
Investigation of Phospholipid Differences in Valproic Acid-Induced Autistic Mouse Model Brain Using Mass Spectrometry Imaging
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
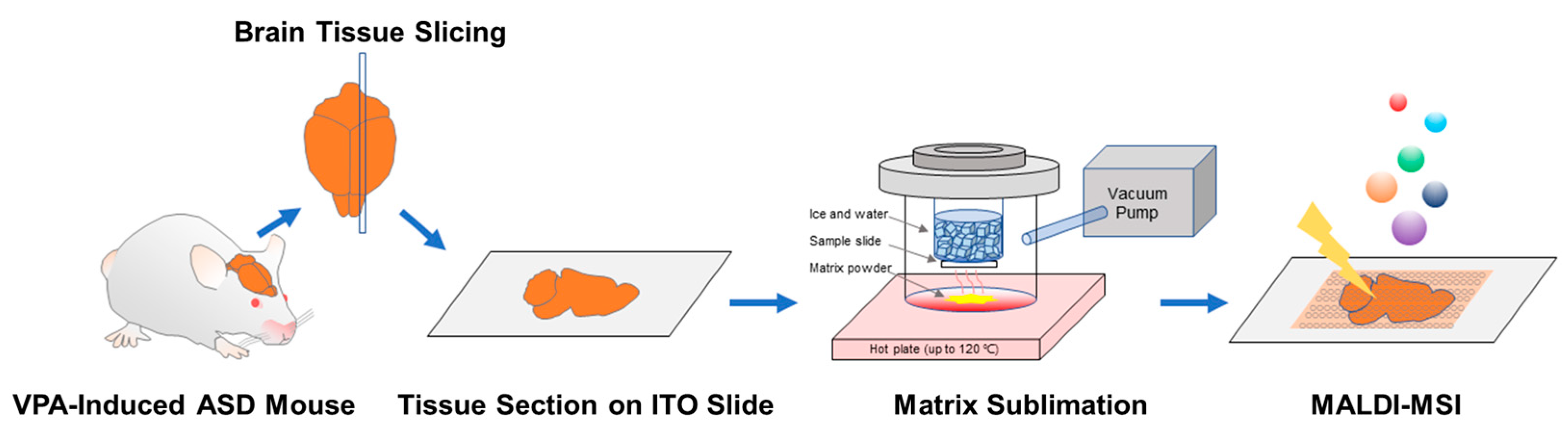
2.1. VPA-Induced ASD Mouse Model and Sample Preparation for MALDI-MSI
2.2. MALDI-MSI
2.3. Tandem Mass Spectrometry
2.4. Data Analysis
3. Results and Discussion
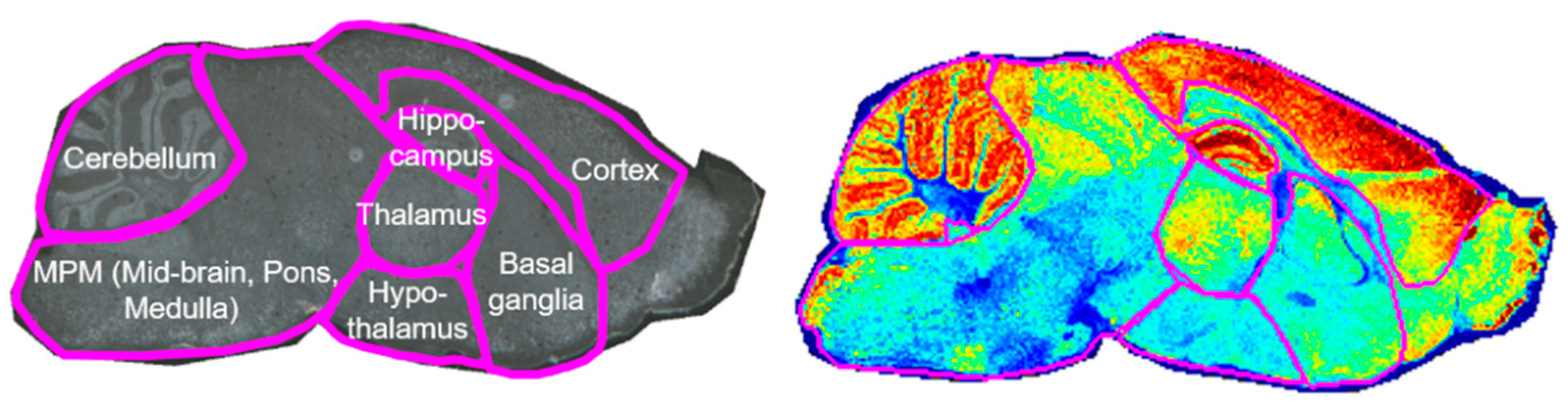
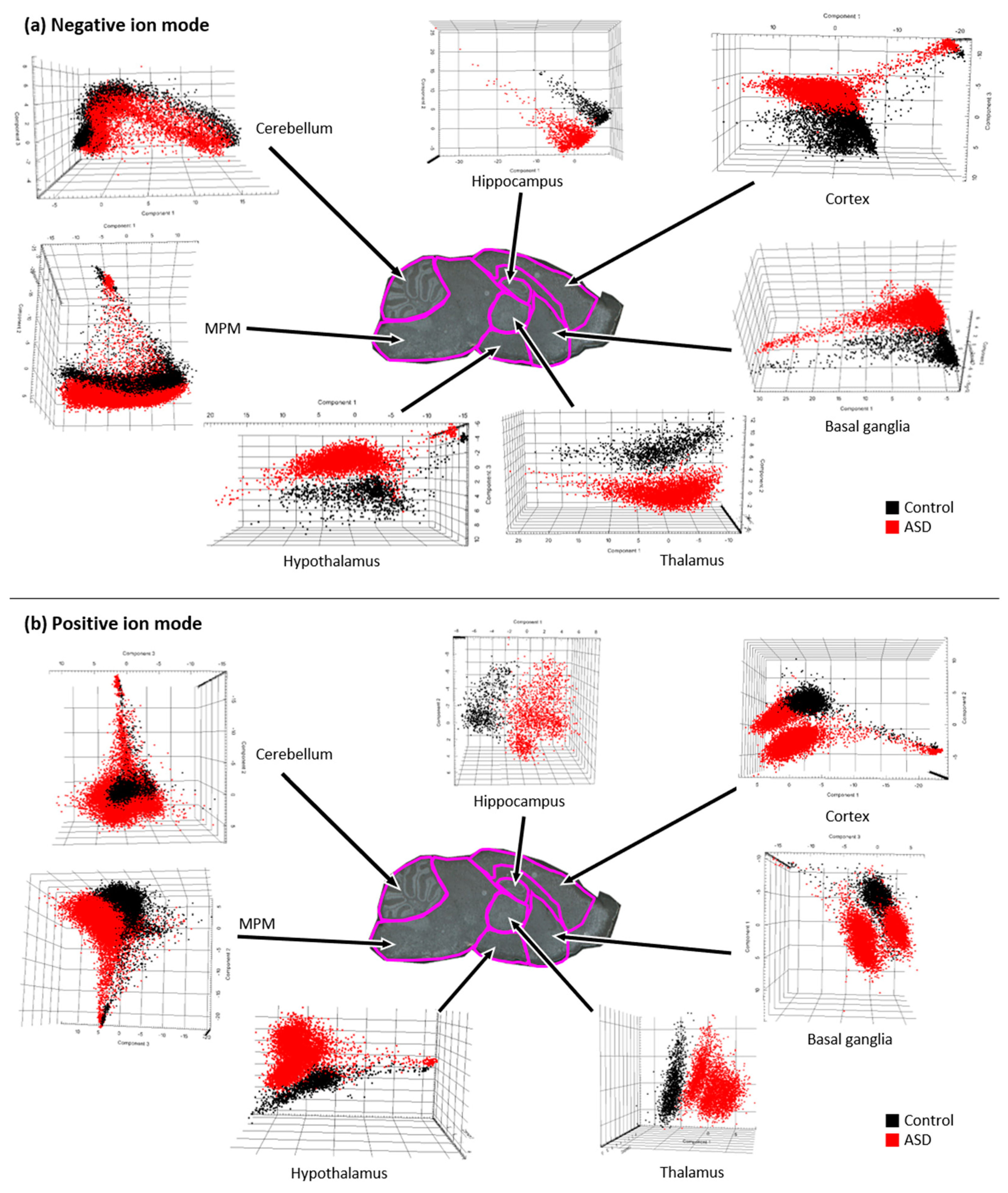
3.1. Brain Compartmentalization of the ASD Mouse Model
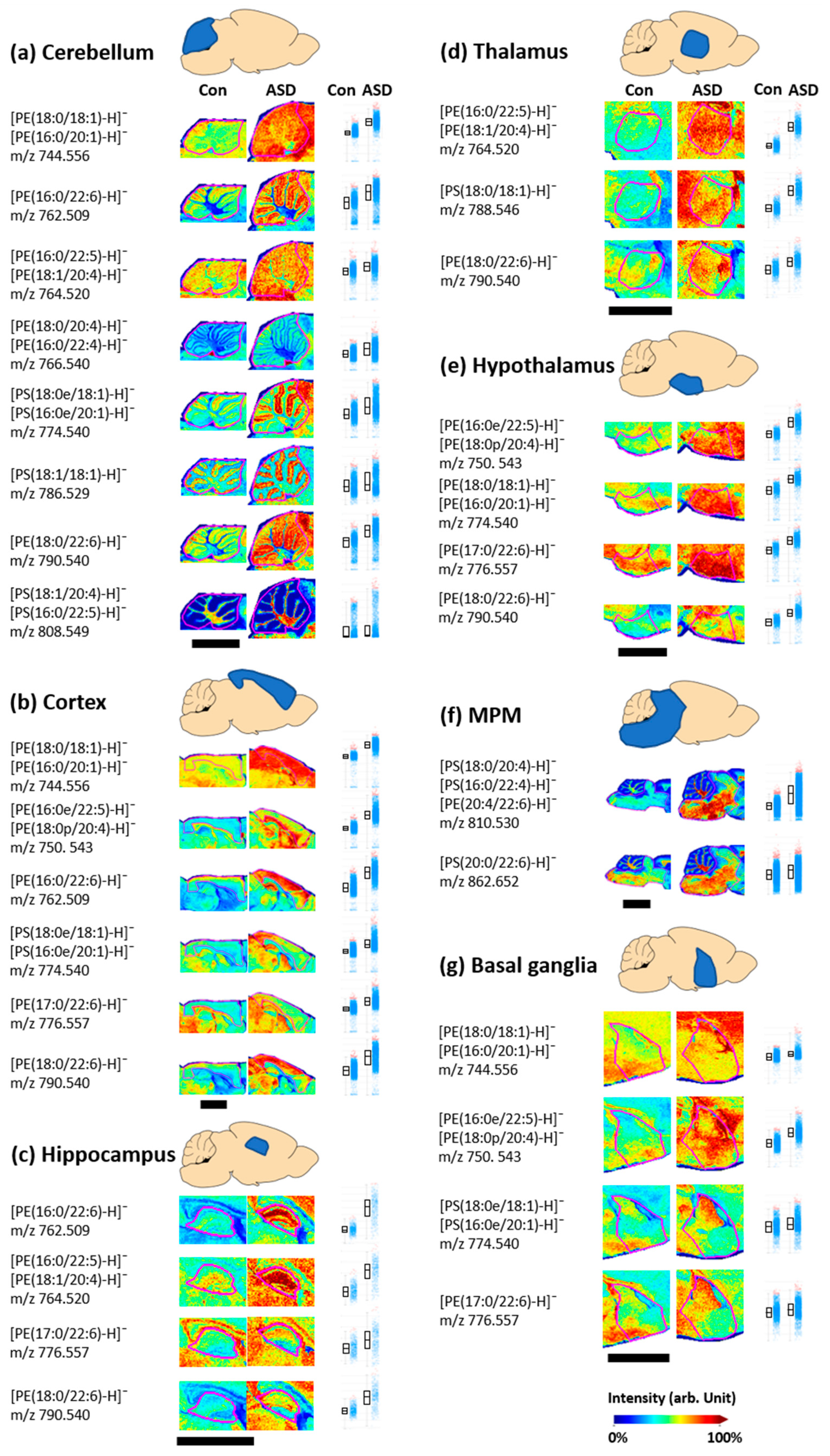
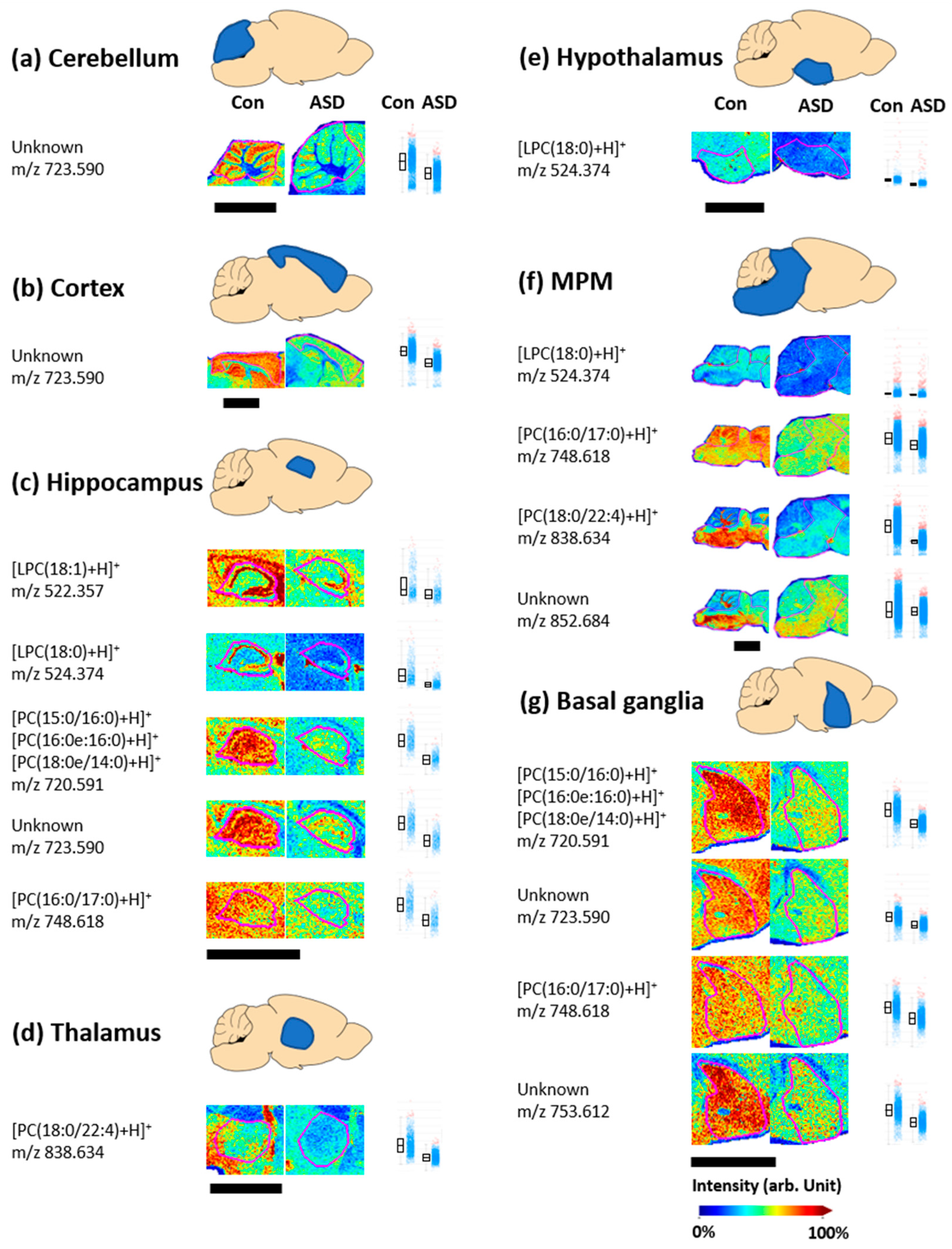
3.2. Changes in Phospholipid Levels in the VPA-Induced ASD Mouse Brain
3.3. Multivariate Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bodzon-Kulakowska, A.; Suder, P. Imaging mass spectrometry: Instrumentation, applications, and combination with other visualization techniques. Mass Spectrom. Rev. 2016, 35, 147–169. [Google Scholar] [CrossRef] [PubMed]
- Buchberger, A.R.; DeLaney, K.; Johnson, J.; Li, L. Mass spectrometry imaging: A review of emerging advancements and future insights. Anal. Chem. 2018, 90, 240. [Google Scholar] [CrossRef] [PubMed]
- Erlmeier, F.; Sun, N.; Shen, J.; Feuchtinger, A.; Buck, A.; Prade, V.M.; Kunzke, T.; Schraml, P.; Moch, H.; Autenrieth, M. MALDI Mass spectrometry imaging—Prognostic pathways and metabolites for renal cell carcinomas. Cancers 2022, 14, 1763. [Google Scholar] [CrossRef] [PubMed]
- Hawkinson, T.R.; Clarke, H.A.; Young, L.E.; Conroy, L.R.; Markussen, K.H.; Kerch, K.M.; Johnson, L.A.; Nelson, P.T.; Wang, C.; Allison, D.B. In situ spatial glycomic imaging of mouse and human Alzheimer’s disease brains. Alzheimer’s Dement. 2022, 18, 1721–1735. [Google Scholar] [CrossRef] [PubMed]
