
Seleno-Metabolites and Their Precursors: A New Dawn for Several Illnesses?

 , , , and
, , , and 
Abstract
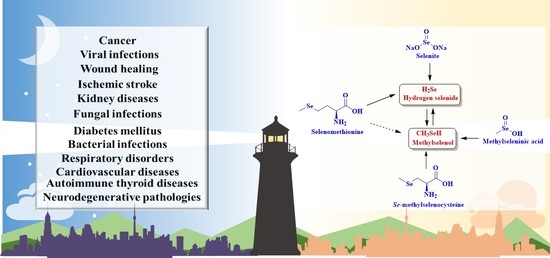
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
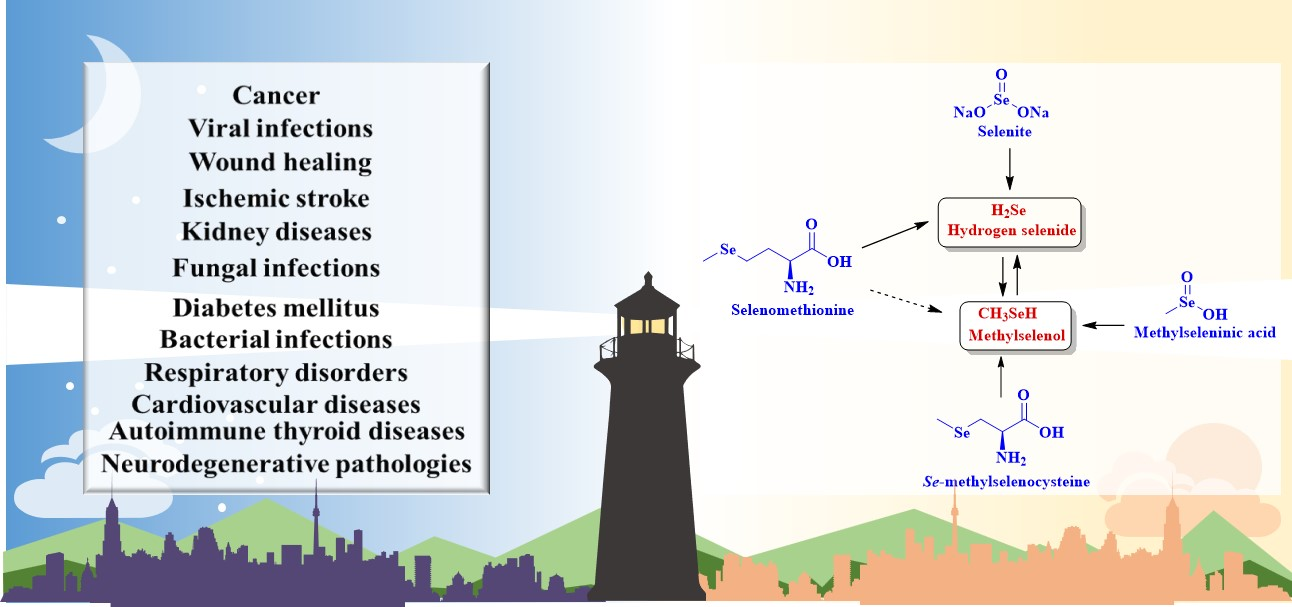
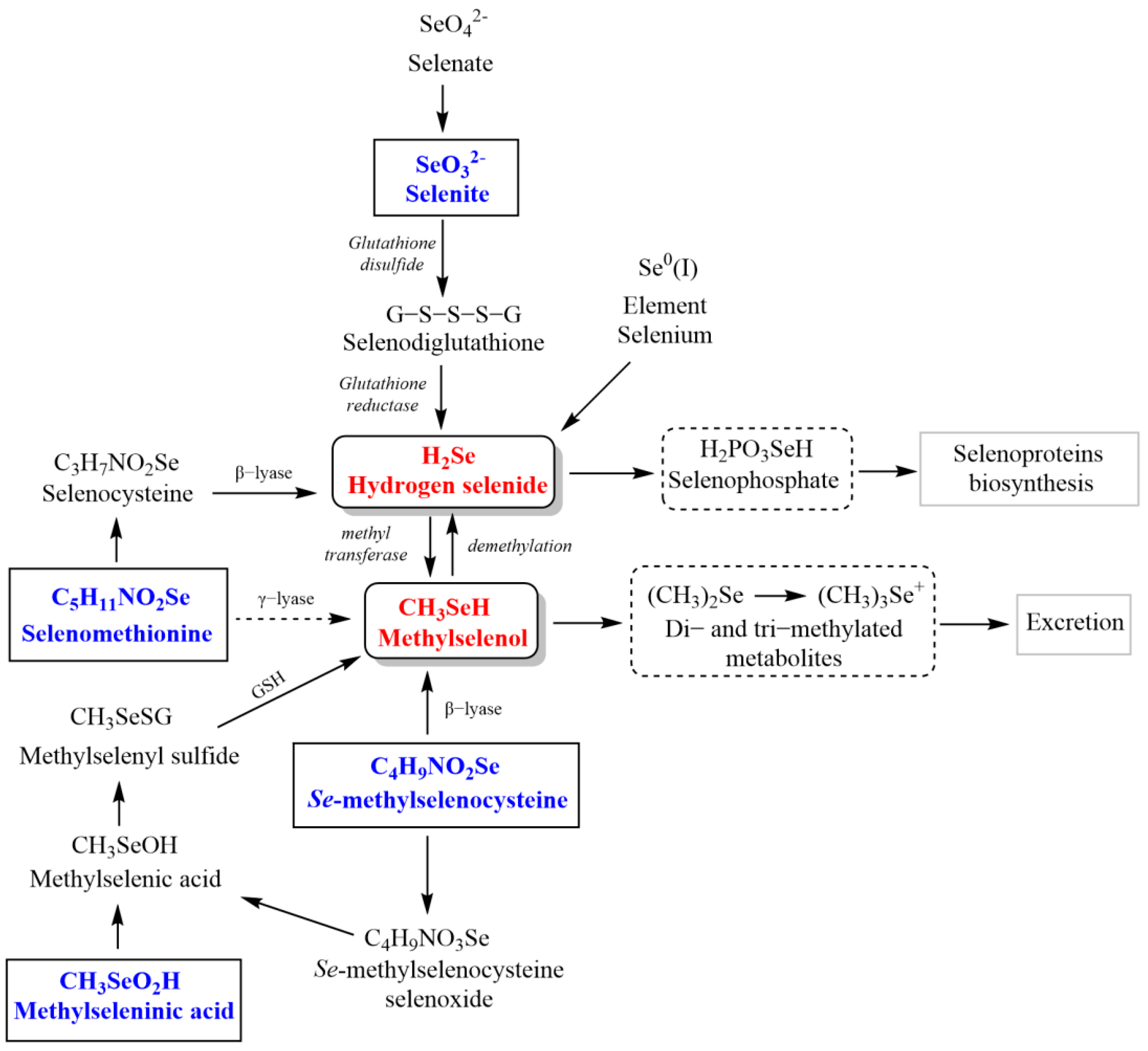
2. Hydrogen Selenide Precursors
2.1. Sodium Selenite
2.1.1. Antibacterial Activity of Sodium Selenite
2.1.2. Sodium Selenite and Asthma
2.1.3. Sodium Selenite and Cancer
2.1.4. Sodium Selenite and Cardiovascular Diseases
2.1.5. Sodium Selenite and Diabetes
2.1.6. Sodium Selenite and Neurodegenerative Diseases
2.1.7. Sodium Selenite and Viral Infections
3. Methylselenol Precursors
3.1. Methylseleninic Acid
3.1.1. Methylseleninic Acid and Cancer
3.1.2. Methylseleninic Acid and Endometritis
3.1.3. Methylseleninic Acid and COVID-19
3.1.4. Methylseleninic Acid and Mycobacterium tuberculosis Infection
3.2. Se-methylselenocysteine
3.2.1. Se-methylselenocysteine and Alzheimer’s Disease
3.2.2. Se-methylselenocysteine and Cancer
3.2.3. Se-methylselenocysteine and Vulvar Candidiasis
3.2.4. Se-methylselenocysteine and Ischemic Stroke
3.2.5. Se-methylselenocysteine and Wound Healing
3.3. Selenomethionine
3.3.1. Selenomethionine and Autoimmune Thyroid Diseases
3.3.2. Selenomethionine and Cancer
3.3.3. Selenomethionine and Cardiovascular Diseases
3.3.4. Selenomethionine and Intestinal Ischemia-Reperfusion Injury
3.3.5. Selenomethionine and Kidney Diseases
3.3.6. Selenomethionine and Neurodegenerative Pathologies
3.3.7. Selenomethionine and Viral Infections
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kieliszek, M. Selenium–fascinating microelement, properties and sources in food. Molecules 2019, 24, 1298. [Google Scholar] [CrossRef] [PubMed]
- Santi, C.; Bagnoli, L. Celebrating two centuries of research in selenium chemistry: State of the art and new prospective. Molecules 2017, 22, 2124. [Google Scholar] [CrossRef] [PubMed]
- Avery, J.C.; Hoffmann, P.R. Selenium, selenoproteins, and immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium intake, status, and health: A complex relationship. Hormones 2020, 19, 9–14. [Google Scholar] [CrossRef]
- Barchielli, G.; Capperucci, A.; Tanini, D. The role of selenium in pathologies: An updated review. Antioxidants 2022, 11, 251. [Google Scholar] [CrossRef]
- Kuršvietienė, L.; Mongirdienė, A.; Bernatonienė, J.; Šulinskienė, J.; Stanevičienė, I. Selenium anticancer properties and impact on cellular redox status. Antioxidants 2020, 9, 80. [Google Scholar] [CrossRef]
- Radomska, D.; Czarnomysy, R.; Radomski, D.; Bielawska, A.; Bielawski, K. Selenium as a bioactive micronutrient in the human diet and its cancer chemopreventive activity. Nutrients 2021, 13, 1649. [Google Scholar] [CrossRef]
- Dos Santos, M.; da Silva Júnior, F.M.R.; Muccillo-Baisch, A.L. Selenium content of brazilian foods: A review of the literature values. J. Food Compost. Anal. 2017, 58, 10–15. [Google Scholar] [CrossRef]
- Ferreira, R.L.U.; Sena-Evangelista, K.C.M.; de Azevedo, E.P.; Pinheiro, F.I.; Cobucci, R.N.; Pedrosa, L.F.C. Selenium in human health and gut microflora: Bioavailability of selenocompounds and relationship with diseases. Front. Nutr. 2021, 8, 685317. [Google Scholar] [CrossRef]
- Chuai, H.; Zhang, S.-Q.; Bai, H.; Li, J.; Wang, Y.; Sun, J.; Wen, E.; Zhang, J.; Xin, M. Small molecule selenium-containing compounds: Recent development and therapeutic applications. Eur. J. Med. Chem. 2021, 223, 113621. [Google Scholar] [CrossRef]
- Yang, R.; Liu, Y.; Zhou, Z. Selenium and selenoproteins, from structure, function to food resource and nutrition. Food Sci. Technol. Res. 2017, 23, 363–373. [Google Scholar] [CrossRef]
- Tsuji, P.A.; Santesmasses, D.; Lee, B.J.; Gladyshev, V.N.; Hatfield, D.L. Historical roles of selenium and selenoproteins in health and development: The good, the bad and the ugly. Int. J. Mol. Sci. 2021, 23, 5. [Google Scholar] [CrossRef]
- Hariharan, S.; Dharmaraj, S. Selenium and selenoproteins: It’s role in regulation of inflammation. Inflammopharmacology 2020, 28, 667–695. [Google Scholar] [CrossRef]
- Qazi, I.H.; Angel, C.; Yang, H.; Zoidis, E.; Pan, B.; Wu, Z.; Ming, Z.; Zeng, C.J.; Meng, Q.; Han, H.; et al. Role of selenium and selenoproteins in male reproductive function: A review of past and present evidences. Antioxidants 2019, 8, 268. [Google Scholar] [CrossRef]
- Arias-Borrego, A.; Callejón-Leblic, B.; Calatayud, M.; Gómez-Ariza, J.L.; Collado, M.C.; García-Barrera, T. Insights into cancer and neurodegenerative diseases through selenoproteins and the connection with gut microbiota—Current analytical methodologies. Expert Rev. Proteom. 2019, 16, 805–814. [Google Scholar] [CrossRef]
- Qazi, I.H.; Angel, C.; Yang, H.; Pan, B.; Zoidis, E.; Zeng, C.-J.; Han, H.; Zhou, G.-B. Selenium, selenoproteins, and female reproduction: A review. Molecules 2018, 23, 3053. [Google Scholar] [CrossRef]
- Barrett, C.W.; Short, S.P.; Williams, C.S. Selenoproteins and oxidative stress-induced inflammatory tumorigenesis in the gut. Cell. Mol. Life Sci. 2017, 74, 607–616. [Google Scholar] [CrossRef]
- Jehan, C.; Cartier, D.; Bucharles, C.; Anouar, Y.; Lihrmann, I. Emerging roles of ER-resident selenoproteins in brain physiology and physiopathology. Redox Biol. 2022, 55, 102412. [Google Scholar] [CrossRef]
- Koeberle, S.C.; Gollowitzer, A.; Laoukili, J.; Kranenburg, O.; Werz, O.; Koeberle, A.; Kipp, A.P. Distinct and overlapping functions of glutathione peroxidases 1 and 2 in limiting NF-κB-driven inflammation through redox-active mechanisms. Redox Biol. 2020, 28, 101388. [Google Scholar] [CrossRef]
- Lee, J.H.; Jang, J.K.; Ko, K.Y.; Jin, Y.; Ham, M.; Kang, H.; Kim, I.Y. Degradation of selenoprotein S and selenoprotein K through PPARγ-mediated ubiquitination is required for adipocyte differentiation. Cell Death Differ. 2019, 26, 1007–1023. [Google Scholar] [CrossRef]
- Bevinakoppamath, S.; Saleh Ahmed, A.M.; Ramachandra, S.C.; Vishwanath, P.; Prashant, A. Chemopreventive and anticancer property of selenoproteins in obese breast cancer. Front. Pharmacol. 2021, 12, 618172. [Google Scholar] [CrossRef] [PubMed]
- Lennicke, C.; Rahn, J.; Bukur, J.; Hochgräfe, F.; Wessjohann, L.A.; Lichtenfels, R.; Seliger, B. Modulation of MHC class I surface expression in B16F10 melanoma cells by methylseleninic acid. OncoImmunology 2017, 6, e1259049. [Google Scholar] [CrossRef] [PubMed]
- Poluboyarinov, P.A.; Elistratov, D.G.; Moiseeva, I.J. Antitumor activity of selenium and search parameters for its new potentially active derivatives. Russ. J. Bioorg. Chem. 2020, 46, 989–1003. [Google Scholar] [CrossRef]
- Mal’tseva, V.N.; Goltyaev, M.V.; Turovsky, E.A.; Varlamova, E.G. Immunomodulatory and anti-inflammatory properties of selenium-containing agents: Their role in the regulation of defense mechanisms against COVID-19. Int. J. Mol. Sci. 2022, 23, 2360. [Google Scholar] [CrossRef]
- Kang, X.; Huang, H.; Jiang, C.; Cheng, L.; Sang, Y.; Cai, X.; Dong, Y.; Sun, L.; Wen, X.; Xi, Z.; et al. Cysteine-activated small-molecule H2Se donors inspired by synthetic H2S donors. J. Am. Chem. Soc. 2022, 144, 3957–3967. [Google Scholar] [CrossRef]
- Newton, T.D.; Bolton, S.G.; Garcia, A.C.; Chouinard, J.E.; Golledge, S.L.; Zakharov, L.N.; Pluth, M.D. Hydrolysis-based small-molecule hydrogen selenide (H2Se) donors for intracellular H2Se delivery. J. Am. Chem. Soc. 2021, 143, 19542–19550. [Google Scholar] [CrossRef]
