
Influence of Vitamin D Status and Supplementation on Metabolomic Profiles of Older Adults

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Study Design
2.2. Dietary and Lifestyle Assessment
2.3. Blood Sample Collection and Analysis
2.4. Serum Vitamin D Measurement
2.5. Biomarkers of Metabolic Health
2.6. Metabolomics Analysis
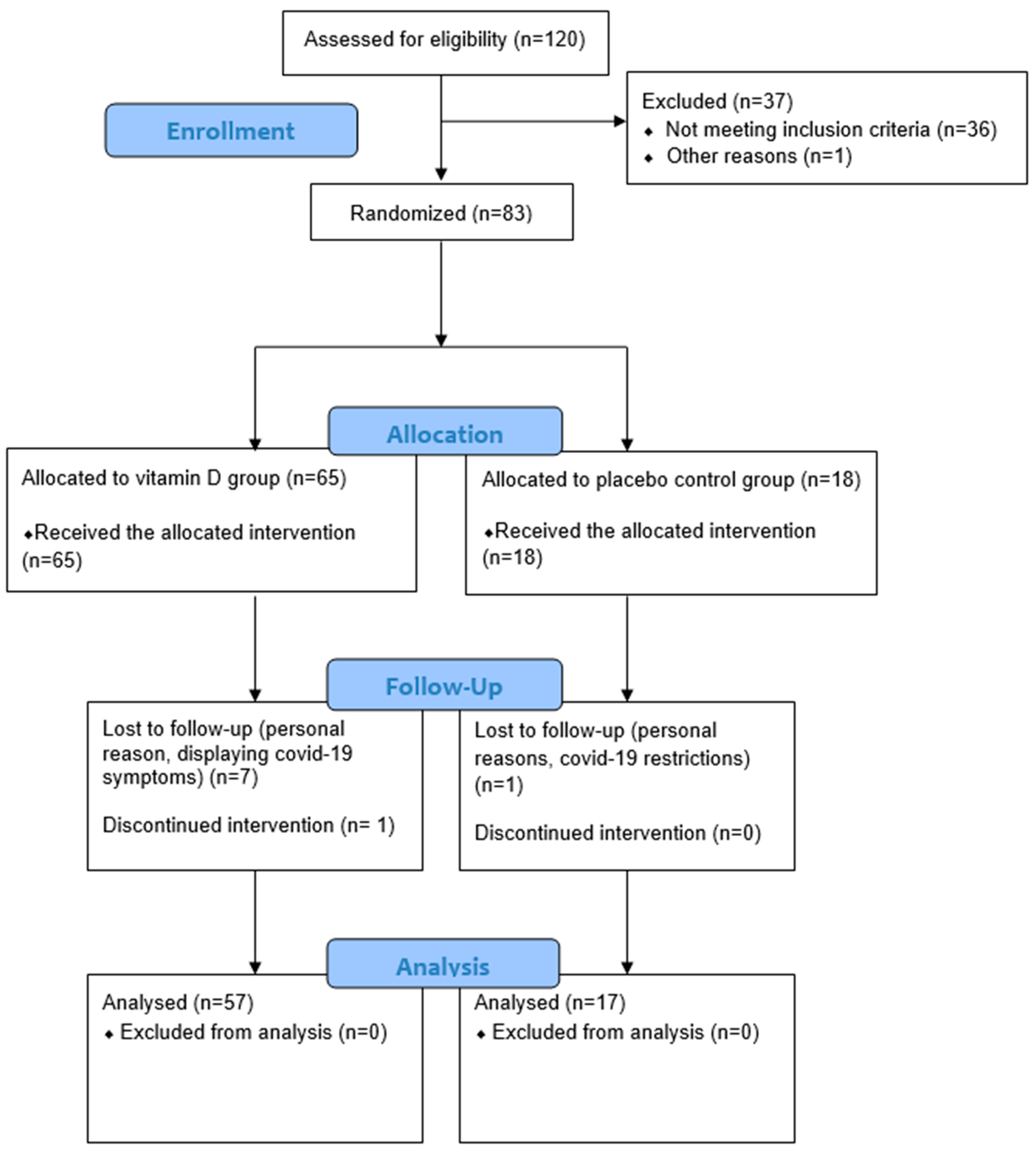
2.7. Participant Flow and Data Collection
2.8. Statistical Analysis
3. Results
3.1. Participant Characteristics
3.2. Baseline Vitamin D and Metabolomic Profiles
3.3. Effect of Vitamin D Supplementation on Metabolomic Profiles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brennan, L. Metabolomics in nutrition research—A powerful window into nutritional metabolism. Essays Biochem. 2016, 60, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Emwas, A.H. The strengths and weaknesses of NMR spectroscopy and mass spectrometry with particular focus on metabolomics research. Methods Mol. Biol. 2015, 1277, 161–193. [Google Scholar] [CrossRef] [PubMed]
- Schrimpe-Rutledge, A.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies-Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, R.Y.H.; Li, G.H.Y.; Cheung, B.M.Y.; Tan, K.C.B.; Kung, A.W.C.; Cheung, C.L. Serum metabolomic profiling and its association with 25-hydroxyvitamin D. Clin. Nutr. 2020, 39, 1179–1187. [Google Scholar] [CrossRef]
- Prendiville, O.; Walton, J.; Flynn, A.; Nugent, A.P.; McNulty, B.A.; Brennan, L. Classifying Individuals Into a Dietary Pattern Based on Metabolomic Data. Mol. Nutr. Food Res. 2021, 65, 2001183. [Google Scholar] [CrossRef]
- McNamara, A.E.; Walton, J.; Flynn, A.; Nugent, A.P.; McNulty, B.A.; Brennan, L. The Potential of Multi-Biomarker Panels in Nutrition Research: Total Fruit Intake as an Example. Front. Nutr. 2021, 7, 577720. [Google Scholar] [CrossRef]
- O’Sullivan, A.; Gibney, M.J.; Connor, A.O.; Mion, B.; Kaluskar, S.; Cashman, K.D.; Flynn, A.; Shanahan, F.; Brennan, L. Biochemical and metabolomic phenotyping in the identification of a vitamin D responsive metabotype for markers of the metabolic syndrome. Mol. Nutr. Food Res. 2011, 55, 679–690. [Google Scholar] [CrossRef]
- Cheng, D.; Zhao, X.; Yang, S.; Cui, H.; Wang, G. Metabolomic Signature between Metabolically Healthy Overweight/Obese and Metabolically Unhealthy Overweight/Obese: A Systematic Review. Diabetes Metab. Syndr. Obesity Targets Ther. 2021, 14, 991–1010. [Google Scholar] [CrossRef]
- Ross, A.C.; Manson, J.E.; Abrams, S.A.; Aloia, J.F.; Brannon, P.M.; Clinton, S.K.; Durazo-Arvizu, R.A.; Gallagher, J.C.; Gallo, R.L.; Jones, G.; et al. The 2011 report on dietary reference intakes for calcium and vitamin D from the Institute of Medicine: What clinicians need to know. J. Clin. Endocrinol. Metab. 2011, 96, 53–58. [Google Scholar] [CrossRef]