- Shanta, S.R.; Choi, C.S.; Lee, J.H.; Shin, C.Y.; Kim, Y.J.; Kim, K.-H.; Kim, K.P. Global changes in phospholipids identified by MALDI MS in rats with focal cerebral ischemia. J. Lipid Res. 2012, 53, 1823–1831. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Baluya, D.; Tosun, M.; Li, F.; Maletic-Savatic, M. Imaging mass spectrometry: A new tool to assess molecular underpinnings of neurodegeneration. Metabolites 2019, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Schnackenberg, L.K.; Thorn, D.A.; Barnette, D.; Jones, E.E. MALDI imaging mass spectrometry: An emerging tool in neurology. Metab. Brain Dis. 2022, 37, 105–121. [Google Scholar] [CrossRef]
- Shen, L.; Liu, X.; Zhang, H.; Lin, J.; Feng, C.; Iqbal, J. Biomarkers in autism spectrum disorders: Current progress. Clin. Chim. Acta 2020, 502, 41–54. [Google Scholar] [CrossRef]
- Won, H.; Mah, W.; Kim, E. Autism spectrum disorder causes, mechanisms, and treatments: Focus on neuronal synapses. Front. Mol. Neurosci. 2013, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Sisorders: DSM-5; American Psychiatric Association: Washington, DC, USA, 2013; Volume 10. [Google Scholar]
- Goldani, A.A.; Downs, S.R.; Widjaja, F.; Lawton, B.; Hendren, R.L. Biomarkers in autism. Front. Psychiatry 2014, 5, 100. [Google Scholar] [CrossRef]
- Pardo, C.A.; Vargas, D.L.; Zimmerman, A.W. Immunity, neuroglia and neuroinflammation in autism. Int. Rev. Psychiatry 2005, 17, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C.M.; Simonyi, A.; Sun, A.Y.; Sun, G.Y. Phospholipases A2 and neural membrane dynamics: Implications for Alzheimer’s disease. J. Neurochem. 2011, 116, 813–819. [Google Scholar] [CrossRef]
- Brigandi, S.A.; Shao, H.; Qian, S.Y.; Shen, Y.; Wu, B.-L.; Kang, J.X. Autistic children exhibit decreased levels of essential fatty acids in red blood cells. Int. J. Mol. Sci. 2015, 16, 10061–10076. [Google Scholar] [CrossRef] [Green Version]
- Jory, J. Abnormal fatty acids in Canadian children with autism. Nutrition 2016, 32, 474–477. [Google Scholar] [CrossRef]