- Luan, D.; Zhao, Z.; Xia, D.; Zheng, Q.; Gao, X.; Xu, K.; Tang, B. Hydrogen selenide, a vital metabolite of sodium selenite, uncouples the sulfilimine bond and promotes the reversal of liver fibrosis. Sci. China Life Sci. 2021, 64, 443–451. [Google Scholar] [CrossRef]
- Kuganesan, M.; Samra, K.; Evans, E.; Singer, M.; Dyson, A. Selenium and hydrogen selenide: Essential micronutrient and the fourth gasotransmitter? Intensive Care Med. Exp. 2019, 7, 71. [Google Scholar] [CrossRef]
- Hartmann, C.; Nussbaum, B.; Calzia, E.; Radermacher, P.; Wepler, M. Gaseous mediators and mitochondrial function: The future of pharmacologically induced suspended animation? Front. Physiol. 2017, 8, 691. [Google Scholar] [CrossRef]
- Samra, K.; Kuganesan, M.; Smith, W.; Kleyman, A.; Tidswell, R.; Arulkumaran, N.; Singer, M.; Dyson, A. The pharmacology and therapeutic utility of sodium hydroselenide. Int. J. Mol. Sci. 2021, 22, 3258. [Google Scholar] [CrossRef]
- Kong, F.; Ge, L.; Pan, X.; Xu, K.; Liu, X.; Tang, B. A highly selective near-infrared fluorescent probe for imaging H2Se in living cells and in vivo. Chem. Sci. 2016, 7, 1051–1056. [Google Scholar] [CrossRef]
- Kong, F.; Zhao, Y.; Liang, Z.; Liu, X.; Pan, X.; Luan, D.; Xu, K.; Tang, B. Highly selective fluorescent probe for imaging H2Se in living cells and in vivo based on the disulfide bond. Anal. Chem. 2017, 89, 688–693. [Google Scholar] [CrossRef]
- Pan, X.; Song, X.; Wang, C.; Cheng, T.; Luan, D.; Xu, K.; Tang, B. H2Se induces reductive stress in HepG2 cells and activates cell autophagy by regulating the redox of HMGB1 protein under hypoxia. Theranostics 2019, 9, 1794–1808. [Google Scholar] [CrossRef]
- Kim, S.J.; Choi, M.C.; Park, J.M.; Chung, A.S. Antitumor effects of selenium. Int. J. Mol. Sci. 2021, 22, 11844. [Google Scholar] [CrossRef]
- Hagemann-Jensen, M.; Uhlenbrock, F.; Kehlet, S.; Andresen, L.; Gabel-Jensen, C.; Ellgaard, L.; Gammelgaard, B.; Skov, S. The selenium metabolite methylselenol regulates the expression of ligands that trigger immune activation through the lymphocyte receptor NKG2D. J. Biol. Chem. 2014, 289, 31576–31590. [Google Scholar] [CrossRef]
- Dauplais, M.; Bierla, K.; Maizeray, C.; Lestini, R.; Lobinski, R.; Plateau, P.; Szpunar, J.; Lazard, M. Methylselenol produced in vivo from methylseleninic acid or dimethyl diselenide induces toxic protein aggregation in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2021, 22, 2241. [Google Scholar] [CrossRef]
- Díaz-Argelich, N.; Encío, I.; Plano, D.; Fernandes, A.P.; Palop, J.A.; Sanmartín, C. Novel methylselenoesters as antiproliferative agents. Molecules 2017, 22, 1288. [Google Scholar] [CrossRef]
- Dauplais, M.; Mahou, P.; Plateau, P.; Lazard, M. Exposure to the methylselenol precursor dimethyldiselenide induces a reductive endoplasmic reticulum stress in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2021, 22, 5467. [Google Scholar] [CrossRef]
- Ebert, F.; Müller, S.M.; Meyer, S.; Schwerdtle, T. Selenium and toxicological aspects: Cytotoxicity, cellular bioavailability, and biotransformation of Se species. In Selenium; Michalke, B., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 373–391. [Google Scholar]
- Font, M.; Romano, B.; González-Peñas, E.; Sanmartín, C.; Plano, D.; Palop, J.A. Methylselenol release as a cytotoxic tool: A study of the mechanism of the activity achieved by two series of methylselenocarbamate derivatives. Metallomics 2018, 10, 1128–1140. [Google Scholar] [CrossRef]
- Kieliszek, M.; Lipinski, B.; Blazejak, S. Application of sodium selenite in the prevention and treatment of cancers. Cells 2017, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Hiffler, L.; Rakotoambinina, B. Selenium and RNA virus interactions: Potential implications for SARS-CoV-2 infection (COVID-19). Front. Nutr. 2020, 7, 164. [Google Scholar] [CrossRef]
- Varela, M.F.; Stephen, J.; Lekshmi, M.; Ojha, M.; Wenzel, N.; Sanford, L.M.; Hernandez, A.J.; Parvathi, A.; Kumar, S.H. Bacterial resistance to antimicrobial agents. Antibiotics 2021, 10, 593. [Google Scholar] [CrossRef]
- Lai, C.-C.; Chen, S.-Y.; Ko, W.-C.; Hsueh, P.-R. Increased antimicrobial resistance during the COVID-19 pandemic. Int. J. Antimicrob. Agents 2021, 57, 106324. [Google Scholar] [CrossRef]
- Rodríguez-Baño, J.; Rossolini, G.M.; Schultsz, C.; Tacconelli, E.; Murthy, S.; Ohmagari, N.; Holmes, A.; Bachmann, T.; Goossens, H.; Canton, R.; et al. Key considerations on the potential impacts of the COVID-19 pandemic on antimicrobial resistance research and surveillance. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 1122–1129. [Google Scholar] [CrossRef]
- Hu, X.; Guo, J.; Xu, M.; Jiang, P.; Yuan, X.; Zhao, C.; Maimai, T.; Cao, Y.; Zhang, N.; Fu, Y. Clostridium tyrobutyricum alleviates Staphylococcus aureus-induced endometritis in mice by inhibiting endometrial barrier disruption and inflammatory response. Food Funct. 2019, 10, 6699–6710. [Google Scholar] [CrossRef]
- Pellissery, A.J.; Vinayamohan, P.G.; Yin, H.B.; Mooyottu, S.; Venkitanarayanan, K. In vitro efficacy of sodium selenite in reducing toxin production, spore outgrowth and antibiotic resistance in hypervirulent Clostridium difficile. J. Med. Microbiol. 2019, 68, 1118–1128. [Google Scholar] [CrossRef]
- Mosolygo, T.; Kincses, A.; Csonka, A.; Tonki, A.S.; Witek, K.; Sanmartin, C.; Marc, M.A.; Handzlik, J.; Kiec-Kononowicz, K.; Dominguez-Alvarez, E.; et al. Selenocompounds as novel antibacterial agents and bacterial efflux pump inhibitors. Molecules 2019, 24, 1487. [Google Scholar] [CrossRef]
- Kumar, B.S.; Tiwari, S.K.; Manoj, G.; Kunwar, A.; Amrita, N.; Sivaram, G.; Abid, Z.; Ahmad, A.; Khan, A.A.; Priyadarsini, K.I. Anti-unlcer and antimicrobial activities of sodium selenite against Helicobacter pylori: In vitro and in vivo evaluation. Scand. J. Infect. Dis. 2010, 42, 266–274. [Google Scholar] [CrossRef]
- Boonpiyathad, T.; Sözener, Z.C.; Satitsuksanoa, P.; Akdis, C.A. Immunologic mechanisms in asthma. Semin. Immunol. 2019, 46, 101333. [Google Scholar] [CrossRef]
- Allam, V.; Paudel, K.R.; Gupta, G.; Singh, S.K.; Vishwas, S.; Gulati, M.; Gupta, S.; Chaitanya, M.; Jha, N.K.; Gupta, P.K.; et al. Nutraceuticals and mitochondrial oxidative stress: Bridging the gap in the management of bronchial asthma. Environ. Sci. Pollut. Res. Int. 2022, 29, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Ammar, M.; Bahloul, N.; Amri, O.; Omri, R.; Ghozzi, H.; Kammoun, S.; Zeghal, K.; Ben Mahmoud, L. Oxidative stress in patients with asthma and its relation to uncontrolled asthma. J. Clin. Lab. Anal. 2022, 36, e24345. [Google Scholar] [CrossRef] [PubMed]
- Karadogan, B.; Beyaz, S.; Gelincik, A.; Buyukozturk, S.; Arda, N. Evaluation of oxidative stress biomarkers and antioxidant parameters in allergic asthma patients with different level of asthma control. J. Asthma 2022, 59, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, S.O.; Newson, R.B.; Rayman, M.P.; Wong, A.P.; Tumilty, M.K.; Phillips, J.M.; Potts, J.F.; Kelly, F.J.; White, P.T.; Burney, P.G. Randomised, double blind, placebo-controlled trial of selenium supplementation in adult asthma. Thorax 2007, 62, 483–490. [Google Scholar] [CrossRef] [PubMed]
- WHO Organization. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 3 July 2022).
