
Urinary ATP Levels Are Controlled by Nucleotidases Released from the Urothelium in a Regulated Manner
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. Ethical Approval
2.3. Denuded Bladder Preparation
2.4. Evaluation of s-NTD Activities in the Lumen of Detrusor-Free Bladder Preparations
2.5. Evaluation of Combined NTD Activities in the Lumen of Detrusor-Free Bladder Preparations
2.6. Evaluation of s-NTDs Activities in the Lamina Propria of Detrusor-Free Bladder Preparations
2.7. Effects of Common ENTPDase Inhibitors on Soluble Nucleotidase Activities
2.8. Effects of Activation and Inhibition of Adenylyl Cyclases on the Release of s-NTDs in the Bladder Lumen and in Lamina Propria
2.9. Effects of Adenosine on the Release of s-NTDs in the Bladder Lumen and the Lamina Propria
2.10. Preparation of 1,N6-Etheno-Nucleotides
2.11. HPLC Analysis of 1,N6-Etheno-Nucleotides
2.12. Automated Capillary Electrophoresis and Immunodetection with Wes Simple Western
2.13. Antibodies
2.14. Drugs and Reagents
2.15. Data Analysis
3. Results
3.1. Enzymes That Hydrolyze eATP Are Available on the Luminal Side of the Bladder Urothelium
3.2. Enzymes That Hydrolyze eATP Are Released in the Bladder Lumen during Filling
3.3. Effects of Common ENTPDase Inhibitors on the Activities of s-NTDs in the Bladder Lumen
3.4. Enzymes That Hydrolyze AMP Are Released in the Bladder Lumen during Filling
3.4.1. Degradation of eAMP in ILS from WT and Nt5e−/− Bladders
3.4.2. Effects of Levamisole and L-p-BT on the Degradation of eAMP in ILS from Nt5e−/− Bladders
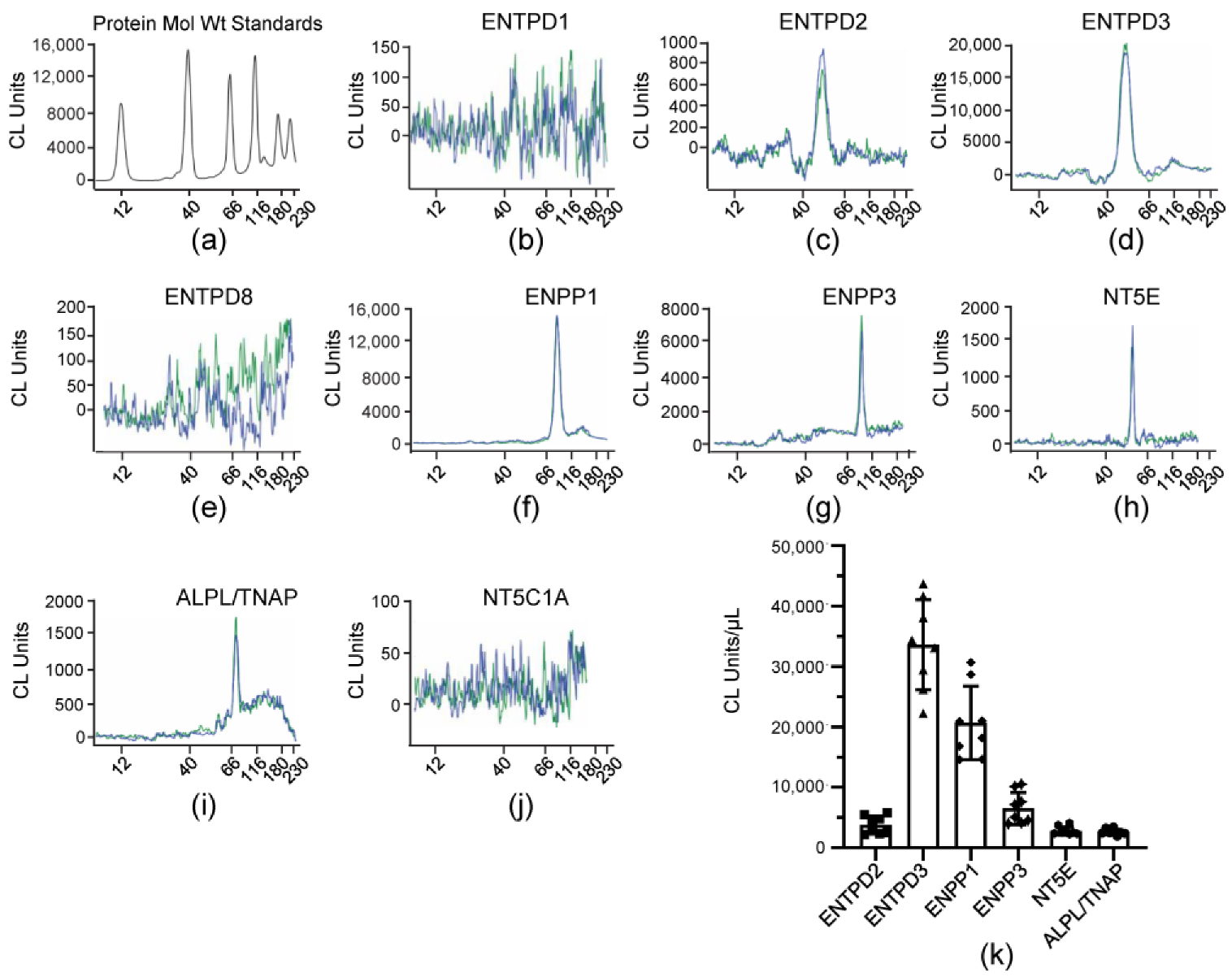
3.5. Soluble Enzymes in the Bladder Lumen Are Similar to Known ENTPDases
3.6. Activation of Adenylyl Cyclases Stimulates the Release of s-NTDs in the Bladder Lumen and Inhibits the Release of s-NTDs in the Lamina Propria
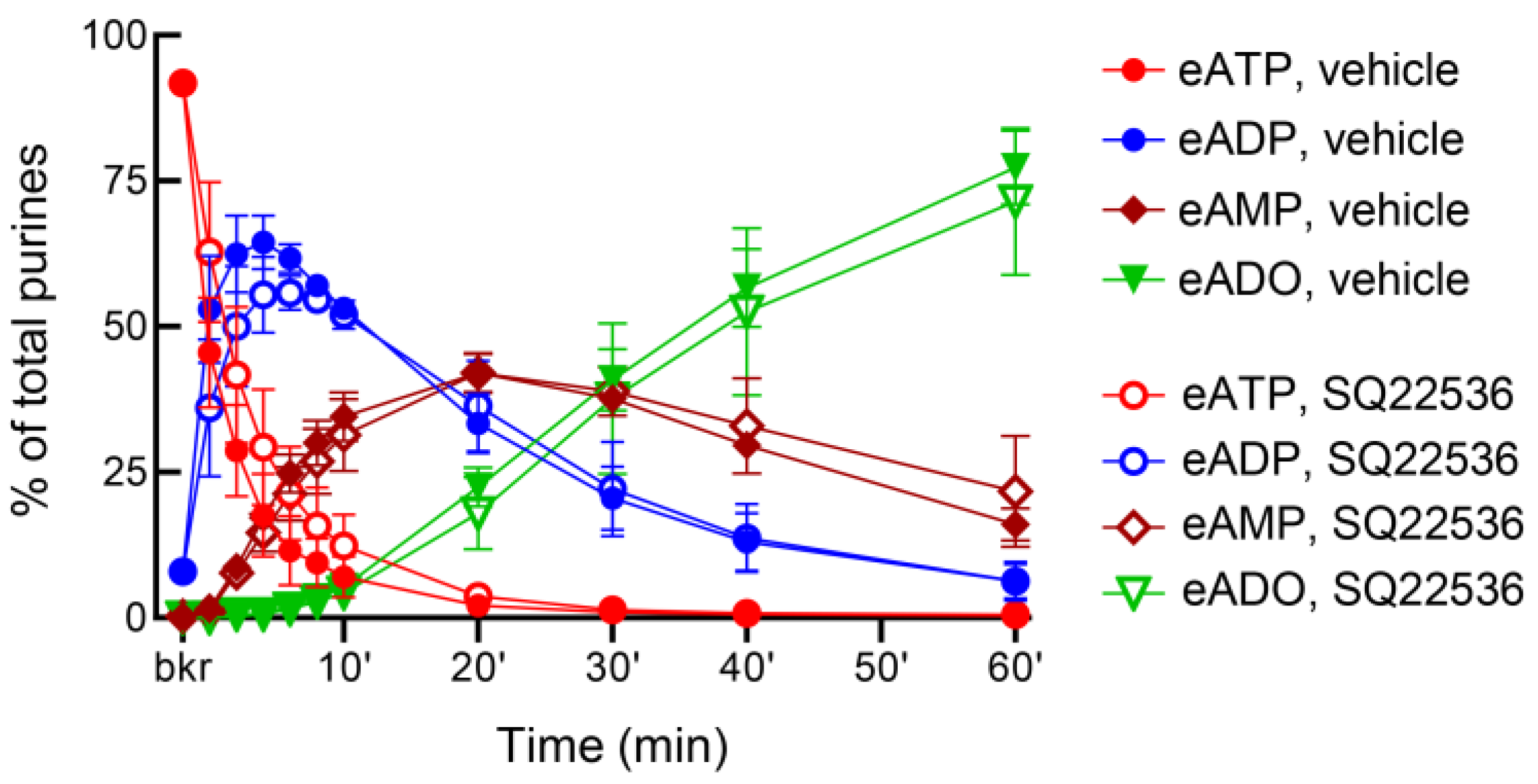
3.7. Inhibition of Adenylyl Cyclases with SQ22536 Has Distinct Effects on the Release of s-NTDs in the Bladder Lumen and in Lamina Propria
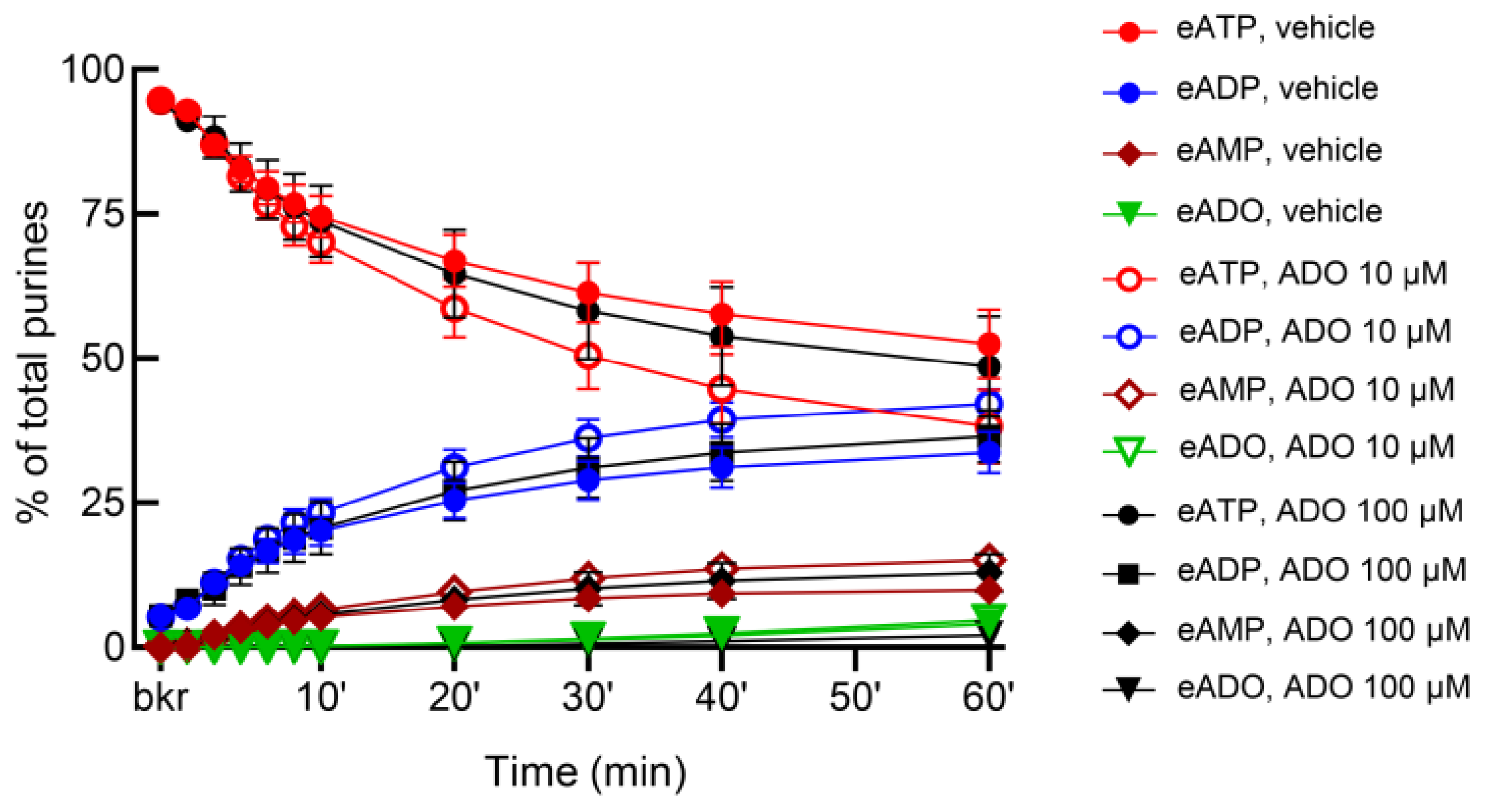
3.8. Adenosine Facilitates the Release of s-NTDs in the Bladder Lumen and Lamina Propria
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daly, D.M.; Nocchi, L.; Liaskos, M.; McKay, N.G.; Chapple, C.; Grundy, D. Age-related changes in afferent pathways and urothelial function in the male mouse bladder. J. Physiol. 2014, 592, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Beckel, J.M.; Daugherty, S.L.; Tyagi, P.; Wolf-Johnston, A.S.; Birder, L.A.; Mitchell, C.H.; de Groat, W.C. Pannexin 1 channels mediate the release of ATP into the lumen of the rat urinary bladder. J. Physiol. 2015, 593, 1857–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durnin, L.; Hayoz, S.; Corrigan, R.D.; Yanez, A.; Koh, S.D.; Mutafova-Yambolieva, V.N. Urothelial purine release during filling of murine and primate bladders. Am. J. Physiol. Renal Physiol. 2016, 311, F708–F716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durnin, L.; Kwok, B.; Kukadia, P.; McAvera, R.; Corrigan, R.D.; Ward, S.M.; Zhang, Y.; Chen, Q.; Koh, S.D.; Sanders, K.M.; et al. An ex vivo bladder model with detrusor smooth muscle removed to analyse biologically active mediators released from the suburothelium. J. Physiol. 2019, 597, 1467–1485. [Google Scholar] [CrossRef]