- Park, J.E.; Pichiah, P.B.T.; Cha, Y.-S. Vitamin D and Metabolic Diseases: Growing Roles of Vitamin D. J. Obes. Metab. Syndr. 2018, 27, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Joseph, J.J.; Langan, S.; Lunyera, J.; Kluwe, B.; Williams, A.; Chen, H.; Sachs, M.C.; Hairston, K.G.; Bertoni, A.G.; Hsueh, W.A.; et al. The association of serum vitamin D with incident diabetes in an African American Population. Nutr. Diabetes 2022, 12, 43. [Google Scholar] [CrossRef]
- Yang, J.; Ou-Yang, J.; Huang, J. Low serum vitamin D levels increase the mortality of cardiovascular disease in older adults A dose-response meta-analysis of prospective studies. Medicine 2019, 98, e16733. [Google Scholar] [CrossRef]
- Infante, M.; Ricordi, C.; Sanchez, J.; Clare-Salzler, M.J.; Padilla, N.; Fuenmayor, V.; Chavez, C.; Alvarez, A.; Baidal, D.; Alejandro, R.; et al. Influence of Vitamin D on Islet Autoimmunity and Beta-Cell Function in Type 1 Diabetes. Nutrients 2019, 11, 2185. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, J.L.; Pressman, E.K.; Cooper, E.M.; Kent, T.R.; Bar, H.Y.; O’Brien, K.O. Vitamin D Status Affects Serum Metabolomic Profiles in Pregnant Adolescents. Reprod. Sci. 2015, 22, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Nelson, S.M.; A Panagiotou, O.; Anic, G.M.; Mondul, A.M.; Männistö, S.; Weinstein, S.J.; Albanes, D. Metabolomics analysis of serum 25-hydroxy-vitamin D in the Alpha-Tocopherol, Beta-Carotene Cancer Prevention (ATBC) Study. Int. J. Epidemiol. 2016, 45, 1458–1468. [Google Scholar] [CrossRef] [Green Version]
- Lasky-Su, J.; Dahlin, A.; Litonjua, A.A.; Rogers, A.J.; McGeachie, M.J.; Baron, R.M.; Gazourian, L.; Barragan-Bradford, D.; Fredenburgh, L.E.; Choi, A.M.K.; et al. Metabolome alterations in severe critical illness and vitamin D status. Crit. Care 2017, 21, 193. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Kelly, R.S.; Kachroo, P.; Chu, S.H.; Lee-Sarwar, K.; Chawes, B.L.; Bisgaard, H.; Litonjua, A.A.; Weiss, S.T.; Lasky-Su, J. Plasma 25-Hydroxyvitamin D Concentrations are Associated with Polyunsaturated Fatty Acid Metabolites in Young Children: Results from the Vitamin D Antenatal Asthma Reduction Trial. Metabolites 2020, 10, 151. [Google Scholar] [CrossRef]
- Stepien, M.; Nugent, A.P.; Brennan, L. Metabolic Profiling of Human Peripheral Blood Mononuclear Cells: Influence of Vitamin D Status and Gender. Metabolites 2014, 4, 396–406. [Google Scholar] [CrossRef]
- Bhargava, P.; Fitzgerald, K.C.; Calabresi, P.A.; Mowry, E.M. Metabolic alterations in multiple sclerosis and the impact of vitamin D supplementation. JCI Insight 2017, 2, e95302. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, J.A.; Chong, E.Y.; Walker, D.I.; Chandler, J.D.; Michalski, E.S.; Grossmann, R.E.; Uppal, K.; Li, S.; Frediani, J.K.; Tirouvanziam, R.; et al. Plasma metabolomics in adults with cystic fibrosis during a pulmonary exacerbation: A pilot randomized study of high-dose vitamin D(3) administration. Metabolism 2017, 70, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Bagheri, M.; Djazayery, A.; Qi, L.; Yekaninejad, M.S.; Chamari, M.; Naderi, M.; Ebrahimi, Z.; Koletzko, B.; Uhl, O.; Farzadfar, F. Effectiveness of vitamin D therapy in improving metabolomic biomarkers in obesity phenotypes: Two randomized clinical trials. Int. J. Obes. 2018, 42, 1782–1796. [Google Scholar] [CrossRef] [PubMed]