- Qasem, H.; Al-Ayadhi, L.; Al Dera, H.; El-Ansary, A. Increase of cytosolic phospholipase A2 as hydrolytic enzyme of phospholipids and autism cognitive, social and sensory dysfunction severity. Lipids Health Dis. 2017, 16, 117. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.C.; Kim, P.; Go, H.S.; Choi, C.S.; Yang, S.-I.; Cheong, J.H.; Shin, C.Y.; Ko, K.H. The critical period of valproate exposure to induce autistic symptoms in Sprague–Dawley rats. Toxicol. Lett. 2011, 201, 137–142. [Google Scholar] [CrossRef]
- Schneider, T.; Przewłocki, R. Behavioral alterations in rats prenatally exposed to valproic acid: Animal model of autism. Neuropsychopharmacol. 2005, 30, 80–89. [Google Scholar] [CrossRef]
- Choi, C.S.; Gonzales, E.L.; Kim, K.C.; Yang, S.M.; Kim, J.-W.; Mabunga, D.F.; Cheong, J.H.; Han, S.-H.; Bahn, G.H.; Shin, C.Y. The transgenerational inheritance of autism-like phenotypes in mice exposed to valproic acid during pregnancy. Sci. Rep. 2016, 6, 36250. [Google Scholar] [CrossRef]
- Mabunga, D.F.N.; Gonzales, E.L.T.; Kim, J.-W.; Kim, K.C.; Shin, C.Y. Exploring the validity of valproic acid animal model of autism. Exp. Neurobiol. 2015, 24, 285. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.J.; Le, M.U.T.; Park, J.H.; Chung, C.G.; Shon, J.G.; Lee, G.S.; Moon, J.H.; Lee, S.B.; Choi, J.S.; Lee, T.G. Matrix-assisted laser desorption/ionization mass spectrometry imaging of phospholipid changes in a Drosophila model of early amyotrophic lateral sclerosis. J. Am. Soc. Mass Spectrom. 2021, 32, 2536–2545. [Google Scholar] [CrossRef]
- Jang, H.J.; Park, J.H.; Lee, G.S.; Lee, S.B.; Moon, J.H.; Choi, J.S.; Lee, T.G.; Yoon, S. Comparison of lipid profiles in head and brain samples of Drosophila melanogaster using electrospray ionization mass spectrometry (ESI-MS). Mass Spectrom. Lett. 2019, 10, 11–17. [Google Scholar]