- Birder, L.; Andersson, K.E. Urothelial signaling. Physiol. Rev. 2013, 93, 653–680. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, E.J.; Merrill, L.; Vizzard, M.A. Bladder sensory physiology: Neuroactive compounds and receptors, sensory transducers, and target-derived growth factors as targets to improve function. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R869–R878. [Google Scholar] [CrossRef] [Green Version]
- Truschel, S.T.; Wang, E.; Ruiz, W.G.; Leung, S.M.; Rojas, R.; Lavelle, J.; Zeidel, M.; Stoffer, D.; Apodaca, G. Stretch-regulated exocytosis/endocytosis in bladder umbrella cells. Mol. Biol. Cell 2002, 13, 830–846. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.C.; Lee, J.-M.; Ruiz, W.G.; Balestreire, E.M.; von Bodungen, M.; Barrick, S.; Cockayne, D.A.; Birder, L.A.; Apodaca, G. ATP and purinergic receptor-dependent membrane traffic in bladder umbrella cells. J. Clin. Investig. 2005, 115, 2412–2422. [Google Scholar] [CrossRef] [Green Version]
- Stenqvist, J.; Winder, M.; Carlsson, T.; Aronsson, P.; Tobin, G. Urothelial acetylcholine involvement in ATP-induced contractile responses of the rat urinary bladder. Eur. J. Pharmacol. 2017, 809, 253–260. [Google Scholar] [CrossRef]
- Sui, G.; Fry, C.H.; Montgomery, B.; Roberts, M.; Wu, R.; Wu, C. Purinergic and muscarinic modulation of ATP release from the urothelium and its paracrine actions. Am. J. Physiol. Renal Physiol. 2014, 306, F286–F298. [Google Scholar] [CrossRef]
- Cheng, Y.; Mansfield, K.J.; Allen, W.; Chess-Williams, R.; Burcher, E.; Moore, K.H. ATP during early bladder stretch is important for urgency in detrusor overactivity patients. Biomed. Res. Int. 2014, 2014, 204604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Ramos, M.; Silva, I.; Oliveira, O.; Ferreira, S.; Reis, M.J.; Oliveira, J.C.; Correia-de-Sa, P. Urinary ATP may be a dynamic biomarker of detrusor overactivity in women with overactive bladder syndrome. PLoS ONE 2013, 8, e64696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Keay, S.; De Deyne, P.G.; Chai, T.C. Augmented stretch activated adenosine triphosphate release from bladder uroepithelial cells in patients with interstitial cystitis. J. Urol. 2001, 166, 1951–1956. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Chai, T.C. Augmented extracellular ATP signaling in bladder urothelial cells from patients with interstitial cystitis. Am. J. Physiol. Cell Physiol. 2006, 290, C27–C34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taidi, Z.; Mansfield, K.J.; Bates, L.; Sana-Ur-Rehman, H.; Liu, L. Purinergic P2X7 receptors as therapeutic targets in interstitial cystitis/bladder pain syndrome; key role of ATP signaling in inflammation. Bladder 2019, 6, e38. [Google Scholar] [CrossRef] [Green Version]
- Abbasian, B.; Shair, A.; O’Gorman, D.B.; Pena-Diaz, A.M.; Brennan, L.; Engelbrecht, K.; Koenig, D.W.; Reid, G.; Burton, J.P. Potential Role of Extracellular ATP Released by Bacteria in Bladder Infection and Contractility. mSphere 2019, 4, e00439-19. [Google Scholar] [CrossRef] [Green Version]
- Ueda, N.; Kondo, M.; Takezawa, K.; Kiuchi, H.; Sekii, Y.; Inagaki, Y.; Soda, T.; Fukuhara, S.; Fujita, K.; Uemura, M.; et al. Bladder urothelium converts bacterial lipopolysaccharide information into neural signaling via an ATP-mediated pathway to enhance the micturition reflex for rapid defense. Sci. Rep. 2020, 10, 21167. [Google Scholar] [CrossRef]