- Kumar, A.; Jaitak, V. Natural products as multidrug resistance modulators in cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef]
- Yuan, J.; Li, X.; Zhang, Y.; Zhang, G.; Cheng, W.; Wang, W.; Lei, Y.; Song, G. USP39 attenuates the antitumor activity of cisplatin on colon cancer cells dependent on p53. Cell Biol. Toxicol. 2021, 1–16. [Google Scholar] [CrossRef]
- Olatunde, A.; Nigam, M.; Singh, R.K.; Panwar, A.S.; Lasisi, A.; Alhumaydhi, F.A.; Jyoti Kumar, V.; Mishra, A.P.; Sharifi-Rad, J. Cancer and diabetes: The interlinking metabolic pathways and repurposing actions of antidiabetic drugs. Cancer Cell Int. 2021, 21, 499. [Google Scholar] [CrossRef]
- Su, Z.; Dong, S.; Zhao, S.-C.; Liu, K.; Tan, Y.; Jiang, X.; Assaraf, Y.G.; Qin, B.; Chen, Z.-S.; Zou, C. Novel nanomedicines to overcome cancer multidrug resistance. Drug Resist. Updat. 2021, 58, 100777. [Google Scholar] [CrossRef]
- Ahmad, M.H.; Rizvi, M.A.; Fatima, M.; Mondal, A.C. Pathophysiological implications of neuroinflammation mediated HPA axis dysregulation in the prognosis of cancer and depression. Mol. Cell. Endocrinol. 2021, 520, 111093. [Google Scholar] [CrossRef]
- Evans, S.O.; Jacobson, G.M.; Goodman, H.J.B.; Bird, S.; Jameson, M.B. Comparative safety and pharmacokinetic evaluation of three oral selenium compounds in cancer patients. Biol. Trace Elem. Res. 2019, 189, 395–404. [Google Scholar] [CrossRef]
- Hu, W.; Zhao, C.; Hu, H.; Yin, S. Food sources of selenium and its relationship with chronic diseases. Nutrients 2021, 13, 1739. [Google Scholar] [CrossRef]
- Gandin, V.; Khalkar, P.; Braude, J.; Fernandes, A.P. Organic selenium compounds as potential chemotherapeutic agents for improved cancer treatment. Free Radic. Biol. Med. 2018, 127, 80–97. [Google Scholar] [CrossRef]
- Rataan, A.O.; Geary, S.M.; Zakharia, Y.; Rustum, Y.M.; Salem, A.K. Potential role of selenium in the treatment of cancer and viral infections. Int. J. Mol. Sci. 2022, 23, 2215. [Google Scholar] [CrossRef]
- Radomska, D.; Czarnomysy, R.; Radomski, D.; Bielawski, K. Selenium compounds as novel potential anticancer agents. Int. J. Mol. Sci. 2021, 22, 1009. [Google Scholar] [CrossRef]
- Tan, H.W.; Mo, H.Y.; Lau, A.T.Y.; Xu, Y.M. Selenium species: Current status and potentials in cancer prevention and therapy. Int. J. Mol. Sci. 2018, 20, 75. [Google Scholar] [CrossRef]
- Stolwijk, J.M.; Garje, R.; Sieren, J.C.; Buettner, G.R.; Zakharia, Y. Understanding the redox biology of selenium in the search of targeted cancer therapies. Antioxidants 2020, 9, 420. [Google Scholar] [CrossRef]
- Kim, J.B.; Yang, E.Y.; Woo, J.; Kwon, H.; Lim, W.; Moon, B.I. Sodium selenite enhanced the anti-proliferative effect of MEK-ERK inhibitor in thyroid cancer cells. In Vivo 2020, 34, 185–190. [Google Scholar] [CrossRef]
- Domínguez-Álvarez, E.; Rácz, B.; Marć, M.A.; Nasim, M.J.; Szemerédi, N.; Viktorová, J.; Jacob, C.; Spengler, G. Selenium and tellurium in the development of novel small molecules and nanoparticles as cancer multidrug resistance reversal agents. Drug Resist. Updat. 2022, 63, 100844. [Google Scholar] [CrossRef]
- Szemerédi, N.; Dobiasová, S.; Salardón-Jiménez, N.; Kincses, A.; Nové, M.; Habibullah, G.; Sevilla-Hernández, C.; Benito-Lama, M.; Alonso-Martínez, F.-J.; Viktorová, J.; et al. Cyano- and ketone-containing selenoesters as multi-target compounds against resistant cancers. Cancers 2021, 13, 4563. [Google Scholar] [CrossRef]
- Valente, A.; Podolski-Renić, A.; Poetsch, I.; Filipović, N.; López, Ó.; Turel, I.; Heffeter, P. Metal- and metalloid-based compounds to target and reverse cancer multidrug resistance. Drug Resist. Updat. 2021, 58, 100778. [Google Scholar] [CrossRef]
- Lendvai, G.; Szekerczes, T.; Kontsek, E.; Selvam, A.; Szakos, A.; Schaff, Z.; Bjornstedt, M.; Kiss, A. The effect of methylselenocysteine and sodium selenite treatment on microRNA expression in liver cancer cell lines. Pathol. Oncol. Res. 2020, 26, 2669–2681. [Google Scholar] [CrossRef]
- Jablonska, E.; Li, Q.; Reszka, E.; Wieczorek, E.; Tarhonska, K.; Wang, T. Therapeutic potential of selenium and selenium compounds in cervical cancer. Cancer Control 2021, 28, 10732748211001808. [Google Scholar] [CrossRef] [PubMed]
- Augustyniak, M. Selenium and cancer or adenoma related to the large bowel. Postepy Hig. Med. Dosw. 2021, 75, 426–436. [Google Scholar] [CrossRef]
- Razaghi, A.; Poorebrahim, M.; Sarhan, D.; Björnstedt, M. Selenium stimulates the antitumour immunity: Insights to future research. Eur. J. Cancer 2021, 155, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Huang, Y.; Ge, Y.; Hong, X.; Lin, X.; Tang, K.; Wang, Q.; Yang, Y.; Sun, W.; Huang, Y.; et al. Selenite-induced ROS/AMPK/FoxO3a/GABARAPL-1 signaling pathway modulates autophagy that antagonize apoptosis in colorectal cancer cells. Discov. Oncol. 2021, 12, 35. [Google Scholar] [CrossRef]
- Doello, K.; Mesas, C.; Quinonero, F.; Perazzoli, G.; Cabeza, L.; Prados, J.; Melguizo, C.; Ortiz, R. The antitumor activity of sodium selenite alone and in combination with gemcitabine in pancreatic cancer: An in vitro and in vivo study. Cancers 2021, 13, 3169. [Google Scholar] [CrossRef]
- Soukupova, K.; Rudolf, E. Suppression of proliferation and activation of cell death by sodium selenite involves mitochondria and lysosomes in chemoresistant bladder cancer cells. J. Trace Elem. Med. Biol. 2019, 52, 58–67. [Google Scholar] [CrossRef]
- Larrouquère, L.; Berthier, S.; Chovelon, B.; Garrel, C.; Vacchina, V.; Paucot, H.; Boutonnat, J.; Faure, P.; Hazane-Puch, F. Preclinical evaluation of sodium selenite in mice: Toxicological and tumor regression studies after striatum implantation of human glioblastoma stem cells. Int. J. Mol. Sci. 2021, 22, 10646. [Google Scholar] [CrossRef]
- Powers, M.; Liu, L.; Deemer, D.; Chen, S.; Scholl, A.; Yoshinaga, M.; Liu, Z. Selenite inhibits Notch signaling in cells and mice. Int. J. Mol. Sci. 2021, 22, 2518. [Google Scholar] [CrossRef]
- Woo, J.; Kim, J.B.; Cho, T.; Yoo, E.H.; Moon, B.-I.; Kwon, H.; Lim, W. Selenium inhibits growth of trastuzumab-resistant human breast cancer cells via downregulation of Akt and beclin-1. PLoS ONE 2021, 16, e0257298. [Google Scholar] [CrossRef]
- Shimada, B.K.; Alfulaij, N.; Seale, L.A. The impact of selenium deficiency on cardiovascular function. Int. J. Mol. Sci. 2021, 22, 10713. [Google Scholar] [CrossRef]
- Bomer, N.; Grote Beverborg, N.; Hoes, M.F.; Streng, K.W.; Vermeer, M.; Dokter, M.M.; IJmker, J.; Anker, S.D.; Cleland, J.G.F.; Hillege, H.L.; et al. Selenium and outcome in heart failure. Eur. J. Heart Fail. 2020, 22, 1415–1423. [Google Scholar] [CrossRef]
- Shi, Y.; Yang, W.; Tang, X.; Yan, Q.; Cai, X.; Wu, F. Keshan disease: A potentially fatal endemic cardiomyopathy in remote mountains of China. Front. Pediatr. 2021, 9, 576916. [Google Scholar] [CrossRef]
- Yan, C.; Luo, R.; Li, F.; Liu, M.; Li, J.; Hua, W.; Li, X. The epidemiological status, environmental and genetic factors in the etiology of Keshan disease. Cardiovasc. Endocrinol. Metab. 2021, 10, 14–21. [Google Scholar] [CrossRef]
- Xia, J.; Xia, X.; Wang, W.; Xia, J.; Li, M. Protective effect of Se-methylselenocysteine on elaidic acid-induced inflammation in human arterial endothelial cells. J. Nutr. Sci. Vitaminol. 2020, 66, 577–582. [Google Scholar] [CrossRef]
- Cai, C.; Wu, Y.; Yang, L.; Xiang, Y.; Zhu, N.; Zhao, H.; Hu, W.; Lv, L.; Zeng, C. Sodium selenite attenuates balloon injury-induced and monocrotaline-induced vascular remodeling in rats. Front. Pharmacol. 2021, 12, 618493. [Google Scholar] [CrossRef]
- Ye, C.; Zheng, F.; Wu, N.; Zhu, G.-q.; Li, X.-z. Extracellular vesicles in vascular remodeling. Acta Pharmacol. Sin. 2022, 43, 2191–2201. [Google Scholar] [CrossRef]
- Xiao, J.; Li, N.; Xiao, S.; Wu, Y.; Liu, H. Comparison of selenium nanoparticles and sodium selenite on the alleviation of early atherosclerosis by inhibiting endothelial dysfunction and inflammation in apolipoprotein E-deficient mice. Int. J. Mol. Sci. 2021, 22, 11612. [Google Scholar] [CrossRef]
- Dong, H.L.; Yuan, N.; Sun, T.; Dun, A.S.; Hou, H.F. Effects of selenium supplement on atherogenesis of ApoE-knockout mice fed high fat diet. Zhonghua Xin Xue Guan Bing Za Zhi 2016, 44, 244–249. [Google Scholar]
- Krohn, R.M.; Lemaire, M.; Negro Silva, L.F.; Lemarié, C.; Bolt, A.; Mann, K.K.; Smits, J.E. High-selenium lentil diet protects against arsenic-induced atherosclerosis in a mouse model. J. Nutr. Biochem. 2016, 27, 9–15. [Google Scholar] [CrossRef]
- Wang, X.; Sui, H.; Su, Y.; Zhao, S. Protective effects of sodium selenite on insulin secretion and diabetic retinopathy in rats with type 1 diabetes mellitus. Pak. J. Pharm. Sci. 2021, 34, 1729–1735. [Google Scholar]
- Schomburg, L. Selenium deficiency due to diet, pregnancy, severe illness, or COVID-19—A preventable trigger for autoimmune disease. Int. J. Mol. Sci. 2021, 22, 8532. [Google Scholar] [CrossRef]
- Ilonen, J.; Lempainen, J.; Veijola, R. The heterogeneous pathogenesis of type 1 diabetes mellitus. Nat. Rev. Endocrinol. 2019, 15, 635–650. [Google Scholar] [CrossRef]
- Cecilia, O.M.; José Alberto, C.G.; José, N.P.; Ernesto Germán, C.M.; Ana Karen, L.C.; Luis Miguel, R.P.; Ricardo Raúl, R.R.; Adolfo Daniel, R.C. Oxidative stress as the main target in diabetic retinopathy pathophysiology. J. Diabetes Res. 2019, 2019, 8562408. [Google Scholar] [CrossRef]