- Wang, S.S.-H.; Kloth, A.D.; Badura, A. The cerebellum, sensitive periods, and autism. Neuron 2014, 83, 518–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gąssowska-Dobrowolska, M.; Kolasa, A.; Beversdorf, D.Q.; Adamczyk, A. Alterations in cerebellar microtubule cytoskeletal network in a valproic acid-induced rat model of autism spectrum disorders. Biomedicines 2022, 10, 3031. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, K.; Yeung, M.K.; Chan, A.S.; Han, Y.M.Y. Effortful control and prefrontal cortex functioning in children with autism spectrum disorder: An fNIRS study. Brain Sci. 2020, 10, 880. [Google Scholar] [CrossRef]
- Sacai, H.; Sakoori, K.; Konno, K.; Nagahama, K.; Suzuki, H.; Watanabe, T.; Watanabe, M.; Uesaka, N.; Kano, M. Autism spectrum disorder-like behavior caused by reduced excitatory synaptic transmission in pyramidal neurons of mouse prefrontal cortex. Nat. Commun. 2020, 11, 5140. [Google Scholar] [CrossRef]
- Richards, R.; Greimel, E.; Kliemann, D.; Koerte, I.K.; Schulte-Körne, G.; Reuter, M.; Wachinger, C. Increased hippocampal shape asymmetry and volumetric ventricular asymmetry in autism spectrum disorder. NeuroImage Clin. 2020, 26, 102207. [Google Scholar] [CrossRef]
- Banker, S.M.; Gu, X.; Schiller, D.; Foss-Feig, J.H. Hippocampal contributions to social and cognitive deficits in autism spectrum disorder. Trends Neurosci. 2021, 44, 793–807. [Google Scholar] [CrossRef]
- Ayub, R.; Sun, K.L.; Flores, R.E.; Lam, V.T.; Jo, B.; Saggar, M.; Fung, L.K. Thalamocortical connectivity is associated with autism symptoms in high-functioning adults with autism and typically developing adults. Transl. Psychiatry 2021, 11, 93. [Google Scholar] [CrossRef]
- Schuetze, M.; Park, M.T.M.; Cho, I.Y.; MacMaster, F.P.; Chakravarty, M.M.; Bray, S.L. Morphological alterations in the thalamus, striatum, and pallidum in autism spectrum disorder. Neuropsychopharmacology 2016, 41, 2627–2637. [Google Scholar] [CrossRef] [Green Version]
- Caria, A.; Ciringione, L.; de Falco, S. Morphofunctional alterations of the hypothalamus and social behavior in autism spectrum disorders. Brain Sci. 2020, 10, 435. [Google Scholar] [CrossRef]