- Shirvani, A.; Kalajian, T.A.; Song, A.; Allen, R.; Charoenngam, N.; Lewanczuk, R.; Holick, M.F. Variable Genomic and Metabolomic Responses to Varying Doses of Vitamin D Supplementation. Anticancer. Res. 2020, 40, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Bislev, L.S.; Sundekilde, U.K.; Kilic, E.; Dalsgaard, T.K.; Rejnmark, L.; Bertram, H.C. Circulating Levels of Muscle-Related Metabolites Increase in Response to a Daily Moderately High Dose of a Vitamin D3 Supplement in Women with Vitamin D Insufficiency-Secondary Analysis of a Randomized Placebo-Controlled Trial. Nutrients 2020, 12, 1310. [Google Scholar] [CrossRef] [PubMed]
- Amrein, K.; Lasky-Su, J.A.; Dobnig, H.; Christopher, K.B. Metabolomic basis for response to high dose vitamin D in critical illness. Clin. Nutr. 2021, 40, 2053–2060. [Google Scholar] [CrossRef] [PubMed]
- Laird, E.; McNulty, H.; Ward, M.; Hoey, L.; McSorley, E.; Wallace, J.M.W.; Carson, E.; Molloy, A.; Healy, M.; Casey, M.C.; et al. Vitamin D Deficiency Is Associated with Inflammation in Older Irish Adults. J. Clin. Endocrinol. Metab. 2014, 99, 1807–1815. [Google Scholar] [CrossRef] [Green Version]
- Laird, E.; O’Halloran, A.M.; Carey, D.; Healy, M.; O’Connor, D.; Moore, P.; Shannon, T.; Molloy, A.M.; Kenny, R.A. The prevalence of Vitamin D deficiency and the determinants of 25(OH)D concentration in older Irish adults: Data from The Irish Longitudinal Study on Ageing (TILDA). J. Gerontol. A. Biol. Med. Sci 2018, 73, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Scully, H.; Laird, E.; Healy, M.; Walsh, J.B.; Crowley, V.; McCarrol, K. Geomapping Vitamin D Status in a Large City and Surrounding Population-Exploring the Impact of Location and Demographics. Nutrients 2020, 12, 2663. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Foroutan, A.; Fitzsimmons, C.; Mandal, R.; Piri-Moghadam, H.; Zheng, J.; Guo, A.; Li, C.; Guan, L.L.; Wishart, D.S. The Bovine Metabolome. Metabolites 2020, 10, 233. [Google Scholar] [CrossRef]
- Foroutan, A.; Guo, A.C.; Vazquez-Fresno, R.; Lipfert, M.; Zhang, L.; Zheng, J.; Badran, H.; Budinski, Z.; Mandal, R.; Ametaj, B.N.; et al. Chemical Composition of Commercial Cow’s Milk. J. Agric. Food Chem. 2019, 67, 4897–4914. [Google Scholar] [CrossRef]
- Asano, L.; Watanabe, M.; Ryoden, Y.; Usuda, K.; Yamaguchi, T.; Khambu, B.; Takashima, M.; Sato, S.-I.; Sakai, J.; Nagasawa, K.; et al. Vitamin D Metabolite, 25-Hydorxyvitamin D, Regulates Lipid Metabolism by Inducing Degradation of SREBP/SCAP. Cell Chem. Biol. 2017, 24, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Tsutsumi, M.; Alvarez, U.; Avioli, L.V.; Hruska, K.A. Effect of 1,25-dihydroxyvitamin D3 on phospholipid composition of rat renal brush border membrane. Am. J. Physiol. 1985, 249, F117–F123. [Google Scholar] [CrossRef]
- Thompson, V.W.; Deluca, H.F. Vitamin D and phospholipid metabolism. J. Biol. Chem. 1964, 239, 984–989. [Google Scholar] [CrossRef]