- Muller, T.E.; Nunes, M.E.; Menezes, C.C.; Marins, A.T.; Leitemperger, J.; Gressler, A.C.L.; Carvalho, F.B.; de Freitas, C.M.; Quadros, V.A.; Fachinetto, R.; et al. Sodium selenite prevents paraquat-induced neurotoxicity in zebrafish. Mol. Neurobiol. 2018, 55, 1928–1941. [Google Scholar] [CrossRef]
- Colle, D.; Santos, D.B.; de Souza, V.; Lopes, M.W.; Leal, R.B.; de Souza Brocardo, P.; Farina, M. Sodium selenite protects from 3-nitropropionic acid-induced oxidative stress in cultured primary cortical neurons. Mol. Biol. Rep. 2019, 46, 751–762. [Google Scholar] [CrossRef]
- Vicente-Zurdo, D.; Romero-Sanchez, I.; Rosales-Conrado, N.; Leon-Gonzalez, M.E.; Madrid, Y. Ability of selenium species to inhibit metal-induced Aβ aggregation involved in the development of Alzheimer’s disease. Anal. Bioanal. Chem. 2020, 412, 6485–6497. [Google Scholar] [CrossRef]
- Juszczyk, G.; Mikulska, J.; Kasperek, K.; Pietrzak, D.; Mrozek, W.; Herbet, M. Chronic stress and oxidative stress as common factors of the pathogenesis of depression and Alzheimer’s disease: The role of antioxidants in prevention and treatment. Antioxidants 2021, 10, 1439. [Google Scholar] [CrossRef]
- Solovyev, N. Selenoprotein P and its potential role in Alzheimer’s disease. Hormones 2020, 19, 73–79. [Google Scholar] [CrossRef]
- Sharma, G.; Shin, E.J.; Sharma, N.; Nah, S.Y.; Mai, H.N.; Nguyen, B.T.; Jeong, J.H.; Lei, X.G.; Kim, H.C. Glutathione peroxidase-1 and neuromodulation: Novel potentials of an old enzyme. Food Chem. Toxicol. 2021, 148, 111945. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Song, G.L. Roles of selenoproteins in brain function and the potential mechanism of selenium in Alzheimer’s disease. Front. Neurosci. 2021, 15, 646518. [Google Scholar] [CrossRef] [PubMed]
- Ying, H.; Zhang, Y. Systems biology of selenium and complex disease. Biol. Trace Elem. Res. 2019, 192, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Reichert, C.O.; de Freitas, F.A.; Sampaio-Silva, J.; Rokita-Rosa, L.; Barros, P.d.L.; Levy, D.; Bydlowski, S.P. Ferroptosis mechanisms involved in neurodegenerative diseases. Int. J. Mol. Sci. 2020, 21, 8765. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xie, Z.; Pei, T.; Zeng, Y.; Xiong, Q.; Wei, H.; Wang, Y.; Cheng, W. Salidroside attenuates neuronal ferroptosis by activating the Nrf2/HO1 signaling pathway in Aβ1-42-induced Alzheimer’s disease mice and glutamate-injured HT22 cells. Chin. Med. 2022, 17, 82. [Google Scholar] [CrossRef]
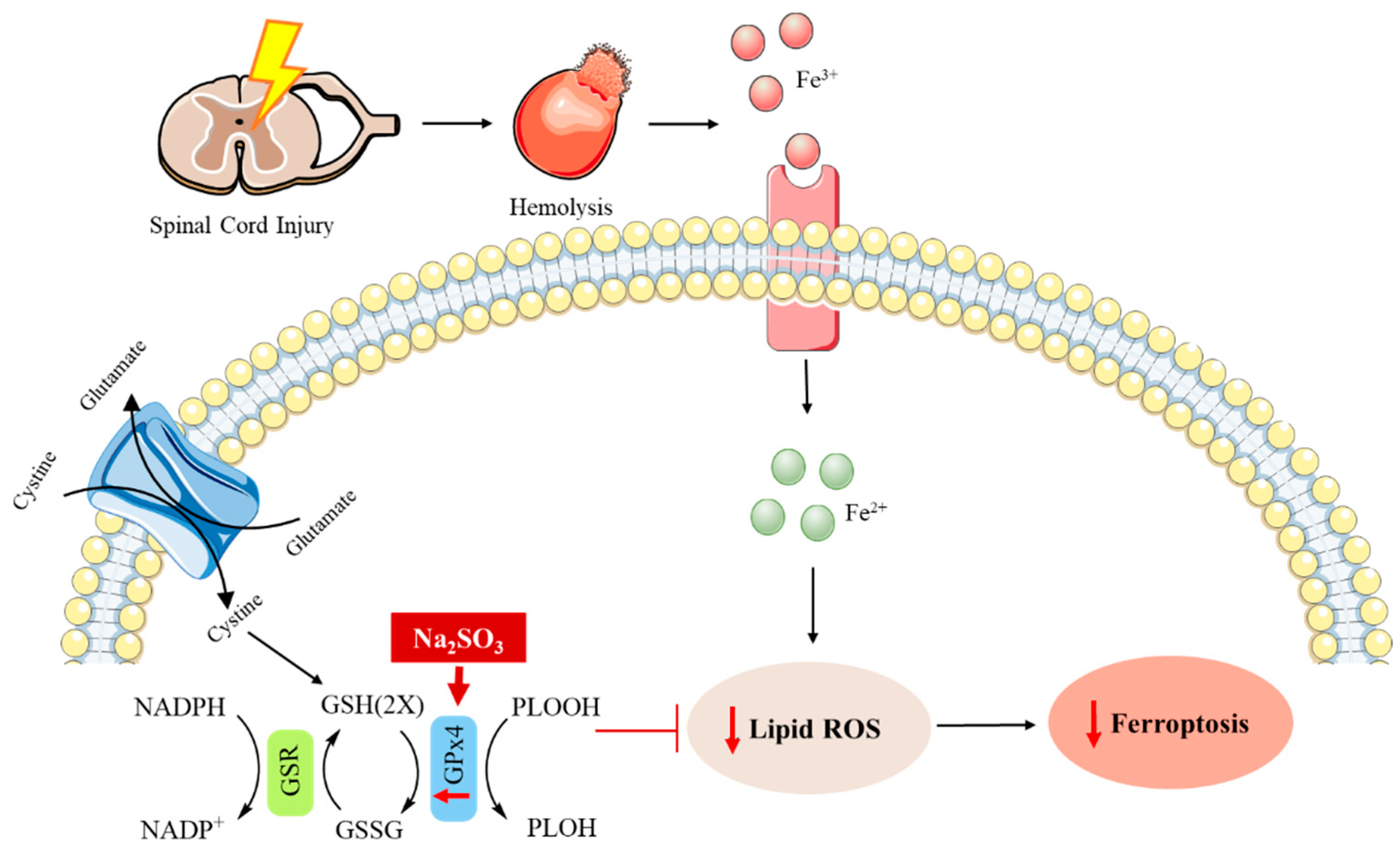
- Li, F.; Wang, H.; Chen, H.; Guo, J.; Dang, X.; Ru, Y.; Wang, H. Mechanism of ferroptosis and its role in spinal cord injury. Front. Neurol. 2022, 13, 926780. [Google Scholar] [CrossRef]
- Chen, Y.X.; Zuliyaer, T.; Liu, B.; Guo, S.; Yang, D.G.; Gao, F.; Yu, Y.; Yang, M.L.; Du, L.J.; Li, J.J. Sodium selenite promotes neurological function recovery after spinal cord injury by inhibiting ferroptosis. Neural Regen. Res. 2022, 17, 2702–2709. [Google Scholar] [CrossRef]
- Dang, X.; Huan, X.; Du, X.; Chen, X.; Bi, M.; Yan, C.; Jiao, Q.; Jiang, H. Correlation of ferroptosis and other types of cell death in neurodegenerative diseases. Neurosci. Bull. 2022, 38, 938–952. [Google Scholar] [CrossRef]
- Wu, H.; Luan, Y.; Wang, H.; Zhang, P.; Liu, S.; Wang, P.; Cao, Y.; Sun, H.; Wu, L. Selenium inhibits ferroptosis and ameliorates autistic-like behaviors of BTBR mice by regulating the Nrf2/GPX4 pathway. Brain Res. Bull. 2022, 183, 38–48. [Google Scholar] [CrossRef]
- Onukwufor, J.O.; Dirksen, R.T.; Wojtovich, A.P. Iron dysregulation in mitochondrial dysfunction and Alzheimer’s disease. Antioxidants 2022, 11, 692. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, S.; Li, J.; Li, Z.; Quan, J.; Liu, X.; Tang, Y.; Liu, B. The latest view on the mechanism of ferroptosis and its research progress in spinal cord injury. Oxid. Med. Cell. Longev. 2020, 2020, 6375938. [Google Scholar] [CrossRef]
- Wu, X.; Li, R.; Hong, Q.; Chi, X. Development and validation of a novel diagnostic model for childhood autism spectrum disorder based on ferroptosis-related genes. Front. Psychiatry 2022, 13, 886055. [Google Scholar] [CrossRef]
- Lord, C.; Brugha, T.S.; Charman, T.; Cusack, J.; Dumas, G.; Frazier, T.; Jones, E.J.H.; Jones, R.M.; Pickles, A.; State, M.W.; et al. Autism spectrum disorder. Nat. Rev. Dis. Primers 2020, 6, 5. [Google Scholar] [CrossRef]
- Tuo, Q.Z.; Masaldan, S.; Southon, A.; Mawal, C.; Ayton, S.; Bush, A.I.; Lei, P.; Belaidi, A.A. Characterization of selenium compounds for anti-ferroptotic activity in neuronal cells and after cerebral ischemia-reperfusion injury. Neurotherapeutics 2021, 18, 2682–2691. [Google Scholar] [CrossRef]
- Alim, I.; Caulfield, J.T.; Chen, Y.; Swarup, V.; Geschwind, D.H.; Ivanova, E.; Seravalli, J.; Ai, Y.; Sansing, L.H.; Ste Marie, E.J.; et al. Selenium drives a transcriptional adaptive program to block ferroptosis and treat stroke. Cell 2019, 177, 1262–1279.e1225. [Google Scholar] [CrossRef] [PubMed]
- Capuz, A.; Karnoub, M.A.; Osien, S.; Rose, M.; Mériaux, C.; Fournier, I.; Devos, D.; Vanden Abeele, F.; Rodet, F.; Cizkova, D.; et al. The antibody dependant neurite outgrowth modulation response involvement in spinal cord injury. Front. Immunol. 2022, 13, 882830. [Google Scholar] [CrossRef]
- Zhang, L.-J.; Chen, Y.; Wang, L.-X.; Zhuang, X.-Q.; Xia, H.-C. Identification of potential oxidative stress biomarkers for spinal cord injury in erythrocytes using mass spectrometry. Neural Regen. Res. 2021, 16, 1294–1301. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, C.-B.; Yang, D.-G.; Qin, C.; Dong, X.-C.; Li, D.-P.; Zhang, C.; Guo, Y.; Du, L.-J.; Gao, F.; et al. Dynamic changes in intramedullary pressure 72 hours after spinal cord injury. Neural Regen. Res. 2019, 14, 886–895. [Google Scholar]
- McCreedy, D.A.; Abram, C.L.; Hu, Y.; Min, S.W.; Platt, M.E.; Kirchhoff, M.A.; Reid, S.K.; Jalufka, F.L.; Lowell, C.A. Spleen tyrosine kinase facilitates neutrophil activation and worsens long-term neurologic deficits after spinal cord injury. J. Neuroinflamm. 2021, 18, 302. [Google Scholar] [CrossRef]
- Ni, Y.; Gu, J.; Wu, J.; Xu, L.; Rui, Y. MGMT-mediated neuron apoptosis in injured rat spinal cord. Tissue Cell 2020, 62, 101311. [Google Scholar] [CrossRef]
- Shi, Z.; Yuan, S.; Shi, L.; Li, J.; Ning, G.; Kong, X.; Feng, S. Programmed cell death in spinal cord injury pathogenesis and therapy. Cell Prolif. 2021, 54, e12992. [Google Scholar] [CrossRef]
- Yao, X.; Zhang, Y.; Hao, J.; Duan, H.-Q.; Zhao, C.-X.; Sun, C.; Li, B.; Fan, B.-Y.; Wang, X.; Li, W.-X.; et al. Deferoxamine promotes recovery of traumatic spinal cord injury by inhibiting ferroptosis. Neural Regen. Res. 2019, 14, 532–541. [Google Scholar]
- Zhou, H.; Yin, C.; Zhang, Z.; Tang, H.; Shen, W.; Zha, X.; Gao, M.; Sun, J.; Xu, X.; Chen, Q. Proanthocyanidin promotes functional recovery of spinal cord injury via inhibiting ferroptosis. J. Chem. Neuroanat. 2020, 107, 101807. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Xue, X.; Xian, J.; Yuan, L.; Wang, L.; Zou, Y.; Zhong, J.; Jiang, Z.; Shi, J.; Chen, T.; et al. Ferrostatin-1 alleviates white matter injury via decreasing ferroptosis following spinal cord injury. Mol. Neurobiol. 2022, 59, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, C.; Zhao, C.; Hao, J.; Zhang, Y.; Fan, B.; Li, B.; Duan, H.; Liu, C.; Kong, X.; et al. Ferroptosis inhibitor SRS 16-86 attenuates ferroptosis and promotes functional recovery in contusion spinal cord injury. Brain Res. 2019, 1706, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.H.; Tian, H.; Mao, L.; Li, D.Y.; Lin, J.Q.; Hu, H.S.; Huang, S.C.; Zhang, C.J.; Mei, X.F. Zinc attenuates ferroptosis and promotes functional recovery in contusion spinal cord injury by activating Nrf2/GPX4 defense pathway. CNS Neurosci. Ther. 2021, 27, 1023–1040. [Google Scholar] [CrossRef]
- Jia, Z.-Q.; Li, S.-Q.; Qiao, W.-Q.; Xu, W.-Z.; Xing, J.-W.; Liu, J.-T.; Song, H.; Gao, Z.-Y.; Xing, B.-W.; He, X.-J. Ebselen protects mitochondrial function and oxidative stress while inhibiting the mitochondrial apoptosis pathway after acute spinal cord injury. Neurosci. Lett. 2018, 678, 110–117. [Google Scholar] [CrossRef]
- Luo, W.; Wang, Y.; Lin, F.; Liu, Y.; Gu, R.; Liu, W.; Xiao, C. Selenium-doped carbon quantum dots efficiently ameliorate secondary spinal cord injury via scavenging reactive oxygen species. Int. J. Nanomed. 2020, 15, 10113–10125. [Google Scholar] [CrossRef]
- Javdani, M.; Ghorbani, R.; Hashemnia, M. Histopathological evaluation of spinal cord with experimental traumatic injury following implantation of a controlled released drug delivery system of chitosan hydrogel loaded with selenium nanoparticle. Biol. Trace Elem. Res. 2021, 199, 2677–2686. [Google Scholar] [CrossRef]
- Rao, S.; Lin, Y.; Lin, R.; Liu, J.; Wang, H.; Hu, W.; Chen, B.; Chen, T. Traditional Chinese medicine active ingredients-based selenium nanoparticles regulate antioxidant selenoproteins for spinal cord injury treatment. J. Nanobiotechnol. 2022, 20, 278. [Google Scholar] [CrossRef]
- Wang, W.; Huang, X.; Zhang, Y.; Deng, G.; Liu, X.; Fan, C.; Xi, Y.; Yu, J.; Ye, X. Se@SiO2 nanocomposites suppress microglia-mediated reactive oxygen species during spinal cord injury in rats. RSC Adv. 2018, 8, 16126–16138. [Google Scholar] [CrossRef]