- Xie, X.; Liang, J.; Huang, R.; Luo, C.; Yang, J.; Xing, H.; Zhou, L.; Qiao, H.; Ergu, E.; Chen, H. Molecular pathways underlying tissue injuries in the bladder with ketamine cystitis. FASEB J. 2021, 35, e21703. [Google Scholar] [CrossRef]
- Walsh, C.A.; Cheng, Y.; Mansfield, K.J.; Parkin, K.; Mukerjee, C.; Moore, K.H. Decreased intravesical adenosine triphosphate in patients with refractory detrusor overactivity and bacteriuria. J. Urol. 2013, 189, 1383–1387. [Google Scholar] [CrossRef]
- Krishnan, A.; Sharma, G.; Devana, S.K.; Zohmangaihi, D.; Mavuduru, R.S.; Mandal, A.K.; Sharma, A.P.; Bora, G.S. Urinary adenosine triphosphate and nitric oxide levels in patients with underactive bladder: A preliminary study. World J. Urol. 2021, 39, 4421–4425. [Google Scholar] [CrossRef]
- Munoz, A.; Smith, C.P.; Boone, T.B.; Somogyi, G.T. Overactive and underactive bladder dysfunction is reflected by alterations in urothelial ATP and NO release. Neurochem. Int. 2011, 58, 295–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes-Lopes, T.; Cruz, F. Urinary Biomarkers in Overactive Bladder: Revisiting the Evidence in 2019. Eur. Urol. Focus. 2019, 5, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Firouzmand, S.; Young, J.S. A pilot study to investigate the associations of urinary concentrations of NO, ATP and derivatives with overactive bladder symptom severity. Exp. Physiol. 2020, 105, 932–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiapakidou, S.; Apostolidis, A.; Pantazis, K.; Grimbizis, G.F.; Mikos, T. The use of urinary biomarkers in the diagnosis of overactive bladder in female patients. A systematic review and meta-analysis. Int. Urogynecol. J. 2021, 32, 3143–3155. [Google Scholar] [CrossRef]
- McLatchie, L.; Sahai, A.; Caldwell, A.; Dasgupta, P.; Fry, C. ATP shows more potential as a urinary biomarker than acetylcholine and PGE(2), but its concentration in urine is not a simple function of dilution. Neurourol. Urodyn. 2021, 40, 753–762. [Google Scholar] [CrossRef]
- Lee, W.C.; Wu, H.P.; Tai, T.Y.; Liu, S.P.; Chen, J.; Yu, H.J. Effects of diabetes on female voiding behavior. J. Urol. 2004, 172, 989–992. [Google Scholar] [CrossRef]
- Gomez, C.S.; Kanagarajah, P.; Gousse, A.E. Bladder dysfunction in patients with diabetes. Curr. Urol. Rep. 2011, 12, 419–426. [Google Scholar] [CrossRef]
- Mossa, A.H.; Galan, A.; Cammisotto, P.G.; Velasquez Flores, M.; Shamout, S.; Barcelona, P.; Saragovi, H.U.; Campeau, L. Antagonism of proNGF or its receptor p75(NTR) reverses remodelling and improves bladder function in a mouse model of diabetic voiding dysfunction. Diabetologia 2020, 63, 1932–1946. [Google Scholar] [CrossRef]
- Abler, L.L.; O’Driscoll, C.A.; Colopy, S.A.; Stietz, K.P.K.; Wang, P.; Wang, Z.; Hartmann, F.; Crader-Smith, S.M.; Oellete, J.N.; Mehta, V.; et al. The influence of intermittent hypoxia, obesity, and diabetes on male genitourinary anatomy and voiding physiology. Am. J. Physiol. Renal Physiol. 2021, 321, F82–F92. [Google Scholar] [CrossRef]
- Karaci, M. Obesity contributes to lower urinary system voiding dysfunction in childhood. Ir. J. Med. Sci. 2021, 190, 1459–1463. [Google Scholar] [CrossRef]