- Dadalko, O.I.; Travers, B.G. Evidence for brainstem contributions to autism spectrum disorders. Front. Integr. Neurosci. 2018, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Abbott, A.E.; Linke, A.C.; Nair, A.; Jahedi, A.; Alba, L.A.; Keown, C.L.; Fishman, I.; Müller, R.-A. Repetitive behaviors in autism are linked to imbalance of corticostriatal connectivity: A functional connectivity MRI study. Soc. Cogn. Affect. Neurosci. 2018, 13, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Kohls, G.; Yerys, B.; Schultz, R.T. Striatal development in autism: Repetitive behaviors and the reward circuitry. Biol. Psychiatry 2014, 76, 358. [Google Scholar] [CrossRef] [Green Version]
- Qiu, T.; Chang, C.; Li, Y.; Qian, L.; Xiao, C.Y.; Xiao, T.; Xiao, X.; Xiao, Y.H.; Chu, K.K.; Lewis, M.H. Two years changes in the development of caudate nucleus are involved in restricted repetitive behaviors in 2–5-year-old children with autism spectrum disorder. Dev. Cogn. Neurosci. 2016, 19, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Pastural, É.; Ritchie, S.; Lu, Y.; Jin, W.; Kavianpour, A.; Su-Myat, K.K.; Heath, D.; Wood, P.L.; Fisk, M.; Goodenowe, D.B. Novel plasma phospholipid biomarkers of autism: Mitochondrial dysfunction as a putative causative mechanism. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 253–264. [Google Scholar] [CrossRef]
- Paoletti, L.; Elena, C.; Domizi, P.; Banchio, C. Role of phosphatidylcholine during neuronal differentiation. IUBMB Life 2011, 63, 714–720. [Google Scholar] [CrossRef]
- Law, S.-H.; Chan, M.-L.; Marathe, G.K.; Parveen, F.; Chen, C.-H.; Ke, L.-Y. An updated review of lysophosphatidylcholine metabolism in human diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [Green Version]
- Needham, B.D.; Adame, M.D.; Serena, G.; Rose, D.R.; Preston, G.M.; Conrad, M.C.; Campbell, A.S.; Donabedian, D.H.; Fasano, A.; Ashwood, P. Plasma and fecal metabolite profiles in autism spectrum disorder. Biol. Psychiatry 2021, 89, 451–462. [Google Scholar] [CrossRef]