- Garcia, E.; Hernández-Ayvar, F.; Rodríguez-Barrera, R.; Flores-Romero, A.; Borlongan, C.; Ibarra, A. Supplementation with vitamin E, zinc, selenium, and copper re-establishes T-cell function and improves motor recovery in a rat model of spinal cord injury. Cell Transplant. 2022, 31, 9636897221109884. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhi, X.; Sun, G.; Guo, W.; Huang, Y.; Sun, W.; Tian, X.; Zhao, F.; Hu, K. Sodium selenite suppresses hepatitis B virus transcription and replication in human hepatoma cell lines. J. Med. Virol. 2016, 88, 653–663. [Google Scholar] [CrossRef]
- Bermano, G.; Méplan, C.; Mercer, D.K.; Hesketh, J.E. Selenium and viral infection: Are there lessons for COVID-19? Br. J. Nutr. 2021, 125, 618–627. [Google Scholar] [CrossRef]
- Guillin, O.M.; Vindry, C.; Ohlmann, T.; Chavatte, L. Selenium, selenoproteins and viral infection. Nutrients 2019, 11, 2101. [Google Scholar] [CrossRef]
- Martinez, S.S.; Huang, Y.; Acuna, L.; Laverde, E.; Trujillo, D.; Barbieri, M.A.; Tamargo, J.; Campa, A.; Baum, M.K. Role of selenium in viral infections with a major focus on SARS-CoV-2. Int. J. Mol. Sci. 2021, 23, 280. [Google Scholar] [CrossRef]
- Lima, L.W.; Nardi, S.; Santoro, V.; Schiavon, M. The relevance of plant-derived Se compounds to human health in the SARS-CoV-2 (COVID-19) pandemic era. Antioxidants 2021, 10, 1031. [Google Scholar] [CrossRef]
- Zhu, Y.-H.; Wang, X.-F.; Yang, G.; Wei, J.; Tan, W.-H.; Wang, L.-X.; Guo, X.; Lammi, M.J.; Xu, J.-H. Efficacy of long-term selenium supplementation in the treatment of chronic Keshan disease with congestive heart failure. Curr. Med. Sci. 2019, 39, 237–242. [Google Scholar] [CrossRef]
- Thompson, R.; Wood, J.G.; Tempia, S.; Muscatello, D.J. Global variation in early epidemic growth rates and reproduction number of seasonal influenza. Int. J. Infect. Dis. 2022, 122, 382–388. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for the Clinical Management of Severe Illness from Influenza Virus Infections; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Lyons, D.M.; Lauring, A.S. Mutation and epistasis in influenza virus evolution. Viruses 2018, 10, 407. [Google Scholar] [CrossRef]
- Yin, H.; Jiang, N.; Shi, W.; Chi, X.; Liu, S.; Chen, J.-L.; Wang, S. Development and effects of influenza antiviral drugs. Molecules 2021, 26, 810. [Google Scholar] [CrossRef]
- Honce, R.; Schultz-Cherry, S. Impact of obesity on influenza a virus pathogenesis, immune response, and evolution. Front. Immunol. 2019, 10, 1071. [Google Scholar] [CrossRef]
- Gong, G.; Li, Y.; He, K.; Yang, Q.; Guo, M.; Xu, T.; Wang, C.; Zhao, M.; Chen, Y.; Du, M.; et al. The inhibition of H1N1 influenza induced apoptosis by sodium selenite through ROS-mediated signaling pathways. RSC Adv. 2020, 10, 8002–8007. [Google Scholar] [CrossRef]
- Bai, Y.; Jones, J.C.; Wong, S.-S.; Zanin, M. Antivirals targeting the surface glycoproteins of influenza virus: Mechanisms of action and resistance. Viruses 2021, 13, 624. [Google Scholar] [CrossRef]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef]
- Majeed, M.; Nagabhushanam, K.; Prakasan, P.; Mundkur, L. Can selenium reduce the susceptibility and severity of SARS-CoV-2?—A comprehensive review. Int. J. Mol. Sci. 2022, 23, 4809. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Saad, R.; Taylor, E.W.; Rayman, M.P. Selenium and selenoproteins in viral infection with potential relevance to COVID-19. Redox Biol. 2020, 37, 101715. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Lipinski, B. Selenium supplementation in the prevention of coronavirus infections (COVID-19). Med. Hypotheses 2020, 143, 109878. [Google Scholar] [CrossRef] [PubMed]
- Ali, W.; Benedetti, R.; Handzlik, J.; Zwergel, C.; Battistelli, C. The innovative potential of selenium-containing agents for fighting cancer and viral infections. Drug Discov. Today 2021, 26, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Varlamova, E.G.; Turovsky, E.A. The main cytotoxic effects of methylseleninic acid on various cancer cells. Int. J. Mol. Sci. 2021, 22, 6614. [Google Scholar] [CrossRef]
- Park, J.-M.; Kim, D.-H.; Na, H.-K.; Surh, Y.-J. Methylseleninic acid induces NAD(P)H:quinone oxidoreductase-1 expression through activation of NF-E2-related factor 2 in Chang liver cells. Oncotarget 2016, 9, 3014–3028. [Google Scholar] [CrossRef]
- Nair, D.; Rådestad, E.; Khalkar, P.; Diaz-Argelich, N.; Schröder, A.; Klynning, C.; Ungerstedt, J.; Uhlin, M.; Fernandes, A.P. Methylseleninic acid sensitizes ovarian cancer cells to T-cell mediated killing by decreasing PDL1 and VEGF levels. Front. Oncol. 2018, 8, 407. [Google Scholar] [CrossRef]
- Hu, W.; Ma, Y.; Zhao, C.; Yin, S.; Hu, H. Methylseleninic acid overcomes programmed death-ligand 1-mediated resistance of prostate cancer and lung cancer. Mol. Carcinog. 2021, 60, 746–757. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Hu, G.; Hu, C.; Gao, Y.; Huo, M.; Zhu, H.; Liu, M.; Xu, N. EGFR-IL-6 signaling axis mediated the inhibitory effect of methylseleninic acid on esophageal squamous cell carcinoma. Front. Pharmacol. 2021, 12, 719785. [Google Scholar] [CrossRef]
- Xin, M.; Gao, Q.; Xiang, X.; Xu, J.; Jiao, Y.; Li, X.; Zhang, X.; Jia, X. Autophagy inhibition enhances the anti-tumor activity of methylseleninic acid in cisplatin-resistance human lung adenocarcinoma cells. Front. Pharmacol. 2022, 13, 890974. [Google Scholar] [CrossRef]
- Lafin, J.T.; Sarsour, E.H.; Kalen, A.L.; Wagner, B.A.; Buettner, G.R.; Goswami, P.C. Methylseleninic acid induces lipid peroxidation and radiation sensitivity in head and neck cancer cells. Int. J. Mol. Sci. 2019, 20, 225. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Sun, Y.; Yang, S.; Liang, X. Inhibitory effect of selenium on esophagus cancer cells and the related mechanism. J. Nutr. Sci. Vitaminol. 2020, 66, 456–461. [Google Scholar] [CrossRef]
- Zhang, W.; Hu, C.; Wang, X.; Bai, S.; Cao, S.; Kobelski, M.; Lambert, J.R.; Gu, J.; Zhan, Y. Role of GDF15 in methylseleninic acid-mediated inhibition of cell proliferation and induction of apoptosis in prostate cancer cells. PLoS ONE 2019, 14, e0222812. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, Y.; Han, D.; Lu, J.; Yin, S.; Hu, H.; Zhao, C. Combination of palmitic acid and methylseleninic acid induces mitochondria-dependent apoptosis via attenuation of the IRE1α arm and enhancement of CHOP in hepatoma. ACS Omega 2021, 6, 15708–15715. [Google Scholar] [CrossRef]
- Sundaram, S.; Yan, L. Dietary supplementation with methylseleninic acid inhibits mammary tumorigenesis and metastasis in male MMTV-PyMT mice. Biol. Trace Elem. Res. 2018, 184, 186–195. [Google Scholar] [CrossRef]
- Cai, Z.; Dong, L.; Song, C.; Zhang, Y.; Zhu, C.; Zhang, Y.; Ling, Q.; Hoffmann, P.R.; Li, J.; Huang, Z.; et al. Methylseleninic acid provided at nutritional selenium levels inhibits angiogenesis by down-regulating integrin β3 signaling. Sci. Rep. 2017, 7, 9445. [Google Scholar] [CrossRef]
- Goltyaev, M.V.; Varlamova, E.G.; Novoselov, S.V.; Fesenko, E.E. Activation of signal pathways of apoptosis under conditions of prolonged ER-stress caused by exposure of mouse testicular teratoma cells to selenium-containing compounds. Dokl. Biochem. Biophys. 2020, 490, 9–11. [Google Scholar] [CrossRef]
- Durgan, J.; Florey, O. Cancer cell cannibalism: Multiple triggers emerge for entosis. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 831–841. [Google Scholar] [CrossRef]
- Khalkar, P.; Díaz-Argelich, N.; Antonio Palop, J.; Sanmartín, C.; Fernandes, A.P. Novel methylselenoesters induce programed cell death via entosis in pancreatic cancer cells. Int. J. Mol. Sci. 2018, 19, 2849. [Google Scholar] [CrossRef]
- Mlynarczuk-Bialy, I.; Dziuba, I.; Sarnecka, A.; Platos, E.; Kowalczyk, M.; Pels, K.K.; Wilczynski, G.M.; Wojcik, C.; Bialy, L.P. Entosis: From cell biology to clinical cancer pathology. Cancers 2020, 12, 2481. [Google Scholar] [CrossRef]
- Lobb, R.J.; Jacobson, G.M.; Cursons, R.T.; Jameson, M.B. The interaction of selenium with chemotherapy and radiation on normal and malignant human mononuclear blood cells. Int. J. Mol. Sci. 2018, 19, 3167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, X.; Wu, L.; Liu, D.; Pei, T.; Huang, W. Existence of chronic endometritis and its influence on pregnancy outcomes in infertile women with minimal/mild endometriosis. Int. J. Gynaecol. Obstet. 2022. [Google Scholar] [CrossRef] [PubMed]
- Puente, E.; Alonso, L.; Laganà, A.S.; Ghezzi, F.; Casarin, J.; Carugno, J. Chronic endometritis: Old problem, novel insights and future challenges. Int. J. Fertil. Steril. 2020, 13, 250–256. [Google Scholar] [PubMed]
- Liu, Y.; Qiu, C.; Li, W.; Mu, W.; Li, C.; Guo, M. Selenium plays a protective role in Staphylococcus aureus-induced endometritis in the uterine tissue of rats. Biol. Trace Elem. Res. 2016, 173, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M. Selenium in the prevention of SARS-CoV-2 and other viruses. Biol. Trace Elem. Res. 2022. [Google Scholar] [CrossRef]
- Gopalakrishna, R.; Gundimeda, U.; Zhou, S.; Zung, K.; Forell, K.; Holmgren, A. Imbalance in protein thiol redox regulation and cancer-preventive efficacy of selenium. React. Oxyg. Species 2016, 2, 272–289. [Google Scholar] [CrossRef]
- Gopalakrishna, R.; Gundimeda, U.; Zhou, S.; Bui, H.; Holmgren, A. Redox regulation of protein kinase C by selenometabolites and selenoprotein thioredoxin reductase limits cancer prevention by selenium. Free Radic. Biol. Med. 2018, 127, 55–61. [Google Scholar] [CrossRef]
- World Health Organization. Global Tuberculosis Report 2021. Available online: https://www.who.int/publications/i/item/9789240037021 (accessed on 3 July 2022).
