5267. [Google Scholar] [CrossRef] [PubMed]
- Ogata, H.; Goto, S.; Sato, K. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Lu, J.; Xing, J.; Du, M.; Wang, M.; Zhang, L.; Li, Y.; Zhang, C.; Wu, Y. Transcriptome and metabolome analyses revealing the potential mechanism of seed germination in Polygonatum cyrtonema. Sci. Rep. 2021, 11, 12161. [Google Scholar] [CrossRef]
- Wang, C.H.; Wang, Y.X.; Liu, H.J. Validation and application by HPLC for simultaneous determination of vitexin-2″-O-glucoside, vitexin-2″-O-rhamnoside, rutin, vitexin, and hyperoside. J. Pharm. Anal. 2011, 1, 291–296. [Google Scholar] [CrossRef]
- Thada, R.; Chockalingam, S.; Dhandapani, R.K.; Panchamoorthy, R. Extraction and quantitation of coumarin from cinnamon and its effect on enzymatic browning in fresh apple juice: A bioinformatics approach to illuminate its antibrowning activity. J. Agric. Food Chem. 2013, 61, 5385–5390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tu, Z.C.; Wang, H.; Fu, Z.F.; Wen, Q.H.; Fan, D. Metabolic profiling of antioxidants constituents in Artemisia selengensis leaves. Food Chem. 2015, 186, 123–132. [Google Scholar] [CrossRef]
- Tang, Z.; Zhou, C.; Cai, Y.; Tang, Y.; Sun, W.; Yao, H.; Zheng, T.; Chen, H.; Xiao, Y.; Shan, Z.; et al. Purification, characterization and antioxidant activities in vitro of polysaccharides from Amaranthus hybridus L. PeerJ 2020, 8, e9077. [Google Scholar] [CrossRef]
- Ionita, P. The Chemistry of DPPH· Free Radical and Congeners. Int. J. Mol. Sci. 2021, 22, 1545. [Google Scholar] [CrossRef]
- Richards, S.L.; Wilkins, K.A.; Swarbreck, S.M.; Anderson, A.A.; Habib, N.; Smith, A.G.; McAinsh, M.; Davies, J.M. The hydroxyl radical in plants: From seed to seed. J. Exp. Bot. 2015, 66, 37–46. [Google Scholar] [CrossRef]
- Ghosh, S.; Derle, A.; Ahire, M.; More, P.; Jagtap, S.; Phadatare, S.D.; Patil, A.B.; Jabgunde, A.M.; Sharma, G.K.; Shinde, V.S.; et al. Phytochemical analysis and free radical scavenging activity of medicinal plants Gnidia glauca and Dioscorea bulbifera. PLoS ONE 2013, 8, e82529. [Google Scholar] [CrossRef]
- Arikan, B.; Ozfidan-Konakci, C.; Alp, F.N.; Zengin, G.; Yildiztugay, E. Rosmarinic acid and hesperidin regulate gas exchange, chlorophyll fluorescence, antioxidant system and the fatty acid biosynthesis-related gene expression in Arabidopsis thaliana under heat stress. Phytochemistry 2022, 198, 113157. [Google Scholar] [CrossRef] [PubMed]
- Godoy, J.A.; Rios, J.A.; Picón-Pagès, P.; Herrera-Fernández, V.; Swaby, B.; Crepin, G.; Vicente, R.; Fernández-Fernández, J.M.; Muñoz, F.J. Mitostasis, Calcium and Free Radicals in Health, Aging and Neurodegeneration. Biomolecules 2021, 11, 1012. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process | Differential Metabolites | Differential Genes | ||||
|---|---|---|---|---|---|---|
| Compound ID | Compound | Ko ID | Gene Name | Abbreviation | EC ID | |
| T1 vs. T2 | C12644; C08639 | Pelargonidin-3-O-rutinoside; Cyanidin-3,5-O-diglucoside (Cyanin) | K12930 | anthocyanidin 3-O-glucosyltransferase | BZ1 | 2.4.1.115 |
| K12338 | anthocyanidin 3-O-glucoside 5-O-glucosyltransferase | UGT75C1 | 2.4.1.298 | |||
| T1 vs. T3 | C12644; C12095 | Pelargonidin-3-O-rutinoside; Cyanidin-3-O-(6″-O-p-Coumaroyl)glucoside | K12930 | anthocyanidin 3-O-glucosyltransferase | BZ1 | 2.4.1.115 |
| K12338 | anthocyanidin 3-O-glucoside 5-O-glucosyltransferase | UGT75C1 | 2.4.1.298 | |||
| T2 vs. T3 | C12644; C08620; C08639; C16306 | Pelargonidin-3-O-rutinoside; Cyanidin-3-O-rutinoside (Keracyanin); Cyanidin-3,5-O-diglucoside (Cyanin); Cyanidin-3-O-(2″-O-glucosyl)glucoside | K12930 | anthocyanidin 3-O-glucosyltransferase | BZ1 | 2.4.1.115 |