- El-Ansary, A.K.; Ben Bacha, A.G.; Al-Ayahdi, L.Y. Plasma fatty acids as diagnostic markers in autistic patients from Saudi Arabia. Lipids Health Dis. 2011, 10, 62. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, A.; Chauhan, V.; Brown, W.T.; Cohen, I. Oxidative stress in autism: Increased lipid peroxidation and reduced serum levels of ceruloplasmin and transferrin-the antioxidant proteins. Life Sci. 2004, 75, 2539–2549. [Google Scholar] [CrossRef]
- El-Ansary, A.K.; Ben Bacha, A.G.; Al-Ayadhi, L.Y. Impaired plasma phospholipids and relative amounts of essential polyunsaturated fatty acids in autistic patients from Saudi Arabia. Lipids Health Dis. 2011, 10, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamlin, J.C.; Pauly, M.; Melnyk, S.; Pavliv, O.; Starrett, W.; Crook, T.A.; James, S.J. Dietary intake and plasma levels of choline and betaine in children with autism spectrum disorders. Autism Res. Treat. 2013, 2013, 578429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Yin, T.; Shinozaki, K.; Lampe, J.W.; Stevens, J.F.; Becker, L.B.; Kim, J. Comprehensive analysis of phospholipids in the brain, heart, kidney, and liver: Brain phospholipids are least enriched with polyunsaturated fatty acids. Mol. Cell. Biochem. 2018, 442, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Hussain, G.; Wang, J.; Rasul, A.; Anwar, H.; Imran, A.; Qasim, M.; Zafar, S.; Kamran, S.K.S.; Razzaq, A.; Aziz, N. Role of cholesterol and sphingolipids in brain development and neurological diseases. Lipids Health Dis. 2019, 18, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.M.; Austin, D.W. Autistic disorder and phospholipids: A review. Prostaglandins Leukot. Essent. Fat. Acids 2011, 84, 25–30. [Google Scholar] [CrossRef]