- Park, S.; Jung, J.; Kim, J.; Han, S.B.; Ryoo, S. Investigation of clofazimine resistance and genetic mutations in drug-resistant Mycobacterium tuberculosis isolates. J. Clin. Med. 2022, 11, 1927. [Google Scholar] [CrossRef]
- Shah, I.; Poojari, V.; Meshram, H. Multi-drug resistant and extensively-drug resistant tuberculosis. Indian J. Pediatr. 2020, 87, 833–839. [Google Scholar] [CrossRef]
- Li, H.; Yuan, J.; Duan, S.; Pang, Y. Resistance and tolerance of Mycobacterium tuberculosis to antimicrobial agents—How M. tuberculosis can escape antibiotics. WIREs Mech. Dis. 2022, e1573. [Google Scholar] [CrossRef]
- Perveen, S.; Kumari, D.; Singh, K.; Sharma, R. Tuberculosis drug discovery: Progression and future interventions in the wake of emerging resistance. Eur. J. Med. Chem. 2022, 229, 114066. [Google Scholar] [CrossRef]
- Chen, W.; Liu, Z.; Zheng, Y.; Wei, B.; Shi, J.; Shao, B.; Wang, D. Selenium donor restricts the intracellular growth of Mycobacterium tuberculosis through the induction of c-Jun-mediated both canonical autophagy and LC3-associated phagocytosis of alveolar macrophages. Microb. Pathog. 2021, 161, 105269. [Google Scholar] [CrossRef]
- Hussain, M.I.; Ahmed, W.; Nasir, M.; Mushtaq, M.H.; Sheikh, A.A.; Shaheen, A.Y.; Ali, S.; Iqbal, S. Immune modulatory and anti-oxidative effect of selenium against pulmonary tuberculosis. Pak. J. Pharm. Sci. 2019, 32, 779–784. [Google Scholar]
- Ribeiro, R.C.B.; de Marins, D.B.; Di Leo, I.; da Silva Gomes, L.; de Moraes, M.G.; Abbadi, B.L.; Villela, A.D.; da Silva, W.F.; da Silva, L.C.R.P.; Machado, P.; et al. Anti-tubercular profile of new selenium-menadione conjugates against Mycobacterium tuberculosis H37Rv (ATCC 27294) strain and multidrug-resistant clinical isolates. Eur. J. Med. Chem. 2021, 209, 112859. [Google Scholar] [CrossRef]
- Estevez, H.; Palacios, A.; Gil, D.; Anguita, J.; Vallet-Regi, M.; González, B.; Prados-Rosales, R.; Luque-Garcia, J.L. Antimycobacterial effect of selenium nanoparticles on Mycobacterium tuberculosis. Front. Microbiol. 2020, 11, 800. [Google Scholar] [CrossRef]
- Muzembo, B.A.; Mbendi, N.C.; Ngatu, N.R.; Suzuki, T.; Wada, K.; Ikeda, S. Serum selenium levels in tuberculosis patients: A systematic review and meta-analysis. J. Trace Elem. Med. Biol. 2018, 50, 257–262. [Google Scholar] [CrossRef]
- Du, X.; Shi, Q.; Zhao, Y.; Xie, Y.; Li, X.; Liu, Q.; Iqbal, J.; Zhang, H.; Liu, X.; Shen, L. Se-methylselenocysteine (SMC) improves cognitive deficits by attenuating synaptic and metabolic abnormalities in Alzheimer’s mice model: A proteomic study. ACS Chem. Neurosci. 2021, 12, 1112–1132. [Google Scholar] [CrossRef]
- Rao, S.; Gou, Y.; Yu, T.; Cong, X.; Gui, J.; Zhu, Z.; Zhang, W.; Liao, Y.; Ye, J.; Cheng, S.; et al. Effects of selenate on Se, flavonoid, and glucosinolate in broccoli florets by combined transcriptome and metabolome analyses. Food Res. Int. 2021, 146, 110463. [Google Scholar] [CrossRef]
- Ding, J.; Qi, C.; Li, J.; Huang, C.; Zhang, J.; Zhang, Y.; Li, Y.; Fan, B. Se-methylselenocysteine alleviates liver injury in diethylnitrosamine (DEN)-induced hepatocellular carcinoma rat model by reducing liver enzymes, inhibiting angiogenesis, and suppressing nitric oxide (NO)/nitric oxide synthase (NOS) signaling pathway. Med. Sci. Monit. 2021, 27, e929255. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Zhong, Y.; Qin, L.; Li, J. Sex-dimorphic distribution and anti-oxidative effects of selenomethionine and Se-methylselenocysteine supplementation. J. Food Sci. 2021, 86, 5424–5438. [Google Scholar] [CrossRef]
- Selvam, A.K.; Björnstedt, M. A novel assay method to determine the β-elimination of Se-methylselenocysteine to monomethylselenol by kynurenine aminotransferase 1. Antioxidants 2020, 9, 139. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, Q.; Zheng, L.; Wang, B.; Qu, X.; Ni, J.; Zhang, Y.; Du, X. Se-methylselenocysteine ameliorates neuropathology and cognitive deficits by attenuating oxidative stress and metal dyshomeostasis in Alzheimer model mice. Mol. Nutr. Food Res. 2018, 62, e1800107. [Google Scholar] [CrossRef]
- Ma, J.; Huang, J.; Sun, J.; Zhou, Y.; Ji, X.; Guo, D.; Liu, C.; Li, J.; Zhang, J.; Song, H. L-Se-methylselenocysteine sensitizes lung carcinoma to chemotherapy. Cell Prolif. 2021, 54, e13038. [Google Scholar] [CrossRef]
- Selvam, A.K.; Björnstedt, M.; Misra, S. Therapeutic potential of selenium compounds in the treatment of cancer. In Selenium; Michalke, B., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 251–269. [Google Scholar]
- Adimulam, T.; Arumugam, T.; Foolchand, A.; Ghazi, T.; Chuturgoon, A.A. The effect of organoselenium compounds on histone deacetylase inhibition and their potential for cancer therapy. Int. J. Mol. Sci. 2021, 22, 12952. [Google Scholar] [CrossRef]
- Evans, S.O.; Khairuddin, P.F.; Jameson, M.B. Optimising selenium for modulation of cancer treatments. Anticancer Res. 2017, 37, 6497–6509. [Google Scholar] [PubMed]
- Gonçalves Correa, N.D.; Silva, F.D.; Vieira, D.P.; Soares, C.R.J.; de Queiroz, A.A.A. In vitro cytotoxic data on Se-methylselenocysteine conjugated to dendritic poly(glycerol) against human squamous carcinoma cells. J. Biomater. Sci. Polym. Ed. 2022, 33, 651–667. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Ohman Strickland, P.A.; Kang, H.-G.; Zarbl, H. Uncoupling genotoxic stress responses from circadian control increases susceptibility to mammary carcinogenesis. Oncotarget 2017, 8, 32752–32768. [Google Scholar] [CrossRef] [PubMed]
- Evans, S.O.; Jacobson, G.M.; Goodman, H.J.B.; Bird, S.; Jameson, M.B. Comparison of three oral selenium compounds in cancer patients: Evaluation of differential pharmacodynamic effects in normal and malignant cells. J. Trace Elem. Med. Biol. 2020, 58, 126446. [Google Scholar] [CrossRef]
- Rustum, Y.M.; Chintala, S.; Durrani, F.A.; Bhattacharya, A. Non-coding micro RNAs and hypoxia-inducible factors are selenium targets for development of a mechanism-based combination strategy in clear-cell renal cell carcinoma—bench-to-bedside therapy. Int. J. Mol. Sci. 2018, 19, 3378. [Google Scholar] [CrossRef]
- Zhou, X.; Li, M.; Cheng, Q.; Shao, Y.; Wang, W.; Du, Q.; Liu, J.; Yang, Y. Methylselenocysteine potentiates etoposide-induced cytotoxicity by enhancing gap junction activity. Biol. Pharm. Bull. 2022, 45, 467–476. [Google Scholar] [CrossRef]
- Farr, A.; Effendy, I.; Frey Tirri, B.; Hof, H.; Mayser, P.; Petricevic, L.; Ruhnke, M.; Schaller, M.; Schaefer, A.P.A.; Sustr, V.; et al. Guideline: Vulvovaginal candidosis (AWMF 015/072, level S2k). Mycoses 2021, 64, 583–602. [Google Scholar] [CrossRef]
- Pereira, L.C.; Correia, A.F.; da Silva, Z.D.L.; de Resende, C.N.; Brandão, F.; Almeida, R.M.; de Medeiros Nóbrega, Y.K. Vulvovaginal candidiasis and current perspectives: New risk factors and laboratory diagnosis by using MALDI TOF for identifying species in primary infection and recurrence. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1681–1693. [Google Scholar] [CrossRef]
- Mohankumar, B.; Shandil, R.K.; Narayanan, S.; Krishnan, U.M. Vaginosis: Advances in new therapeutic development and microbiome restoration. Microb. Pathog. 2022, 168, 105606. [Google Scholar] [CrossRef]
- Jafarzadeh, L.; Ranjbar, M.; Nazari, T.; Naeimi Eshkaleti, M.; Aghaei Gharehbolagh, S.; Sobel, J.D.; Mahmoudi, S. Vulvovaginal candidiasis: An overview of mycological, clinical, and immunological aspects. J. Obstet. Gynaecol. Res. 2022, 48, 1546–1560. [Google Scholar] [CrossRef]
- Sobel, J.D.; Sobel, R. Current treatment options for vulvovaginal candidiasis caused by azole-resistant Candida species. Expert Opin. Pharmacother. 2018, 19, 971–977. [Google Scholar] [CrossRef]
- Denning, D.W.; Kneale, M.; Sobel, J.D.; Rautemaa-Richardson, R. Global burden of recurrent vulvovaginal candidiasis: A systematic review. Lancet Infect. Dis. 2018, 18, e339–e347. [Google Scholar] [CrossRef]
- Yang, X.; Wang, M.; Kang, X.; Mo, F.; Si, P.; Ma, J.; Zhang, P.; Zheng, S.; Li, J.; Wang, Y.; et al. L-Se-methylselenocysteine loaded mucoadhesive thermogel for effective treatment of vulvar candidiasis. Int. J. Pharm. 2022, 622, 121851. [Google Scholar] [CrossRef]
- Thapa, K.; Khan, H.; Kanojia, N.; Singh, T.G.; Kaur, A.; Kaur, G. Therapeutic insights on ferroptosis in Parkinson’s disease. Eur. J. Pharmacol. 2022, 930, 175133. [Google Scholar] [CrossRef] [PubMed]
- Peeples, E.S.; Genaro-Mattos, T.C. Ferroptosis: A promising therapeutic target for neonatal hypoxic-ischemic brain injury. Int. J. Mol. Sci. 2022, 23, 7420. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Shen, Y.; Wang, Y.; Huang, Y.; Wang, J.; Zhu, Q.; Tong, G.; Yu, K.; Cao, W.; Wang, Q.; et al. Ferroptosis participates in neuron damage in experimental cerebral malaria and is partially induced by activated CD8+T cells. Mol. Brain 2022, 15, 57. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Yuan, Q.; Du, Z.; Fei, M.; Zhang, Q.; Yang, L.; Wang, M.; Yang, W.; Yu, J.; Wu, G.; et al. Ferroptosis in brain microvascular endothelial cells mediates blood-brain barrier disruption after traumatic brain injury. Biochem. Biophys. Res. Commun. 2022, 619, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, D.C.; Brüne, B. A graphical journey through iron metabolism, micrornas, and hypoxia in ferroptosis. Redox Biol. 2022, 54, 102365. [Google Scholar] [CrossRef]
- Shi, Y.; Han, L.; Zhang, X.; Xie, L.; Pan, P.; Chen, F. Selenium alleviates cerebral ischemia/reperfusion injury by regulating oxidative stress, mitochondrial fusion and ferroptosis. Neurochem. Res. 2022, 47, 2992–3002. [Google Scholar] [CrossRef]
- Moholkar, D.N.; Sadalage, P.S.; Peixoto, D.; Paiva-Santos, A.C.; Pawar, K.D. Recent advances in biopolymer-based formulations for wound healing applications. Eur. Polym. J. 2021, 160, 110784. [Google Scholar] [CrossRef]
- Kim, D.; Ku, B.; Choi, E.M. Se-methylselenocysteine stimulates migration and antioxidant response in HaCaT keratinocytes: Implications for wound healing. J. Trace Elem. Med. Biol. 2020, 58, 126426. [Google Scholar] [CrossRef]
- Zhong, J.; Wang, H.; Yang, K.; Wang, H.; Duan, C.; Ni, N.; An, L.; Luo, Y.; Zhao, P.; Gou, Y.; et al. Reversibly immortalized keratinocytes (iKera) facilitate re-epithelization and skin wound healing: Potential applications in cell-based skin tissue engineering. Bioact. Mater. 2022, 9, 523–540. [Google Scholar] [CrossRef]
- Haensel, D.; Dai, X. Epithelial-to-mesenchymal transition in cutaneous wound healing: Where we are and where we are heading. Dev. Dyn. 2018, 247, 473–480. [Google Scholar] [CrossRef]
- Tardáguila-García, A.; García-Morales, E.; García-Alamino, J.M.; Álvaro-Afonso, F.J.; Molines-Barroso, R.J.; Lázaro-Martínez, J.L. Metalloproteinases in chronic and acute wounds: A systematic review and meta-analysis. Wound Repair Regen. 2019, 27, 415–420. [Google Scholar] [CrossRef]