- Son, H.; Lee, S.L.; Park, W.H.; Park, K.; Park, S.; Kang, M.S.; Kim, D.Y.; Kim, S.W.; Paick, J.S. New unstable bladder model in hypercholesterolemia rats. Urology 2007, 69, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Kudoh, J.; Masunaga, K.; Nagata, T.; Shiomi, M. Effects of Chronic Hyperlipidemia on Lower Urinary Tract Function-Bladder Dysfunction in Myocardial Infarction-Prone Watanabe Heritable Hyperlipidemic Rabbits. Low. Urin. Tract. Symptoms 2012, 4 (Suppl. S1), 21–26. [Google Scholar] [CrossRef] [PubMed]
- Velasquez Flores, M.; Mossa, A.H.; Cammisotto, P.; Campeau, L. Bladder overdistension with polyuria in a hypertensive rat model. Neurourol. Urodyn. 2018, 37, 1904–1912. [Google Scholar] [CrossRef]
- Shimizu, S.; Nagao, Y.; Kurabayashi, A.; Shimizu, T.; Higashi, Y.; Karashima, T.; Saito, M. Aging-related severe hypertension induces detrusor underactivity in rats. Life Sci. 2021, 283, 119855. [Google Scholar] [CrossRef] [PubMed]
- Langdale, C.L.; Degoski, D.; Milliken, P.H.; Grill, W.M. Voiding behavior in awake unrestrained untethered spontaneously hypertensive and Wistar control rats. Am. J. Physiol. Renal. Physiol. 2021, 321, F195–F206. [Google Scholar] [CrossRef]
- Lee, R.K.; Chung, D.; Chughtai, B.; Te, A.E.; Kaplan, S.A. Central obesity as measured by waist circumference is predictive of severity of lower urinary tract symptoms. BJU Int. 2012, 110, 540–545. [Google Scholar] [CrossRef]
- Pashootan, P.; Ploussard, G.; Cocaul, A.; de Gouvello, A.; Desgrandchamps, F. Association between metabolic syndrome and severity of lower urinary tract symptoms (LUTS): An observational study in a 4666 European men cohort. BJU Int. 2015, 116, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Al Hussein Alawamlh, O.; Al Hussein Al Awamlh, B.; Lee, R.K. Metabolic syndrome and voiding dysfunction. Curr. Opin. Urol. 2017, 27, 403–408. [Google Scholar] [CrossRef]
- Hsu, L.N.; Hu, J.C.; Chen, P.Y.; Lee, W.C.; Chuang, Y.C. Metabolic Syndrome and Overactive Bladder Syndrome May Share Common Pathophysiologies. Biomedicines 2022, 10, 1957. [Google Scholar] [CrossRef]
- Saratlija Novakovic, Z.; Tesija, R.A.; Puljak, L. Association between metabolic syndrome and overactive bladder: A case-control study. Scand. J. Urol. 2017, 51, 470–473. [Google Scholar] [CrossRef]
- Kurokawa, T.; Zha, X.; Ito, H.; Aoki, Y.; Akino, H.; Kobayashi, M.; Yokoyama, O. Underlying mechanisms of urine storage dysfunction in rats with salt-loading hypertension. Life Sci. 2015, 141, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Mossa, A.H.; Velasquez Flores, M.; Cammisotto, P.G.; Campeau, L. Succinate, increased in metabolic syndrome, activates GPR91 receptor signaling in urothelial cells. Cell Signal 2017, 37, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Rose’meyer, R.; McDermott, C.; Chess-Williams, R.; Sellers, D.J. Diabetes-induced alterations in urothelium function: Enhanced ATP release and nerve-evoked contractions in the streptozotocin rat bladder. Clin. Exp. Pharmacol. Physiol. 2018, 45, 1161–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, E.J.; Odom, M.R.; Hannan, J.L.; Grill, W.M. Dysfunctional voiding behavior and impaired muscle contractility in a rat model of detrusor underactivity. Neurourol. Urodyn. 2021, 40, 1889–1899. [Google Scholar] [CrossRef]