- Salem Jr, N.; Litman, B.; Kim, H.Y.; Gawrisch, K. Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids 2001, 36, 945–959. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A. Fatty acids in dyslexia, dyspraxia, ADHD and the autistic spectrum. Nutr. Pract. 2001, 3, 18–24. [Google Scholar]
- Chauhan, A.; Chauhan, V. Oxidative stress in autism. Pathophysiology 2006, 13, 171–181. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Da Costa, K.-A. Choline: An essential nutrient for public health. Nutr. Rev. 2009, 67, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Minshew, N.J.; Goldstein, G.; Dombrowski, S.M.; Panchalingam, K.; Pettegrew, J.W. A preliminary 31P MRS study of autism: Evidence for undersynthesis and increased degradation of brain membranes. Biol. Psychiatry 1993, 33, 762–773. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| m/z | Phospholipid Ions | Fatty Acids | Fragment Ions | References | |
|---|---|---|---|---|---|
| Positive ions | 522.3571 | [LPC + H]+ | 18:1 | 184, 240, 258, 504 | Lipid DB a |
| 524.3729 | [LPC + H]+ | 18:0 | 86, 104, 109, 125, 163, 184, 258, 341, 447, 506 | Lipid DB a | |
| 720.5916 | [PC + H]+ | 15:0/16:0 16:0e/16:0 18:0e/14:0 | 184, 537 184, 537 184, 482, 496 | Lipid DB a | |
| 723.5899 | Unknown | ||||
| 748.5872 | [PC + H]+ | 16:0/17:0 | 184, 478, 492, 496, 510 | Lipid DB a | |
| 753.6016 | Unknown | ||||
| 838.6337 | [PC + H]+ | 18:0/22:4 | 184, 506, 524 | Lipid DB a | |
| 852.6444 | Unknown | ||||
| Negative ions | 744.5547 | [PE − H]− | 18:0/18:1 16:0/20:1 | 79, 140, 153, 281, 283, 460, 462, 478, 480 79, 140, 153, 255, 309, 452, 488, 506 | Lipid DB b |
| 750.5442 | [PE − H]− | 16:0e/22:5 18:0p/20:4 | 79, 153, 258, 329, 420, 438 79, 153, 259, 303, 446, 464 | Lipid DB b | |
| 762.5082 | [PE − H]− | 16:0/22:6 | 79, 97, 122, 140, 153, 255, 283, 327, 434, 452, 506, 524 | Lipid DB b | |
| 764.5237 | [PE − H]− | 16:0/22:5 18:1/20:4 | 79, 97, 122, 140, 153, 255, 285, 329, 434, 452 79, 97, 122, 140, 153, 259, 281, 303, 460, 478, 482, 500 | Lipid DB b | |