- Victor, P.; Sarada, D.; Ramkumar, K.M. Pharmacological activation of Nrf2 promotes wound healing. Eur. J. Pharmacol. 2020, 886, 173395. [Google Scholar] [CrossRef]
- Zhang, Y.; Roh, Y.J.; Han, S.-J.; Park, I.; Lee, H.M.; Ok, Y.S.; Lee, B.C.; Lee, S.-R. Role of selenoproteins in redox regulation of signaling and the antioxidant system: A review. Antioxidants 2020, 9, 383. [Google Scholar] [CrossRef]
- Vozza, G.; Khalid, M.; Byrne, H.J.; Ryan, S.M.; Frias, J.M. Nutraceutical formulation, characterisation, and in-vitro evaluation of methylselenocysteine and selenocystine using food derived chitosan: Zein nanoparticles. Food Res. Int. 2019, 120, 295–304. [Google Scholar] [CrossRef]
- Nasim, M.J.; Zuraik, M.M.; Abdin, A.Y.; Ney, Y.; Jacob, C. Selenomethionine: A pink trojan redox horse with implications in aging and various age-related diseases. Antioxidants 2021, 10, 882. [Google Scholar] [CrossRef]
- Alcântara, D.B.; Dionísio, A.P.; Artur, A.G.; Silveira, B.K.S.; Lopes, A.F.; Guedes, J.A.C.; Luz, L.R.; Nascimento, R.F.; Lopes, G.S.; Hermsdorff, H.H.M.; et al. Selenium in brazil nuts: An overview of agronomical aspects, recent trends in analytical chemistry, and health outcomes. Food Chem. 2022, 372, 131207. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, B.; Deng, K.; Gao, X.; Sun, G.; Zhang, Z.; Li, P.; Wang, W.; Li, H.; Zhang, Z.; et al. NRT1.1B improves selenium concentrations in rice grains by facilitating selenomethinone translocation. Plant Biotechnol. J. 2019, 17, 1058–1068. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, J.; Cheng, Y.; Chen, M.; Zhang, G.; Wu, J. Selenomethionine-dominated selenium-enriched peanut protein ameliorates alcohol-induced liver disease in mice by suppressing oxidative stress. Foods 2021, 10, 2979. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S. Current knowledge on the importance of selenium in food for living organisms: A review. Molecules 2016, 21, 609. [Google Scholar] [CrossRef]
- Santos, L.R.; Neves, C.; Melo, M.; Soares, P. Selenium and selenoproteins in immune mediated thyroid disorders. Diagnostics 2018, 8, 70. [Google Scholar] [CrossRef]
- Gorini, F.; Sabatino, L.; Pingitore, A.; Vassalle, C. Selenium: An element of life essential for thyroid function. Molecules 2021, 26, 7084. [Google Scholar] [CrossRef]
- Ragusa, F.; Fallahi, P.; Elia, G.; Gonnella, D.; Paparo, S.R.; Giusti, C.; Churilov, L.P.; Ferrari, S.M.; Antonelli, A. Hashimotos’ thyroiditis: Epidemiology, pathogenesis, clinic and therapy. Best Pract. Res. Clin. Endocrinol. Metab. 2019, 33, 101367. [Google Scholar] [CrossRef]
- Ventura, M.; Melo, M.; Carrilho, F. Selenium and thyroid function. In Selenium; Michalke, B., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 157–173. [Google Scholar]
- Antonelli, A.; Ferrari, S.M.; Ragusa, F.; Elia, G.; Paparo, S.R.; Ruffilli, I.; Patrizio, A.; Giusti, C.; Gonnella, D.; Cristaudo, A.; et al. Graves’ disease: Epidemiology, genetic and environmental risk factors and viruses. Best Pract. Res. Clin. Endocrinol. Metab. 2020, 34, 101387. [Google Scholar] [CrossRef]
- Ventura, M.; Melo, M.; Carrilho, F. Selenium and thyroid disease: From pathophysiology to treatment. Int. J. Endocrinol. 2017, 2017, 1297658. [Google Scholar] [CrossRef]
- Winther, K.H.; Rayman, M.P.; Bonnema, S.J.; Hegedus, L. Selenium in thyroid disorders—Essential knowledge for clinicians. Nat. Rev. Endocrinol. 2020, 16, 165–176. [Google Scholar] [CrossRef]
- Ye, R.; Huang, J.; Wang, Z.; Chen, Y.; Dong, Y. The role and mechanism of essential selenoproteins for homeostasis. Antioxidants 2022, 11, 973. [Google Scholar] [CrossRef]
- Davcheva, D.M.; Kirova, G.K.; Miteva, M.Z.; Tzvetkova, T.Z.; Orbetzova, M.M.; Nonchev, B.I.; Simitchiev, K.K.; Kmetov, V.J. Serum selenium concentration in patients with autoimmune thyroid disease. Folia Med. 2022, 64, 443–449. [Google Scholar] [CrossRef]
- Kochman, J.; Jakubczyk, K.; Bargiel, P.; Janda-Milczarek, K. The influence of oxidative stress on thyroid diseases. Antioxidants 2021, 10, 1442. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, R.M.; CampennÌ, A.; Giuffrida, G.; Casciaro, M.; Barbalace, M.C.; Hrelia, S.; Trimarchi, F.; CannavÒ, S.; Gangemi, S. Oxidative stress as a key feature of autoimmune thyroiditis: An update. Minerva Endocrinol. 2020, 45, 326–344. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Li, N.; Su, R.; Dai, C.; Zhang, R. Selenium supplementation may decrease thyroid peroxidase antibody titer via reducing oxidative stress in euthyroid patients with autoimmune thyroiditis. Int. J. Endocrinol. 2020, 2020, 9210572. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.M.; Ragusa, F.; Elia, G.; Paparo, S.R.; Mazzi, V.; Baldini, E.; Benvenga, S.; Antonelli, A.; Fallahi, P. Precision medicine in autoimmune thyroiditis and hypothyroidism. Front. Pharmacol. 2021, 12, 750380. [Google Scholar] [CrossRef]
- Savastano, G.; Del Negro, V.; Pompeo, D.; Sorrenti, S.; Galoppi, P.; Brunelli, R.; Piccioni, M.G. Thyroid disease, pregnancy, and selenium supplementation. Curr. Obstet. Gynecol. Rep. 2021, 10, 115–123. [Google Scholar] [CrossRef]
- Filipowicz, D.; Majewska, K.; Kalantarova, A.; Szczepanek-Parulska, E.; Ruchała, M. The rationale for selenium supplementation in patients with autoimmune thyroiditis, according to the current state of knowledge. Endokrynol. Pol. 2021, 72, 153–162. [Google Scholar] [CrossRef]
- Cannarella, R.; Condorelli, R.A.; Calogero, A.E.; Bagnara, V.; Aversa, A.; Greco, E.A.; Brunetti, A.; La Vignera, S. Effects of selenium supplementation on sperm parameters and DNA-fragmentation rate in patients with chronic autoimmune thyroiditis. J. Clin. Med. 2021, 10, 3755. [Google Scholar] [CrossRef]
- Ferrari, S.M.; Fallahi, P.; Di Bari, F.; Vita, R.; Benvenga, S.; Antonelli, A. Myo-inositol and selenium reduce the risk of developing overt hypothyroidism in patients with autoimmune thyroiditis. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 36–42. [Google Scholar]
- Pace, C.; Tumino, D.; Russo, M.; Le Moli, R.; Naselli, A.; Borzì, G.; Malandrino, P.; Frasca, F. Role of selenium and myo-inositol supplementation on autoimmune thyroiditis progression. Endocr. J. 2020, 67, 1093–1098. [Google Scholar] [CrossRef]
- Benvenga, S.; Ferrari, S.M.; Elia, G.; Ragusa, F.; Patrizio, A.; Paparo, S.R.; Camastra, S.; Bonofiglio, D.; Antonelli, A.; Fallahi, P. Nutraceuticals in thyroidology: A review of in vitro, and in vivo animal studies. Nutrients 2020, 12, 1337. [Google Scholar] [CrossRef]
- Benvenga, S.; Elia, G.; Ragusa, F.; Paparo, S.R.; Sturniolo, M.M.; Ferrari, S.M.; Antonelli, A.; Fallahi, P. Endocrine disruptors and thyroid autoimmunity. Best Pract. Res. Clin. Endocrinol. Metab. 2020, 34, 101377. [Google Scholar] [CrossRef]
- Kyrgios, I.; Giza, S.; Kotanidou, E.P.; Kleisarchaki, A.; Tsinopoulou, V.R.; Papadopoulou, A.; Markantonatou, A.M.; Kanellidou, E.; Giannakou, A.; Galli-Tsinopoulou, A. L-selenomethionine supplementation in children and adolescents with autoimmune thyroiditis: A randomized double-blind placebo-controlled clinical trial. J. Clin. Pharm. Ther. 2019, 44, 102–108. [Google Scholar] [CrossRef]
- Mantovani, G.; Isidori, A.M.; Moretti, C.; Di Dato, C.; Greco, E.; Ciolli, P.; Bonomi, M.; Petrone, L.; Fumarola, A.; Campagna, G.; et al. Selenium supplementation in the management of thyroid autoimmunity during pregnancy: Results of the “SERENA study”, a randomized, double-blind, placebo-controlled trial. Endocrine 2019, 66, 542–550. [Google Scholar] [CrossRef]
- Minnetti, M.; Sada, V.; Feola, T.; Giannetta, E.; Pozza, C.; Gianfrilli, D.; Isidori, A.M.; Cozzolino, A. Selenium supplementation in pregnant women with autoimmune thyroiditis: A practical approach. Nutrients 2022, 14, 2234. [Google Scholar] [CrossRef]
- Kryczyk-Koziol, J.; Zagrodzki, P.; Prochownik, E.; Blazewska-Gruszczyk, A.; Slowiaczek, M.; Sun, Q.; Schomburg, L.; Ochab, E.; Bartyzel, M. Positive effects of selenium supplementation in women with newly diagnosed Hashimoto’s thyroiditis in an area with low selenium status. Int. J. Clin. Pract. 2021, 75, e14484. [Google Scholar] [CrossRef] [PubMed]
- Krysiak, R.; Kowalcze, K.; Okopień, B. The effect of selenomethionine on thyroid autoimmunity in euthyroid men with Hashimoto thyroiditis and testosterone deficiency. J. Clin. Pharmacol. 2019, 59, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Krysiak, R.; Szkróbka, W.; Okopień, B. The effect of vitamin D and selenomethionine on thyroid antibody titers, hypothalamic-pituitary-thyroid axis activity and thyroid function tests in men with Hashimoto’s thyroiditis: A pilot study. Pharmacol. Rep. 2019, 71, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Pirola, I.; Rotondi, M.; Cristiano, A.; Maffezzoni, F.; Pasquali, D.; Marini, F.; Coperchini, F.; Paganelli, M.; Apostoli, P.; Chiovato, L.; et al. Selenium supplementation in patients with subclinical hypothyroidism affected by autoimmune thyroiditis: Results of the SETI study. Endocrinol. Diabetes Nutr. 2020, 67, 28–35. [Google Scholar] [CrossRef]
- Bartalena, L.; Kahaly, G.J.; Baldeschi, L.; Dayan, C.M.; Eckstein, A.; Marcocci, C.; Marinò, M.; Vaidya, B.; Wiersinga, W.M. The 2021 European Group on Graves’ orbitopathy (EUGOGO) clinical practice guidelines for the medical management of Graves’ orbitopathy. Eur. J. Endocrinol. 2021, 185, G43–G67. [Google Scholar] [CrossRef]
- Korbut, E.; Ptak-Belowska, A.; Brzozowski, T. Inhibitory effect of selenomethionine on carcinogenesis in the model of human colorectal cancer in vitro and its link to the Wnt/β-catenin pathway. Acta Biochim. Pol. 2018, 65, 359–366. [Google Scholar] [CrossRef]
- Pons, D.G.; Moran, C.; Alorda-Clara, M.; Oliver, J.; Roca, P.; Sastre-Serra, J. Micronutrients selenomethionine and selenocysteine modulate the redox status of MCF-7 breast cancer cells. Nutrients 2020, 12, 865. [Google Scholar] [CrossRef]
- Dai, X.; Thongchot, S.; Dokduang, H.; Loilome, W.; Khuntikeo, N.; Titapun, A.; Ungarreevittaya, P.; Yongvanit, P.; Techasen, A.; Namwat, N. Potential of selenium compounds as new anticancer agents for cholangiocarcinoma. Anticancer Res. 2016, 36, 5981. [Google Scholar] [CrossRef]
- Zhao, M.-J.; Yuan, S.; Zi, H.; Gu, J.-M.; Fang, C.; Zeng, X.-T. Oxidative stress links aging-associated cardiovascular diseases and prostatic diseases. Oxid. Med. Cell. Longev. 2021, 2021, 5896136. [Google Scholar] [CrossRef]
- Steven, S.; Frenis, K.; Oelze, M.; Kalinovic, S.; Kuntic, M.; Bayo Jimenez, M.T.; Vujacic-Mirski, K.; Helmstädter, J.; Kröller-Schön, S.; Münzel, T.; et al. Vascular inflammation and oxidative stress: Major triggers for cardiovascular disease. Oxid. Med. Cell. Longev. 2019, 2019, 7092151. [Google Scholar] [CrossRef]
- Dubois-Deruy, E.; Peugnet, V.; Turkieh, A.; Pinet, F. Oxidative stress in cardiovascular diseases. Antioxidants 2020, 9, 864. [Google Scholar] [CrossRef]