- Yu, W.; Robson, S.C.; Hill, W.G. Expression and distribution of ectonucleotidases in mouse urinary bladder. PLoS ONE 2011, 6, e18704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, W. Polarized ATP distribution in urothelial mucosal and serosal space is differentially regulated by stretch and ectonucleotidases. Am. J. Physiol. Renal Physiol. 2015, 309, F864–F872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aresta Branco, M.S.L.; Gutierrez Cruz, A.; Dayton, J.; Perrino, B.A.; Mutafova-Yambolieva, V.N. Mechanosensitive Hydrolysis of ATP and ADP in Lamina Propria of the Murine Bladder by Membrane-Bound and Soluble Nucleotidases. Front. Physiol. 2022, 13, 918100. [Google Scholar] [CrossRef]
- McCloskey, K.D. The detrusor-free bladder-it can still hold its water. J. Physiol. 2018, 10, 1427–1428. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, H.; Zebisch, M.; Strater, N. Cellular function and molecular structure of ecto-nucleotidases. Purinergic Signal. 2012, 8, 437–502. [Google Scholar]
- Todorov, L.D.; Mihaylova-Todorova, S.; Westfall, T.D.; Sneddon, P.; Kennedy, C.; Bjur, R.A.; Westfall, D.P. Neuronal release of soluble nucleotidases and their role in neurotransmitter inactivation. Nature 1997, 387, 76–79. [Google Scholar] [CrossRef]
- Vekaria, R.M.; Unwin, R.J.; Shirley, D.G. Intraluminal ATP concentrations in rat renal tubules. J. Am. Soc. Nephrol. 2006, 17, 1841–1847. [Google Scholar] [CrossRef]
- Lundin, A.; Hallander, H.; Kallner, A.; Lundin, U.K.; Osterberg, E. Bacteriuria testing by the ATP method as an integral part in the diagnosis and therapy of urinary tract infection (UTI). J. Biolumin. Chemilumin. 1989, 4, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Durnin, L.; Corrigan, R.D.; Sanders, K.M.; Mutafova-Yambolieva, V.N. A Decentralized (Ex Vivo) Murine Bladder Model with the Detrusor Muscle Removed for Direct Access to the Suburothelium during Bladder Filling. J. Vis. Exp. 2019, e60344. [Google Scholar] [CrossRef]
- Levitt, B.; Head, R.J.; Westfall, D.P. High-pressure liquid chromatographic-fluorometric detection of adenosine and adenine nucleotides: Application to endogenous content and electrically induced release of adenyl purines in guinea pig vas deferens. Anal. Biochem. 1984, 137, 93–100. [Google Scholar] [CrossRef]
- Bobalova, J.; Bobal, P.; Mutafova-Yambolieva, V.N. High-Performance Liquid Chromatographic Technique for Detection of a Fluorescent Analogue of ADP-Ribose in Isolated Blood Vessel Preparations. Anal. Biochem. 2002, 305, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sasse, K.C.; Bayguinov, Y.; Ward, S.M.; Perrino, B.A. Contractile Protein Expression and Phosphorylation and Contractility of Gastric Smooth Muscles from Obese Patients and Patients with Obesity and Diabetes. J. Diabetes Res. 2018, 2018, 8743874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Han, K.H.; Grainger, N.; Li, W.; Corrigan, R.D.; Perrino, B.A. A role for focal adhesion kinase in facilitating the contractile responses of murine gastric fundus smooth muscles. J. Physiol. 2018, 596, 2131–2146. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Zacharia, L.C.; Jackson, E.K.; Apodaca, G. Adenosine receptor expression and function in bladder uroepithelium. Am. J. Physiol. Cell Physiol. 2006, 291, C254–C265. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic signalling in the urinary tract in health and disease. Purinergic Signal. 2014, 10, 103–155. [Google Scholar] [CrossRef] [Green Version]