- Thomas, R.L.; Jiang, L.; Adams, J.S.; Xu, Z.Z.; Shen, J.; Janssen, S.; Ackermann, G.; Vanderschueren, D.; Pauwels, S.; Knight, R.; et al. Vitamin D metabolites and the gut microbiome in older men. Nat. Commun. 2020, 11, 5997. [Google Scholar] [CrossRef]
- Jang, C.; Chen, L.; Rabinowitz, J.D. Metabolomics and Isotope Tracing. Cell 2018, 173, 822–837. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Vitamin D-Insufficient (<50 nmol/L) | Vitamin D-Sufficient (≥50 nmol/L) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Vitamin D | Placebo Control | Vitamin D | Placebo Control | |||||||
| n = 14 | n = 5 | n = 43 | n = 12 | |||||||
| Mean | SE | Mean | SE | p-Value | Mean | SE | Mean | SE | p-Value | |
| Age (years) | 58.0 | 1.7 | 57.6 | 2.9 | 0.88 | 61.9 | 1.3 | 61.3 | 2.3 | 0.81 |
| BMI (kg/m2) | 29.0 | 0.7 | 27.6 | 1.2 | 0.36 | 28.3 | 0.5 | 27.5 | 0.8 | 0.38 |
| BL 25(OH)D (nmol/L) | 33.8 | 2.5 | 38.9 | 4.3 | 0.33 | 71.4 | 0.3 | 70.4 | 4.0 | 0.83 |
| Post 25(OH)D (nmol/L) | 60.1 | 2.9 | 37.3 | 5.0 | 0.001 | 75.0 | 2.0 | 66.8 | 3.5 | 0.05 |
| BL TC (mmol/L) | 4.9 | 0.2 | 4.3 | 0.3 | 0.06 | 5.1 | 0.2 | 5.6 | 0.3 | 0.15 |
| Post TC (mmol/L) | 5.0 | 0.2 | 4.3 | 0.3 | 0.06 | 5.1 | 0.2 | 5.7 | 0.3 | 0.11 |
| BL HDL (mmol/L) | 1.6 | 0.1 | 1.4 | 0.1 | 0.14 | 1.6 | 0.1 | 1.8 | 0.1 | 0.13 |
| Post HDL (mmol/L) | 1.6 | 0.1 | 1.4 | 0.1 | 0.22 | 1.7 | 0.1 | 1.8 | 0.1 | 0.37 |
| BL LDL (mmol/L) | 2.7 | 0.2 | 2.4 | 0.3 | 0.30 | 3.0 | 0.2 | 3.1 | 0.3 | 0.62 |
| Post LDL (mmol/L) | 2.7 | 0.2 | 2.4 | 0.3 | 0.26 | 3.0 | 0.2 | 3.3 | 0.3 | 0.34 |
| BL TRIG (mmol/L) | 1.3 | 0.2 | 1.2 | 0.4 | 0.78 | 1.2 | 0.1 | 1.3 | 0.2 | 0.35 |
| Post TRIG (mmol/L) | 1.3 | 0.2 | 1.3 | 0.3 | 0.88 | 1.1 | 0.1 | 1.3 | 0.2 | 0.36 |
| BL GLUC (mmol/L) | 5.3 | 0.2 | 5.8 | 0.4 | 0.85 | 5.4 | 0.1 | 5.5 | 0.1 | 0.55 |
| Post GLUC (mmol/L) | 5.3 | 0.2 | 5.7 | 0.4 | 0.46 | 5.2 | 0.2 | 6.3 | 0.4 | 0.027 |
| BL CRP (mmol/L) | 2.3 | 0.5 | 1.5 | 0.8 | 0.37 | 2.2 | 0.3 | 1.8 | 0.6 | 0.60 |
| Post CRP (mmol/L) | 2.5 | 0.6 | 1.5 | 1.1 | 0.44 | 2.0 | 0.3 | 1.8 | 0.5 | 0.75 |
| Insufficient | Sufficient | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| (<50 nmol/L) | (≥50 nmol/L) | ||||||||
| n = 19 | n = 55 | GLM ANCOVA | Pearson’s Correlation | ||||||
| Metabolite (µM) | Mean | SE | Mean | SE | p-Value | q-Value | r | p-Value | q-Value |
| Amino acids | |||||||||
| Threonine | 105.6 | 5.2 | 98.4 | 3.0 | 0.22 | 0.61 | −0.260 | 0.039 | 0.43 |
| Leucine | 119.3 | 4.9 | 108.8 | 3.1 | 0.05 | 0.60 | −0.252 | 0.047 | 0.26 |
| Isoleucine | 65.5 | 2.8 | 57.3 | 1.6 | 0.005 | 0.11 | −0.283 | 0.025 | 0.54 |
| Methionine | 23.3 | 0.9 | 21.8 | 0.4 | 0.12 | 0.37 | −0.271 | 0.031 | 0.15 |
| Biogenic Amines | |||||||||
| Acetyl-ornithine | 0.767 | 0.077 | 1.048 | 0.127 | 0.12 | 0.81 | 0.359 | 0.004 | 0.027 |
| Trans-hydroxy-proline | 8.53 | 0.62 | 7.30 | 0.45 | 0.07 | 1.00 | −0.386 | 0.002 | 0.025 |
| Histidines | |||||||||
| Methylhistidine | 8.64 | 1.14 | 12.08 | 1.05 | 0.044 | 0.24 | 0.034 | 0.79 | 0.91 |
| Short chain fatty acids | |||||||||