- Sha Zhao, C.K.C.; Zhang, C.-L.; Huang, Y. Interplay between oxidative stress, cyclooxygenases, and prostanoids in cardiovascular diseases. Antioxid. Redox Signal. 2021, 34, 784–799. [Google Scholar] [CrossRef]
- Reyes, L.; Bishop, D.P.; Hawkins, C.L.; Rayner, B.S. Assessing the efficacy of dietary selenomethionine supplementation in the setting of cardiac ischemia/reperfusion injury. Antioxidants 2019, 8, 546. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Wang, W.; Kang, P.M. Oxidative stress and antioxidant treatments in cardiovascular diseases. Antioxidants 2020, 9, 1292. [Google Scholar] [CrossRef]
- Flouda, K.; Gammelgaard, B.; Davies, M.J.; Hawkins, C.L. Modulation of hypochlorous acid (HOCl) induced damage to vascular smooth muscle cells by thiocyanate and selenium analogues. Redox Biol. 2021, 41, 101873. [Google Scholar] [CrossRef]
- Carroll, L.; Pattison, D.I.; Fu, S.; Schiesser, C.H.; Davies, M.J.; Hawkins, C.L. Catalytic oxidant scavenging by selenium-containing compounds: Reduction of selenoxides and N-chloramines by thiols and redox enzymes. Redox Biol. 2017, 12, 872–882. [Google Scholar] [CrossRef]
- Nogueira, C.W.; Barbosa, N.V.; Rocha, J.B.T. Toxicology and pharmacology of synthetic organoselenium compounds: An update. Arch. Toxicol. 2021, 95, 1179–1226. [Google Scholar] [CrossRef]
- Zhang, Y.; Cartland, S.P.; Henriquez, R.; Patel, S.; Gammelgaard, B.; Flouda, K.; Hawkins, C.L.; Rayner, B.S. Selenomethionine supplementation reduces lesion burden, improves vessel function and modulates the inflammatory response within the setting of atherosclerosis. Redox Biol. 2020, 29, 101409. [Google Scholar] [CrossRef]
- Karaye, K.M.; Sa’idu, H.; Balarabe, S.A.; Ishaq, N.A.; Sanni, B.; Abubakar, H.; Mohammed, B.L.; Abdulsalam, T.; Tukur, J.; Mohammed, I.Y. Selenium supplementation in patients with peripartum cardiomyopathy: A proof-of-concept trial. BMC Cardiovasc. Disord. 2020, 20, 457. [Google Scholar] [CrossRef]
- Agrawal, A.; Jain, D.; Ram, P.; Leon, J.L.P.; Rangaswami, J. Anticoagulation for intra-cardiac thrombi in peripartum cardiomyopathy: A review of the literature. RCM 2019, 20, 53–58. [Google Scholar]
- Gupta, D.; Wenger, N.K. Peripartum cardiomyopathy: Status 2018. Clin. Cardiol. 2018, 41, 217–219. [Google Scholar] [CrossRef]
- Nadatani, Y.; Watanabe, T.; Shimada, S.; Otani, K.; Tanigawa, T.; Fujiwara, Y. Microbiome and intestinal ischemia/reperfusion injury. J. Clin. Biochem. Nutr. 2018, 63, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Barzegar, M.; Kaur, G.; Gavins, F.N.E.; Wang, Y.; Boyer, C.J.; Alexander, J.S. Potential therapeutic roles of stem cells in ischemia-reperfusion injury. Stem Cell Res. 2019, 37, 101421. [Google Scholar] [CrossRef]
- Li, G.; Wang, S.; Fan, Z. Oxidative stress in intestinal ischemia-reperfusion. Front. Med. 2022, 8, 750731. [Google Scholar] [CrossRef]
- Liu, Z.; Mou, G.; Liang, Z.; Zhao, R.; Jin, C.; Wu, R. Selenomethionine alleviates intestinal ischemia-reperfusion injury in mice through the Bax-caspase pathway. Biol. Trace Elem. Res. 2021, 200, 3205–3214. [Google Scholar] [CrossRef]
- Li, G.; Zhang, Y.; Fan, Z. Cellular signal transduction pathways involved in acute lung injury induced by intestinal ischemia-reperfusion. Oxid. Med. Cell. Longev. 2021, 2021, 9985701. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Sun, R.; Wang, G.; Chen, Z.; Li, Y.; Zhao, Y.; Liu, D.; Zhao, H.; Zhang, F.; Yao, J.; et al. SIRT3-mediated deacetylation of PRDX3 alleviates mitochondrial oxidative damage and apoptosis induced by intestinal ischemia/reperfusion injury. Redox Biol. 2020, 28, 101343. [Google Scholar] [CrossRef] [PubMed]
- Chain, E. Panel o.C.i.t.F.; Schrenk, D.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.-C.; Nebbia, C.S.; et al. Risk assessment of ochratoxin A in food. EFSA J. 2020, 18, e06113. [Google Scholar]
- Wang, L.; Hua, X.; Shi, J.; Jing, N.; Ji, T.; Lv, B.; Liu, L.; Chen, Y. Ochratoxin A: Occurrence and recent advances in detoxification. Toxicon 2022, 210, 11–18. [Google Scholar] [CrossRef]
- Khoi, C.S.; Chen, J.H.; Lin, T.Y.; Chiang, C.K.; Hung, K.Y. Ochratoxin A-induced nephrotoxicity: Up-to-date evidence. Int. J. Mol. Sci. 2021, 22, 11237. [Google Scholar] [CrossRef]
- Stoev, S.D. Balkan endemic nephropathy—Still continuing enigma, risk assessment and underestimated hazard of joint mycotoxin exposure of animals or humans. Chem. Biol. Interact. 2017, 261, 63–79. [Google Scholar] [CrossRef]
- Mao, X.; Li, H.; Ge, L.; Liu, S.; Hou, L.; Yue, D.; Du, H.; Pan, C.; Gan, F.; Liu, Y.; et al. Selenomethionine alleviated ochratoxin A induced pyroptosis and renal fibrotic factors expressions in MDCK cells. J. Biochem. Mol. Toxicol. 2022, 36, e22933. [Google Scholar] [CrossRef]
- Lan, M.; Zhang, Y.; Wan, X.; Pan, M.-H.; Xu, Y.; Sun, S.-C. Melatonin ameliorates ochratoxin A-induced oxidative stress and apoptosis in porcine oocytes. Environ. Pollut. 2020, 256, 113374. [Google Scholar] [CrossRef]
- Li, H.; Malyar, R.M.; Zhai, N.; Wang, H.; Liu, K.; Liu, D.; Pan, C.; Gan, F.; Huang, K.; Miao, J.; et al. Zinc supplementation alleviates OTA-induced oxidative stress and apoptosis in MDCK cells by up-regulating metallothioneins. Life Sci. 2019, 234, 116735. [Google Scholar] [CrossRef]
- Hou, L.; Zhou, X.; Gan, F.; Liu, Z.; Zhou, Y.; Qian, G.; Huang, K. Combination of selenomethionine and N-acetylcysteine alleviates the joint toxicities of aflatoxin B1 and ochratoxin A by ERK MAPK signal pathway in porcine alveolar macrophages. J. Agric. Food Chem. 2018, 66, 5913–5923. [Google Scholar] [CrossRef]
- Le, G.; Yang, L.; Du, H.; Hou, L.; Ge, L.; Sylia, A.; Muhmood, A.; Chen, X.; Han, B.; Huang, K. Combination of zinc and selenium alleviates ochratoxin A-induced fibrosis via blocking ROS-dependent autophagy in HK-2 cells. J. Trace Elem. Med. Biol. 2022, 69, 126881. [Google Scholar] [CrossRef]
- Hou, L.; Lin, Z.; Xu, A.; Le, G.; Ge, L.; Liu, S.; Muhmood, A.; Gan, F.; Huang, K. Combined protective effects of icariin and selenomethionine on novel chronic tubulointerstitial nephropathy models in vivo and in vitro. Br. J. Nutr. 2022, 127, 12–22. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Wu, Q.Y.; Chen, C.; Zheng, R.; Chen, Y.; Ni, J.Z.; Song, G.L. Comparison of the effects of selenomethionine and selenium-enriched yeast in the triple-transgenic mouse model of Alzheimer’s disease. Food Funct. 2018, 9, 3965–3973. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Chen, C.; Wu, Q.Y.; Zheng, R.; Liu, Q.; Ni, J.Z.; Hoffmann, P.R.; Song, G.L. Selenomethionine reduces the deposition of beta-amyloid plaques by modulating β-secretase and enhancing selenoenzymatic activity in a mouse model of Alzheimer’s disease. Metallomics 2016, 8, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Zhang, Z.-H.; Chen, C.; Chen, Y.; Jia, S.-Z.; Liu, Q.; Ni, J.-Z.; Song, G.-L. Selenomethionine promoted hippocampal neurogenesis via the PI3K-Akt-GSK3β-Wnt pathway in a mouse model of Alzheimer’s disease. Biochem. Biophys. Res. Commun. 2017, 485, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Wu, Q.Y.; Chen, C.; Zheng, R.; Chen, Y.; Liu, Q.; Ni, J.Z.; Song, G.L. Selenomethionine attenuates the amyloid-β level by both inhibiting amyloid-β production and modulating autophagy in Neuron-2a/AβPPswe cells. J. Alzheimers Dis. 2017, 59, 591–602. [Google Scholar] [CrossRef] [PubMed]
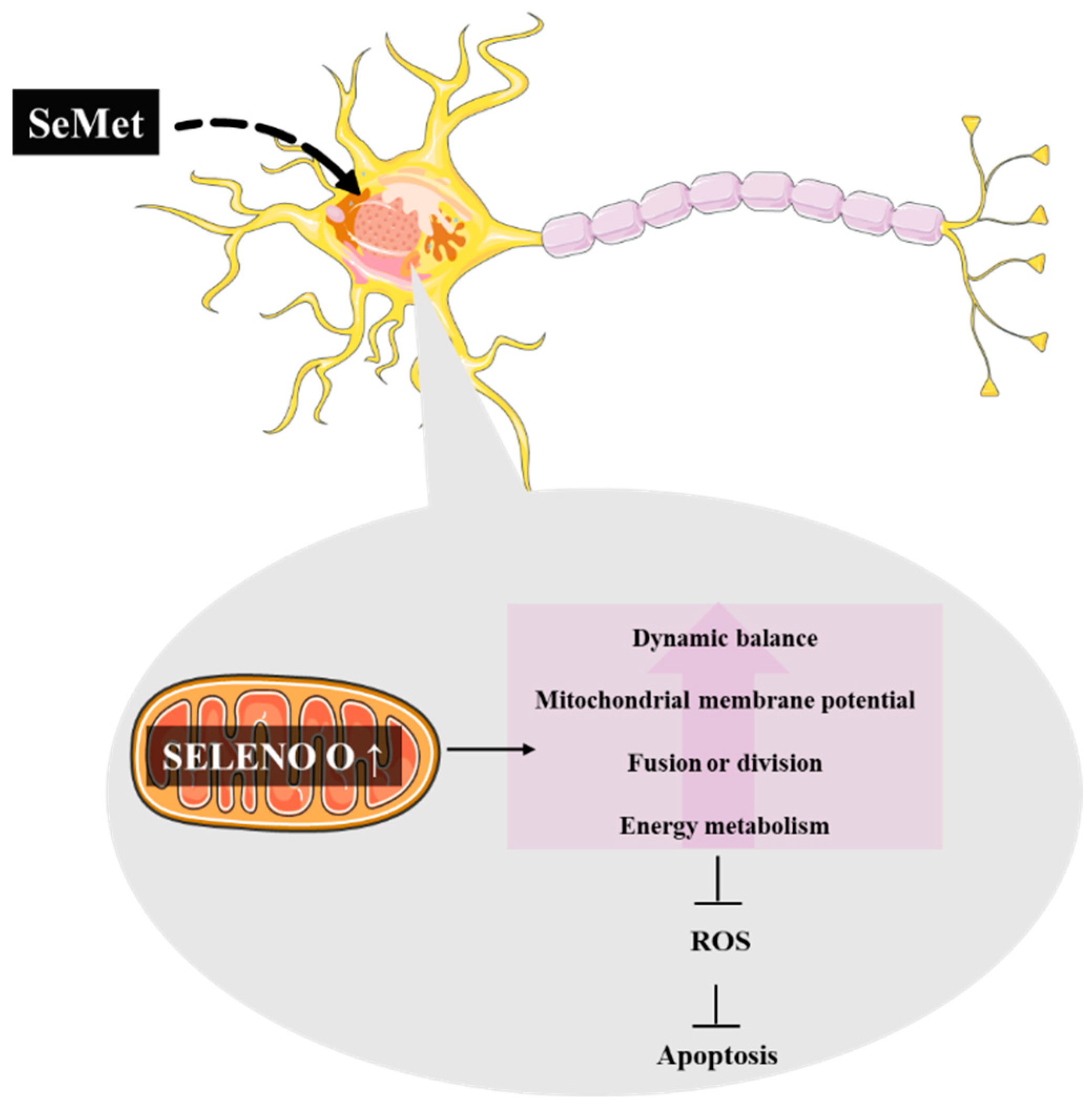
- Chen, C.; Chen, Y.; Zhang, Z.H.; Jia, S.Z.; Chen, Y.B.; Huang, S.L.; Xu, X.W.; Song, G.L. Selenomethionine improves mitochondrial function by upregulating mitochondrial selenoprotein in a model of Alzheimer’s disease. Front. Aging Neurosci. 2021, 13, 750921. [Google Scholar] [CrossRef] [PubMed]
- Moradi, H.R.; Hajali, V.; Khaksar, Z.; Vafaee, F.; Forouzanfar, F.; Negah, S.S. The next step of neurogenesis in the context of Alzheimer’s disease. Mol. Biol. Rep. 2021, 48, 5647–5660. [Google Scholar] [CrossRef] [PubMed]
- Sung, P.-S.; Lin, P.-Y.; Liu, C.-H.; Su, H.-C.; Tsai, K.-J. Neuroinflammation and neurogenesis in Alzheimer’s disease and potential therapeutic approaches. Int. J. Mol. Sci. 2020, 21, 701. [Google Scholar] [CrossRef]
- Essa, H.; Peyton, L.; Hasan, W.; León, B.E.; Choi, D.-S. Implication of adult hippocampal neurogenesis in Alzheimer’s disease and potential therapeutic approaches. Cells 2022, 11, 286. [Google Scholar] [CrossRef]
- Zheng, J. Hippocampal neurogenesis and pro-neurogenic therapies for Alzheimer’s disease. Anim. Model. Exp. Med. 2022, 5, 3–14. [Google Scholar] [CrossRef]
- Muzembo, B.A.; Ngatu, N.R.; Januka, K.; Huang, H.L.; Nattadech, C.; Suzuki, T.; Wada, K.; Ikeda, S. Selenium supplementation in HIV-infected individuals: A systematic review of randomized controlled trials. Clin. Nutr. ESPEN 2019, 34, 1–7. [Google Scholar] [CrossRef]
- Qian, G.; Liu, D.; Hu, J.; Gan, F.; Hou, L.; Zhai, N.; Chen, X.; Huang, K. SeMet attenuates OTA-induced PCV2 replication promotion by inhibiting autophagy by activating the AKT/mTOR signaling pathway. Vet. Res. 2018, 49, 15. [Google Scholar] [CrossRef]
- Ren, Z.; Jia, G.; He, H.; Ding, T.; Yu, Y.; Zuo, Z.; Hu, Y.; Zhong, Z.; Yu, S.; Deng, H.; et al. Antiviral effect of selenomethionine on porcine deltacoronavirus in pig kidney epithelial cells. Front. Microbiol. 2022, 13, 846747. [Google Scholar] [CrossRef]
- Pecoraro, B.M.; Leal, D.F.; Frias-De-Diego, A.; Browning, M.; Odle, J.; Crisci, E. The health benefits of selenium in food animals: A review. J. Anim. Sci. Biotechnol. 2022, 13, 58. [Google Scholar] [CrossRef]
- Dound, Y.A.; Sehgal, R. Preclinical efficacy and safety studies of formulation SSV-003, a potent anti-viral herbal formulation. J. Exp. Pharmacol. 2021, 13, 913–921. [Google Scholar] [CrossRef]
- Sagaama, A.; Brandan, S.A.; Ben Issa, T.; Issaoui, N. Searching potential antiviral candidates for the treatment of the 2019 novel coronavirus based on DFT calculations and molecular docking. Heliyon 2020, 6, e04640. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morán-Serradilla, C.; Angulo-Elizari, E.; Henriquez-Figuereo, A.; Sanmartín, C.; Sharma, A.K.; Plano, D. Seleno-Metabolites and Their Precursors: A New Dawn for Several Illnesses? Metabolites 2022, 12, 874. https://doi.org/10.3390/metabo12090874
Morán-Serradilla C, Angulo-Elizari E, Henriquez-Figuereo A, Sanmartín C, Sharma AK, Plano D. Seleno-Metabolites and Their Precursors: A New Dawn for Several Illnesses? Metabolites. 2022; 12(9):874. https://doi.org/10.3390/metabo12090874
Chicago/Turabian StyleMorán-Serradilla, Cristina, Eduardo Angulo-Elizari, Andreina Henriquez-Figuereo, Carmen Sanmartín, Arun K. Sharma, and Daniel Plano. 2022. "Seleno-Metabolites and Their Precursors: A New Dawn for Several Illnesses?" Metabolites 12, no. 9: 874. https://doi.org/10.3390/metabo12090874