- Takezawa, K.; Kondo, M.; Kiuchi, H.; Ueda, N.; Soda, T.; Fukuhara, S.; Takao, T.; Miyagawa, Y.; Tsujimura, A.; Matsumoto-Miyai, K.; et al. Authentic role of ATP signaling in micturition reflex. Sci. Rep. 2016, 6, 19585. [Google Scholar] [CrossRef] [Green Version]
- Pandita, R.K.; Andersson, K.E. Intravesical adenosine triphosphate stimulates the micturition reflex in awake, freely moving rats. J. Urol. 2002, 168, 1230–1234. [Google Scholar] [CrossRef] [PubMed]
- Ueda, N.; Kondo, M.; Takezawa, K.; Kiuchi, H.; Sekii, Y.; Inagaki, Y.; Soda, T.; Fukuhara, S.; Fujita, K.; Uemura, M.; et al. Intravesical ATP instillation induces urinary frequency because of activation of bladder afferent nerves without inflammatory changes in mice: A promising model for overactive bladder. Biochem. Biophys. Res. Commun. 2018, 506, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Durnin, L.; Hwang, S.J.; Ward, S.M.; Sanders, K.M.; Mutafova-Yambolieva, V.N. Adenosine 5’-diphosphate-ribose (ADPR) is a neural regulator in primate and murine large intestine along with beta-NAD+. J. Physiol. 2012, 590, 1921–1941. [Google Scholar] [CrossRef] [Green Version]
- Durnin, L.; Hwang, S.J.; Kurahashi, M.; Drumm, B.T.; Ward, S.M.; Sasse, K.C.; Sanders, K.M.; Mutafova-Yambolieva, V.N. Uridine adenosine tetraphosphate is a novel neurogenic P2Y1 receptor activator in the gut. Proc. Natl. Acad. Sci. USA 2014, 111, 15821–15826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durnin, L.; Kurahashi, M.; Sanders, K.M.; Mutafova-Yambolieva, V.N. Extracellular metabolism of the enteric inhibitory neurotransmitter beta-nicotinamide adenine dinucleotide (beta-NAD) in the murine colon. J. Physiol. 2020, 598, 4509–4521. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, I.; Timóteo, M.A.; Silva, I.; Vieira, C.; Baldaia, C.; Ferreirinha, F.; Silva-Ramos, M.; Correia-de-Sá, P. Activation of P2Y6 receptors increases the voiding frequency in anaesthetized rats by releasing ATP from the bladder urothelium. Br. J. Pharmacol. 2014, 171, 3404–3419. [Google Scholar] [CrossRef] [Green Version]
- Rockenbach, L.; Braganhol, E.; Dietrich, F.; Figueiró, F.; Pugliese, M.; Edelweiss, M.I.; Morrone, F.B.; Sévigny, J.; Battastini, A.M. NTPDase3 and ecto-5’-nucleotidase/CD73 are differentially expressed during mouse bladder cancer progression. Purinergic Signal. 2014, 10, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Firouzmand, S.; Ajori, L.; Towse, J.; Allameh, F.; Najafi, S.; Javed, S.; John, B.; Langley, S.E.M.; Fry, C.H.; Young, J.S. Investigating the associations of mucosal P2Y6 receptor expression and urinary ATP and ADP concentrations, with symptoms of overactive bladder. Neurourol. Urodyn. 2020, 39, 926–934. [Google Scholar] [CrossRef]
- Lewis, S.A.; Lewis, J.R. Kinetics of urothelial ATP release. Am. J. Physiol. Renal Physiol. 2006, 291, F332–F340. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.Q.; Heldman, M.R.; Herrmann, M.A.; Kedei, N.; Woo, W.; Blumberg, P.M.; Goldsmith, P.K. Absolute quantitation of endogenous proteins with precision and accuracy using a capillary Western system. Anal. Biochem. 2013, 442, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Allred, C.C.; Jensen, M.D. Human adipose tissue protein analyses using capillary western blot technology. Nutr. Diabetes 2018, 8, 26. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.G.; Wu, Y.; Csizmadia, E.; Feldbrügge, L.; Enjyoji, K.; Tigges, J.; Toxavidis, V.; Stephan, H.; Müller, C.E.; McKnight, C.J.; et al. Characterization of circulating microparticle-associated CD39 family ecto-nucleotidases in human plasma. Purinergic Signal. 2014, 10, 611–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, S.F.; Lemberg, M.K.; Fluhrer, R. Proteolytic ectodomain shedding of membrane proteins in mammals-hardware, concepts, and recent developments. EMBO. J. 2018, 37, e99456. [Google Scholar] [CrossRef]