| Butyric acid | 1.074 | 0.061 | 0.852 | 0.042 | 0.003 | 0.033 | −0.293 | 0.020 | 0.15 |
| Propionic acid | 0.978 | 0.049 | 0.841 | 0.029 | 0.018 | 0.13 | −0.266 | 0.035 | 0.19 |
| Isobutyric acid | 1.321 | 0.070 | 1.062 | 0.040 | 0.002 | 0.044 | −0.370 | 0.003 | 0.06 |
| Hippuric acid | 5.67 | 0.67 | 7.06 | 0.74 | 0.44 | 0.56 | 0.301 | 0.016 | 0.18 |
| Glycerophospholipids | |||||||||
| LYSOC16.0 | 46.1612 | 2.2421 | 53.0679 | 1.5080 | 0.023 | 0.05 | 0.334 | 0.007 | 0.08 |
| LYSOC18.1 | 12.2015 | 0.7658 | 13.7980 | 0.4883 | 0.14 | 0.18 | 0.254 | 0.045 | 0.12 |
| LYSOC18.0 | 13.3603 | 0.6416 | 15.1602 | 0.5183 | 0.08 | 0.12 | 0.300 | 0.017 | 0.08 |
| LYSOC24.0 | 0.0764 | 0.0044 | 0.0850 | 0.0025 | 0.07 | 0.11 | 0.273 | 0.030 | 0.09 |
| LYSOC26.1 | 0.0644 | 0.0041 | 0.0793 | 0.0028 | 0.008 | 0.030 | 0.334 | 0.007 | 0.06 |
| LYSOC26.0 | 0.3970 | 0.0197 | 0.5063 | 0.0166 | 0.001 | 0.017 | 0.334 | 0.007 | 0.05 |
| LYSOC28.1 | 0.2567 | 0.0116 | 0.3179 | 0.0134 | 0.022 | 0.05 | 0.231 | 0.07 | 0.15 |
| LYSOC28.0 | 0.4684 | 0.0285 | 0.5402 | 0.0183 | 0.042 | 0.08 | 0.230 | 0.07 | 0.15 |
| X14.1SMOH | 5.9577 | 0.1863 | 7.0782 | 0.2366 | 0.019 | 0.05 | 0.186 | 0.14 | 0.23 |
| X16.0SM | 99.2598 | 2.9551 | 111.2394 | 2.5620 | 0.010 | 0.034 | 0.234 | 0.06 | 0.16 |
| PC36.0AE | 1.2157 | 0.0385 | 1.5191 | 0.0448 | <0.001 | <0.001 | 0.337 | 0.007 | 0.24 |
| PC36.6AA | 0.7877 | 0.0501 | 1.0911 | 0.0557 | 0.003 | 0.020 | 0.190 | 0.14 | 0.41 |
| PC36.0AA | 6.0304 | 0.2371 | 7.2132 | 0.2011 | 0.002 | 0.017 | 0.279 | 0.027 | 0.07 |
| X22.2SMOH | 12.7172 | 0.4626 | 14.5825 | 0.4303 | 0.024 | 0.05 | 0.172 | 0.18 | 0.28 |
| PC38.6AA | 58.3369 | 4.1387 | 79.2220 | 3.5197 | 0.001 | 0.011 | 0.295 | 0.019 | 0.07 |
| PC38.0AA | 2.6341 | 0.1392 | 3.3977 | 0.1397 | 0.003 | 0.017 | 0.296 | 0.019 | 0.16 |
| PC40.6AE | 3.5294 | 0.1794 | 4.5342 | 0.1984 | 0.006 | 0.026 | 0.274 | 0.030 | 0.09 |
| X24.1SMOH | 2.5160 | 0.1159 | 2.8307 | 0.0783 | 0.039 | 0.08 | 0.198 | 0.12 | 0.21 |
| PC40.6AA | 15.9087 | 1.0491 | 20.7540 | 0.9707 | 0.011 | 0.034 | 0.221 | 0.08 | 0.16 |
| PC40.2AA | 0.3716 | 0.0168 | 0.4461 | 0.0182 | 0.018 | 0.05 | 0.315 | 0.012 | 0.07 |
| PC40.1AA | 0.2617 | 0.0118 | 0.3296 | 0.0143 | 0.003 | 0.015 | 0.335 | 0.007 | 0.12 |
| Acylcarnitines | |||||||||
| C3OH | 0.0245 | 0.0013 | 0.0277 | 0.0007 | 0.028 | 0.16 | 0.134 | 0.29 | 0.84 |
| C6 | 0.0732 | 0.0041 | 0.0887 | 0.0043 | 0.035 | 0.18 | 0.143 | 0.26 | 0.87 |
| C5OH | 0.0323 | 0.0012 | 0.0355 | 0.0008 | 0.024 | 0.16 | 0.206 | 0.11 | 1.00 |
| C5.1DC | 0.0137 | 0.0005 | 0.0153 | 0.0004 | 0.010 | 0.13 | 0.299 | 0.017 | 0.35 |
| C9 | 0.0381 | 0.0047 | 0.0517 | 0.0036 | 0.005 | 0.20 | 0.317 | 0.011 | 0.46 |
| C12 | 0.0918 | 0.0098 | 0.1204 | 0.0079 | 0.017 | 0.14 | 0.206 | 0.11 | 0.87 |
| C14 | 0.0366 | 0.0031 | 0.0474 | 0.0026 | 0.011 | 0.11 | 0.192 | 0.13 | 1.00 |
| C16 | 0.1045 | 0.0064 | 0.1243 | 0.0044 | 0.007 | 0.14 | 0.188 | 0.14 | 0.93 |
| Low Consumers (<5 µg/day) | High Consumers (≥5 µg/day) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| n = 35 | n = 40 | GLM ANCOVA | Pearson’s Correlation | ||||||