| 766.5393 | [PE − H]− | 18:0/20:4 16:0/22:4 | 79, 97, 122, 140, 153, 259, 283, 303, 462, 480, 500 79, 97, 122, 140, 153, 255, 331, 434, 452 | Lipid DB b | |
| 774.5392 | [PS − H]− | 18:0e/18:1 16:0e/20:1 | 79, 153, 281, 405, 423, 687 79, 153, 309, 377, 687 | Lipid DB b | |
| 776.5565 | [PE − H]− | 17:0/22:6 | 79, 140, 153, 269, 283, 327, 448, 466 | Lipid DB b | |
| 786.5299 | [PS − H]− | 18:1/18:1 | 79, 97, 153, 281, 417, 435, 699 | Lipid DB b | |
| 788.5458 | [PS − H]− | 18:0/18:1 | 79, 97, 153, 281, 283, 435, 437, 701 | Lipid DB b | |
| 790.5398 | [PE − H]− | 18:0/22:6 | 79, 97, 122, 140, 153, 283, 327, 462, 480, 506, 524 | Lipid DB b | |
| 808.5107 | [PS − H]− | 18:1/20:4 16:0/22:5 | 79, 97, 153, 259, 281, 303, 417, 435, 439, 721 79, 97, 153, 255, 329, 391, 721 | Lipid DB b | |
| 810.5300 | [PE − H]− [PS − H]− | 18:0/20:4 16:0/22:4 20:4/22:6 | 79, 97, 153, 259, 283, 303, 419, 437, 439, 457, 723 79, 97, 153, 255, 331, 391, 467, 723 79, 97, 153, 259, 283, 303, 327 | Lipid DB b | |
| 862.6080 | [PS − H]− | 20:0/22:6 | 79, 97, 153, 283, 311, 327, 447, 465, 775 | Lipid DB b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, H.J.; Kwon, K.J.; Shin, C.Y.; Lee, G.S.; Moon, J.H.; Lee, T.G.; Yoon, S. Investigation of Phospholipid Differences in Valproic Acid-Induced Autistic Mouse Model Brain Using Mass Spectrometry Imaging. Metabolites 2023, 13, 178. https://doi.org/10.3390/metabo13020178
Jang HJ, Kwon KJ, Shin CY, Lee GS, Moon JH, Lee TG, Yoon S. Investigation of Phospholipid Differences in Valproic Acid-Induced Autistic Mouse Model Brain Using Mass Spectrometry Imaging. Metabolites. 2023; 13(2):178. https://doi.org/10.3390/metabo13020178
Chicago/Turabian StyleJang, Hyun Jun, Kyoung Ja Kwon, Chan Young Shin, Ga Seul Lee, Jeong Hee Moon, Tae Geol Lee, and Sohee Yoon. 2023. "Investigation of Phospholipid Differences in Valproic Acid-Induced Autistic Mouse Model Brain Using Mass Spectrometry Imaging" Metabolites 13, no. 2: 178. https://doi.org/10.3390/metabo13020178