- Barat, C.; Martin, G.; Beaudoin, A.R.; Sévigny, J.; Tremblay, M.J. The nucleoside triphosphate diphosphohydrolase-1/CD39 is incorporated into human immunodeficiency type 1 particles, where it remains biologically active. J. Mol. Biol. 2007, 371, 269–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcedo, K.P.; Bowser, J.L.; Snider, N.T. The elegant complexity of mammalian ecto-5’-nucleotidase (CD73). Trends. Cell Biol. 2021, 31, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H. History of ectonucleotidases and their role in purinergic signaling. Biochem. Pharmacol. 2021, 187, 114322. [Google Scholar] [CrossRef]
- Street, S.E.; Kramer, N.J.; Walsh, P.L.; Taylor-Blake, B.; Yadav, M.C.; King, I.F.; Vihko, P.; Wightman, R.M.; Millán, J.L.; Zylka, M.J. Tissue-nonspecific alkaline phosphatase acts redundantly with PAP and NT5E to generate adenosine in the dorsal spinal cord. J. Neurosci. 2013, 33, 11314–11322. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Copeland, J.; Shin, M.; Chang, Y.; Venton, B.J. CD73 or CD39 Deletion Reveals Different Mechanisms of Formation for Spontaneous and Mechanically Stimulated Adenosine and Sex Specific Compensations in ATP Degradation. ACS Chem. Neurosci. 2020, 11, 919–928. [Google Scholar] [CrossRef]
- Jackson, E.K.; Gillespie, D.G.; Cheng, D.; Mi, Z.; Menshikova, E.V. Characterization of the N(6)-etheno-bridge method to assess extracellular metabolism of adenine nucleotides: Detection of a possible role for purine nucleoside phosphorylase in adenosine metabolism. Purinergic Signal. 2020, 16, 187–211. [Google Scholar] [CrossRef]
- Hudoklin, S.; Jezernik, K.; Neumüller, J.; Pavelka, M.; Romih, R. Electron tomography of fusiform vesicles and their organization in urothelial cells. PLoS One 2012, 7, e32935. [Google Scholar] [CrossRef] [Green Version]
- Zupančič, D.; Romih, R.; Robenek, H.; Žužek Rožman, K.; Samardžija, Z.; Kostanjšek, R.; Kreft, M.E. Molecular ultrastructure of the urothelial surface: Insights from a combination of various microscopic techniques. Microsc. Res. Tech. 2014, 77, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.A.; de Moura, J.L. Incorporation of cytoplasmic vesicles into apical membrane of mammalian urinary bladder epithelium. Nature 1982, 297, 685–688. [Google Scholar] [CrossRef]
- Dalghi, M.G.; Montalbetti, N.; Carattino, M.D.; Apodaca, G. The Urothelium: Life in a Liquid Environment. Physiol. Rev. 2020, 100, 1621–1705. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.M.; Ding, M.; Lavker, R.M.; Sun, T.T. Urothelial function reconsidered: A role in urinary protein secretion. Proc. Natl. Acad. Sci. USA 2001, 98, 154–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, C.; Lavriha, P.; Mehta, V.; Khanppnavar, B.; Mohammed, I.; Li, Y.; Lazaratos, M.; Schaefer, J.V.; Dreier, B.; Plückthun, A.; et al. Structural basis of adenylyl cyclase 9 activation. Nat. Commun. 2022, 13, 1045. [Google Scholar] [CrossRef] [PubMed]