| Metabolite (µM) | Mean | SE | Mean | SE | p-Value | q-Value | r | p-Value | q-Value |
| Amino Acids | |||||||||
| Alanine | 340 | 10 | 378 | 11 | 0.038 | 0.84 | 0.166 | 0.15 | 1.00 |
| Betaine | 34.8 | 1.3 | 36.2 | 1.6 | 0.37 | 0.82 | 0.238 | 0.040 | 0.88 |
| Biogenic Amines | |||||||||
| Trans-hydroxy-proline | 7.64 | 0.60 | 7.29 | 0.47 | 0.43 | 0.76 | 0.248 | 0.032 | 0.45 |
| Short chain fatty acids | |||||||||
| Butyric acid | 1.026 | 0.061 | 0.796 | 0.036 | 0.001 | 0.022 | −0.181 | 0.12 | 0.44 |
| Propionic acid | 0.945 | 0.037 | 0.807 | 0.031 | 0.001 | 0.011 | −0.192 | 0.10 | 0.72 |
| Isobutyric acid | 1.218 | 0.051 | 1.037 | 0.049 | 0.002 | 0.015 | −0.221 | 0.06 | 1.00 |
| Organic acids | |||||||||
| Methylmalonic acid | 0.2075 | 0.0245 | 0.1347 | 0.0095 | 0.007 | 0.039 | −0.195 | 0.09 | 1.00 |
| Glycerophospholipids | |||||||||
| LYSOC17.0 | 0.9392 | 0.0399 | 1.0656 | 0.0544 | 0.020 | 0.10 | 0.292 | 0.011 | 0.38 |
| X14.1SMOH | 6.2794 | 0.2000 | 7.2237 | 0.2910 | 0.004 | 0.14 | 0.109 | 0.35 | 0.67 |
| X16.1SMOH | 3.8978 | 0.1376 | 4.3241 | 0.1696 | 0.027 | 0.11 | 0.142 | 0.22 | 0.70 |
| PC36.6AA | 0.9254 | 0.0641 | 1.1019 | 0.0626 | 0.010 | 0.07 | 0.227 | 0.05 | 0.42 |
| X22.2SMOH | 13.4302 | 0.4098 | 14.8973 | 0.5612 | 0.006 | 0.10 | 0.169 | 0.15 | 0.62 |
| PC38.6AA | 67.2424 | 3.7674 | 82.0791 | 4.5278 | 0.016 | 0.09 | 0.207 | 0.07 | 0.51 |
| PC40.6AE | 3.9610 | 0.1998 | 4.6367 | 0.2450 | 0.009 | 0.08 | 0.254 | 0.028 | 0.48 |
| PC40.6AA | 17.4317 | 0.9263 | 21.9442 | 1.2138 | 0.007 | 0.08 | 0.227 | 0.05 | 0.56 |
| Acylcarnitines | |||||||||
| C3 | 0.2505 | 0.0138 | 0.2859 | 0.0117 | 0.015 | 0.30 | 0.061 | 0.61 | 1.00 |
| C4 | 0.1725 | 0.0100 | 0.2119 | 0.0148 | 0.016 | 0.21 | 0.050 | 0.67 | 1.00 |
| C9 | 0.0412 | 0.0035 | 0.0534 | 0.0045 | 0.005 | 0.20 | 0.083 | 0.48 | 1.00 |
| Insufficient (<50 nmol/L) | Sufficient (≥50 nmol/L) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n = 19 | n = 55 | |||||||||||||
| Baseline | Post-Intervention | RM ANCOVA (q-Value) | Baseline | Post-Intervention | RM ANCOVA (q-Value) | |||||||||
| Metabolite (µM) | Mean | SE | Mean | SE | Time | Treatment | T * T | Mean | SE | Mean | SE | Time | Treatment | T * T |
| Amino Acids | ||||||||||||||
| Leucine | 119.3 | 4.9 | 122.3 | 5.9 | 0.33 | 0.83 | 1.00 | 108.8 | 3.1 | 117.5 | 3.5 | 1.00 | 1.00 | 1.00 |
| Betaine | 34 | 2 | 36.5 | 1.9 | 0.26 | 0.68 | 1.00 | 36.1 | 1.2 | 36.1 | 1.3 | 1.00 | 0.82 | 0.10 |
| Taurine | 62.8 | 4.3 | 67.9 | 4.2 | 0.92 | 0.98 | 0.53 | 66.4 | 2.5 | 67.0 | 2.2 | 1.00 | 0.85 | 0.29 |
| Biogenic Amines | ||||||||||||||
| Serotonin | 1.021 | 0.106 | 1.237 | 0.166 | 0.99 | 1.00 | 0.76 | 0.894 | 0.063 | 0.986 | 0.072 | 1.00 | 1.00 | 0.24 |
| Spermidine | 0.238 | 0.008 | 0.242 | 0.007 | 1.00 | 1.00 | 0.14 | 0.241 | 0.004 | 0.234 | 0.004 | 1.00 | 1.00 | 0.77 |
| Organic Acids | ||||||||||||||
| Choline | 9.36 | 0.44 | 9.16 | 0.37 | 0.24 | 1.00 | 1.00 | 9.48 | 0.28 | 9.34 | 0.24 | 1.00 | 1.00 | 0.80 |
| SCFA | ||||||||||||||