| Process | Differential Metabolites | Differential Genes | ||||
|---|---|---|---|---|---|---|
| Compound ID | Compound | Ko ID | Gene Name | Abbreviation | EC ID | |
| T1 vs. T3 | C00509; C09727; C01604 | Naringenin (5,7,4′-Trihydroxyflavanone); Epicatechin; Phloretin-2′-O-glucoside (Phlorizin) | K13065 | shikimate O-hydroxycinnamoyltransferase | HCT | 2.3.1.133 |
| K13082 | Bifunctional dihydroflavonol 4-reductase | DFR | 1.1.1.219 | |||
| K05278 | flavonol synthase | FLS | 1.14.14.82 | |||
| K00588 | caffeoyl-CoA O-methyltransferase | CCoAOMT | 2.1.1.104 | |||
| K13081 | leucoanthocyanidin reductase | LAR | 1.17.1.3 | |||
| K05277 | anthocyanidin synthase | ANS | 1.14.20.4 | |||
| K09754 | 5-O-(4-coumaroyl)-D-quinate 3′-monooxygenase | C3′H | 1.14.14.96 | |||
| K08695 | anthocyanidin reductase | ANR | 1.3.1.77 | |||
| K01859 | chalcone isomerase | CHI | 5.5.1.6 | |||
| K22845 | phlorizin synthase | PGT1 | 2.4.1.357 | |||
| K00660 | chalcone synthase | CHS | 2.3.1.74 | |||
| T2 vs. T3 | C00509; C01378; C09727 | Naringenin (5,7,4′-Trihydroxyflavanone); Fustin; Epicatechin | K13082 | Bifunctional dihydroflavonol 4-reductase | DFR | 1.1.1.219 |
| K05278 | flavonol synthase | FLS | 1.14.14.82 | |||
| K13081 | leucoanthocyanidin reductase | LAR | 1.17.1.3 | |||
| K13083 | flavonoid 3′,5′-hydroxylase | F3′5′H | 1.14.14.81 | |||
| K05277 | anthocyanidin synthase | ANS | 1.14.20.4 | |||
| K08695 | anthocyanidin reductase | ANR | 1.3.1.77 | |||
| K01859 | chalcone isomerase | CHI | 5.5.1.6 | |||
| K00660 | chalcone synthase | CHS | 2.3.1.74 | |||
| K13065 | shikimate O-hydroxycinnamoyltransferase | HCT | 2.3.1.133 | |||
| K22845 | phlorizin synthase | PGT1 | 2.4.1.357 | |||
| Process | Differential Metabolites | Differential Genes | ||||
|---|---|---|---|---|---|---|
| Compound ID | Compound | Ko ID | Gene Name | Abbreviation | EC ID | |
| T1 vs. T3 | C00509 | Naringenin (5,7,4′-Trihydroxyflavanone); | K13260 | 4′-methoxyisoflavone 2′-hydroxylase | CYP81E | 1.14.14.89 |
| K13264 | isoflavone 7-O-glucoside-6″-O-malonyltransferase | IF7MAT | 2.3.1.115 | |||
| K13265 | vestitone reductase | VR | 1.1.1.348 | |||
| K13258 | 2-hydroxyisoflavanone dehydratase | HIDH | 4.2.1.105 | |||
| T2 vs. T3 | C00509 | Naringenin (5,7,4′-Trihydroxyflavanone); | K13260 | 4′-methoxyisoflavone 2′-hydroxylase | CYP81E | 1.14.14.89 |
| K13264 | isoflavone 7-O-glucoside-6″-O-malonyltransferase | IF7MAT | 2.3.1.115 | |||
| K13265 | vestitone reductase | VR | 1.1.1.348 | |||
| K13258 | 2-hydroxyisoflavanone dehydratase | HIDH | 4.2.1.105 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, J.; Zhou, Y.; Ran, L.; Chen, Y.; Zhang, T.; Sun, B.; Yang, Y.; Sang, Q.; Cao, L. Transcriptome and Metabolome Analysis Reveal the Flavonoid Biosynthesis Mechanism of Abelmoschus manihot L. at Different Anthesis Stages. Metabolites 2023, 13, 216. https://doi.org/10.3390/metabo13020216
Hou J, Zhou Y, Ran L, Chen Y, Zhang T, Sun B, Yang Y, Sang Q, Cao L. Transcriptome and Metabolome Analysis Reveal the Flavonoid Biosynthesis Mechanism of Abelmoschus manihot L. at Different Anthesis Stages. Metabolites. 2023; 13(2):216. https://doi.org/10.3390/metabo13020216
Chicago/Turabian StyleHou, Jiaqi, Yuhan Zhou, Liping Ran, Yanzhu Chen, Ting Zhang, Bowei Sun, Yimo Yang, Qianzi Sang, and Li Cao. 2023. "Transcriptome and Metabolome Analysis Reveal the Flavonoid Biosynthesis Mechanism of Abelmoschus manihot L. at Different Anthesis Stages" Metabolites 13, no. 2: 216. https://doi.org/10.3390/metabo13020216