- Prakasam, H.S.; Gallo, L.I.; Li, H.; Ruiz, W.G.; Hallows, K.R.; Apodaca, G. A1 adenosine receptor-stimulated exocytosis in bladder umbrella cells requires phosphorylation of ADAM17 Ser-811 and EGF receptor transactivation. Mol. Biol. Cell 2014, 25, 3798–3812. [Google Scholar] [CrossRef] [PubMed]
- Prakasam, H.S.; Herrington, H.; Roppolo, J.R.; Jackson, E.K.; Apodaca, G. Modulation of bladder function by luminal adenosine turnover and A1 receptor activation. Am. J. Physiol. Renal Physiol. 2012, 303, F279–F292. [Google Scholar] [CrossRef] [Green Version]
- Dunning-Davies, B.M.; Fry, C.H.; Mansour, D.; Ferguson, D.R. The regulation of ATP release from the urothelium by adenosine and transepithelial potential. BJU. Int. 2013, 111, 505–513. [Google Scholar] [CrossRef]
- Kitta, T.; Chancellor, M.B.; de Groat, W.C.; Kuno, S.; Nonomura, K.; Yoshimura, N. Roles of adenosine A1 and A2A receptors in the control of micturition in rats. Neurourol. Urodyn. 2014, 33, 1259–1265. [Google Scholar] [CrossRef]
- Winder, M.; Tobin, G.; Zupancic, D.; Romih, R. Signalling molecules in the urothelium. Biomed. Res. Int. 2014, 2014, 297295. [Google Scholar] [CrossRef] [Green Version]
- Merrill, L.; Gonzalez, E.J.; Girard, B.M.; Vizzard, M.A. Receptors, channels, and signalling in the urothelial sensory system in the bladder. Nat. Rev. Urol. 2016, 13, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Peleli, M.; Fredholm, B.B.; Sobrevia, L.; Carlström, M. Pharmacological targeting of adenosine receptor signaling. Mol. Aspects Med. 2017, 55, 4–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, R.A.; Skennerton, D.E.; Newgreen, D.; Fry, C.H. The contractile potency of adenosine triphosphate and ecto-adenosine triphosphatase activity in guinea pig detrusor and detrusor from patients with a stable, unstable or obstructed bladder. J. Urol. 2002, 168, 1235–1239. [Google Scholar] [CrossRef]
- Takezawa, K.; Kondo, M.; Nonomura, N.; Shimada, S. Urothelial ATP signaling: What is its role in bladder sensation? Neurourol. Urodyn. 2016, 36, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Gill, K.; Horsley, H.; Kupelian, A.S.; Baio, G.; De Iorio, M.; Sathiananamoorthy, S.; Khasriya, R.; Rohn, J.L.; Wildman, S.S.; Malone-Lee, J. Urinary ATP as an indicator of infection and inflammation of the urinary tract in patients with lower urinary tract symptoms. BMC Urol. 2015, 15, 7. [Google Scholar] [CrossRef] [PubMed]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutierrez Cruz, A.; Aresta Branco, M.S.L.; Perrino, B.A.; Sanders, K.M.; Mutafova-Yambolieva, V.N. Urinary ATP Levels Are Controlled by Nucleotidases Released from the Urothelium in a Regulated Manner. Metabolites 2023, 13, 30. https://doi.org/10.3390/metabo13010030
Gutierrez Cruz A, Aresta Branco MSL, Perrino BA, Sanders KM, Mutafova-Yambolieva VN. Urinary ATP Levels Are Controlled by Nucleotidases Released from the Urothelium in a Regulated Manner. Metabolites. 2023; 13(1):30. https://doi.org/10.3390/metabo13010030
Chicago/Turabian StyleGutierrez Cruz, Alejandro, Mafalda S. L. Aresta Branco, Brian A. Perrino, Kenton M. Sanders, and Violeta N. Mutafova-Yambolieva. 2023. "Urinary ATP Levels Are Controlled by Nucleotidases Released from the Urothelium in a Regulated Manner" Metabolites 13, no. 1: 30. https://doi.org/10.3390/metabo13010030