| Butyric acid | 1.074 | 0.061 | 0.934 | 0.053 | 0.27 | 1.00 | 1.00 | 0.852 | 0.042 | 0.96 | 0.052 | 1.00 | 1.00 | 0.94 |
| Propionic acid | 0.978 | 0.049 | 0.97 | 0.057 | 0.22 | 1.00 | 0.95 | 0.841 | 0.029 | 1.032 | 0.036 | 1.00 | 0.98 | 0.98 |
| Isobutryic acid | 1.321 | 0.07 | 1.074 | 0.063 | 0.31 | 1.00 | 0.94 | 1.062 | 0.04 | 1.111 | 0.041 | 1.00 | 1.00 | 0.74 |
| Glycerophospholipids | ||||||||||||||
| LYSOC18.1 | 12.2015 | 0.7658 | 13.6343 | 0.9711 | 1.00 | 0.89 | 1.00 | 13.798 | 0.4883 | 14.3698 | 0.5167 | 0.96 | 0.78 | 0.79 |
| LYSOC20.3 | 0.878 | 0.0834 | 1.0139 | 0.1067 | 1.00 | 0.85 | 0.69 | 0.8636 | 0.0568 | 1.0213 | 0.0578 | 0.99 | 1.00 | 0.17 |
| PC32.2AA | 5.8126 | 0.1846 | 5.8335 | 0.3155 | 1.00 | 0.71 | 0.98 | 6.2932 | 0.2394 | 6.0296 | 0.2074 | 1.00 | 0.73 | 0.85 |
| Acylcarnitines | ||||||||||||||
| C0 | 36.2855 | 1.4732 | 35.161 | 1.7376 | 1.00 | 0.66 | 0.97 | 36.2065 | 1.1691 | 35.8572 | 1.0347 | 1.00 | 0.32 | 0.49 |
| C4OH | 0.0421 | 0.0033 | 0.037 | 0.0023 | 0.96 | 0.51 | 0.96 | 0.0462 | 0.0031 | 0.0393 | 0.0018 | 1.00 | 0.29 | 0.53 |
| C9 | 0.0381 | 0.0047 | 0.0356 | 0.004 | 1.00 | 0.65 | 0.96 | 0.0517 | 0.0036 | 0.0503 | 0.0038 | 1.00 | 0.33 | 0.34 |
| C7DC | 0.0651 | 0.0126 | 0.05 | 0.0123 | 1.00 | 0.86 | 0.53 | 0.0708 | 0.0076 | 0.0559 | 0.0074 | 1.00 | 0.31 | 0.79 |
| C10 2 | 0.0615 | 0.0036 | 0.0441 | 0.0026 | 1.00 | 0.81 | 0.50 | 0.0593 | 0.0021 | 0.048 | 0.0019 | 1.00 | 0.73 | 0.78 |
| C12.1 | 0.1112 | 0.0083 | 0.094 | 0.0071 | 1.00 | 0.06 | 0.97 | 0.1331 | 0.007 | 0.1097 | 0.0063 | 1.00 | 0.25 | 1.00 |
| C12 | 0.0918 | 0.0098 | 0.0741 | 0.0063 | 1.00 | 0.06 | 1.00 | 0.1204 | 0.0079 | 0.0978 | 0.008 | 1.00 | 0.27 | 1.00 |
| C14.2 | 0.0518 | 0.0051 | 0.0382 | 0.0032 | 1.00 | 0.06 | 1.00 | 0.0513 | 0.0033 | 0.0454 | 0.0033 | 1.00 | 0.24 | 0.78 |
| C14.1 | 0.146 | 0.0131 | 0.1082 | 0.0092 | 1.00 | 0.05 | 0.97 | 0.1664 | 0.0102 | 0.1335 | 0.009 | 1.00 | 0.98 | 1.00 |
| C14 | 0.0366 | 0.0031 | 0.0315 | 0.0025 | 1.00 | 0.06 | 1.00 | 0.0474 | 0.0026 | 0.0384 | 0.0024 | 1.00 | 0.28 | 0.72 |
| C12DC | 0.0066 | 0.0003 | 0.0062 | 0.0002 | 1.00 | 0.45 | 1.00 | 0.007 | 0.0002 | 0.0062 | 0.0002 | 1.00 | 0.48 | 0.77 |
| C14.1OH | 0.0198 | 0.0016 | 0.0174 | 0.0012 | 1.00 | 0.020 | 1.00 | 0.0214 | 0.0009 | 0.02 | 0.0009 | 1.00 | 0.45 | 0.74 |
| C16.2 | 0.0129 | 0.0009 | 0.0118 | 0.0008 | 1.00 | <0.001 | 1.00 | 0.0135 | 0.0007 | 0.0127 | 0.0006 | 1.00 | 0.25 | 0.83 |
| C16.1 | 0.0453 | 0.0028 | 0.0435 | 0.0026 | 1.00 | 0.040 | 0.96 | 0.0515 | 0.0022 | 0.0465 | 0.002 | 1.00 | 0.25 | 0.82 |
| C16 | 0.1045 | 0.0064 | 0.101 | 0.0068 | 1.00 | 0.11 | 1.00 | 0.1243 | 0.0044 | 0.1108 | 0.0037 | 1.00 | 0.31 | 0.71 |
| C16.2OH | 0.009 | 0.0004 | 0.0099 | 0.0004 | 1.00 | 0.11 | 1.00 | 0.0097 | 0.0003 | 0.0098 | 0.0003 | 1.00 | 0.25 | 0.75 |
| C16.1OH | 0.0141 | 0.0006 | 0.0137 | 0.0008 | 1.00 | 0.13 | 1.00 | 0.0146 | 0.0005 | 0.0144 | 0.0003 | 1.00 | 0.29 | 0.58 |
| C16OH | 0.0082 | 0.0003 | 0.009 | 0.0004 | 1.00 | 0.19 | 0.81 | 0.0086 | 0.0003 | 0.0093 | 0.0003 | 1.00 | 0.29 | 0.76 |
| C18.2 | 0.0592 | 0.0037 | 0.0547 | 0.0037 | 1.00 | 0.17 | 0.78 | 0.0552 | 0.0021 | 0.0521 | 0.0021 | 1.00 | 0.25 | 0.55 |
| C18.1 | 0.1532 | 0.0085 | 0.1424 | 0.0096 | 1.00 | 0.06 | 1.00 | 0.1619 | 0.0062 | 0.143 | 0.0057 | 1.00 | 0.42 | 0.82 |
| C18 | 0.0431 | 0.0029 | 0.0385 | 0.0024 | 1.00 | 0.42 | 0.96 | 0.0504 | 0.0022 | 0.0419 | 0.0014 | 1.00 | 0.99 | 0.70 |
| C18.1OH | 0.0114 | 0.0005 | 0.0128 | 0.0004 | 1.00 | 0.07 | 1.00 | 0.0117 | 0.0004 | 0.0128 | 0.0004 | 1.00 | 0.35 | 0.24 |
| Metabolite | r | p-Value | q-Value |
|---|---|---|---|
| Baseline 25(OH)D and post-intervention metabolite concentrations | |||
| Glycerophospholipids | |||
| LYSOC26.0 | −0.274 | 0.018 | 0.61 |
| Post-intervention 25(OH)D and metabolite concentrations post-intervention | |||
| Amino acids | |||
| Glycine | 0.291 | 0.021 | 0.12 |
| Taurine | 0.294 | 0.019 | 0.14 |
| Glutamic acid | −0.410 | 0.001 | 0.019 |
| Citrulline | 0.337 | 0.007 | 0.08 |
| Biogenic Amines | |||
| Putrescine | 0.325 | 0.009 | 0.13 |
| Methionine sulfoxide | 0.344 | 0.006 | 0.040 |
| Acetyl-ornithine | 0.314 | 0.012 | 0.06 |
| Organic acids | |||
| HPHPA | 0.266 | 0.035 | 0.39 |
| Succinic acid | 0.352 | 0.005 | 0.10 |
| Methylmalonic acid | 0.251 | 0.047 | 0.26 |
| Homovanillic acid | 0.265 | 0.036 | 0.26 |
| Carbohydrates | |||
| Glucose | −0.263 | 0.037 | 1.00 |
| Glycerophospholipids | |||
| PC36.0AA | 0.333 | 0.008 | 0.26 |
| PC38.0AA | 0.303 | 0.016 | 0.18 |
| PC40.2AA | 0.270 | 0.032 | 0.27 |
| PC40.1AA | 0.323 | 0.010 | 0.17 |
| Acylcarnitines | |||
| C3.1 | −0.264 | 0.036 | 1.00 |
| C12 | 0.249 | 0.049 | 0.98 |
| Baseline 25(OH)D and change in metabolite concentrations | |||
| Glycerophospholipids | |||
| LYSOC16.1 | −0.239 | 0.040 | 0.28 |
| LYSOC16.0 | −0.332 | 0.004 | 0.13 |
| LYSOC17.0 | −0.251 | 0.031 | 0.26 |
| LYSOC18.0 | −0.268 | 0.021 | 0.24 |
| LYSOC26.0 | −0.298 | 0.010 | 0.17 |
| Acylcarnitines | |||
| C5.1DC | 0.243 | 0.037 | 0.019 |
| Change in 25(OH)D and change in metabolite concentrations | |||
| Amino Acids | |||
| Tryptophan | 0.230 | 0.048 | 1.00 |
| Glycerophospholipids | |||
| LYSOC16.1 | 0.269 | 0.020 | 0.14 |
| LYSOC16.0 | 0.338 | 0.003 | 0.06 |
| LYSOC17.0 | 0.249 | 0.032 | 0.16 |
| LYSOC18.0 | 0.309 | 0.007 | 0.08 |
| LYSOC20.3 | 0.264 | 0.023 | 0.13 |
| LYSOC26.0 | 0.356 | 0.002 | 0.06 |
| PC40.6AA | 0.271 | 0.020 | 0.17 |
| Acylcarnitines | |||
| C3OH | 0.275 | 0.018 | 0.70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCourt, A.F.; O’Sullivan, A.M. Influence of Vitamin D Status and Supplementation on Metabolomic Profiles of Older Adults. Metabolites 2023, 13, 166. https://doi.org/10.3390/metabo13020166
McCourt AF, O’Sullivan AM. Influence of Vitamin D Status and Supplementation on Metabolomic Profiles of Older Adults. Metabolites. 2023; 13(2):166. https://doi.org/10.3390/metabo13020166
Chicago/Turabian StyleMcCourt, Aislinn F., and Aifric M. O’Sullivan. 2023. "Influence of Vitamin D Status and Supplementation on Metabolomic Profiles of Older Adults" Metabolites 13, no. 2: 166. https://doi.org/10.3390/metabo13020166