
Orthogonality in Principal Component Analysis Allows the Discovery of Lipids in the Jejunum That Are Independent of Ad Libitum Feeding

 , , , , , and
, , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
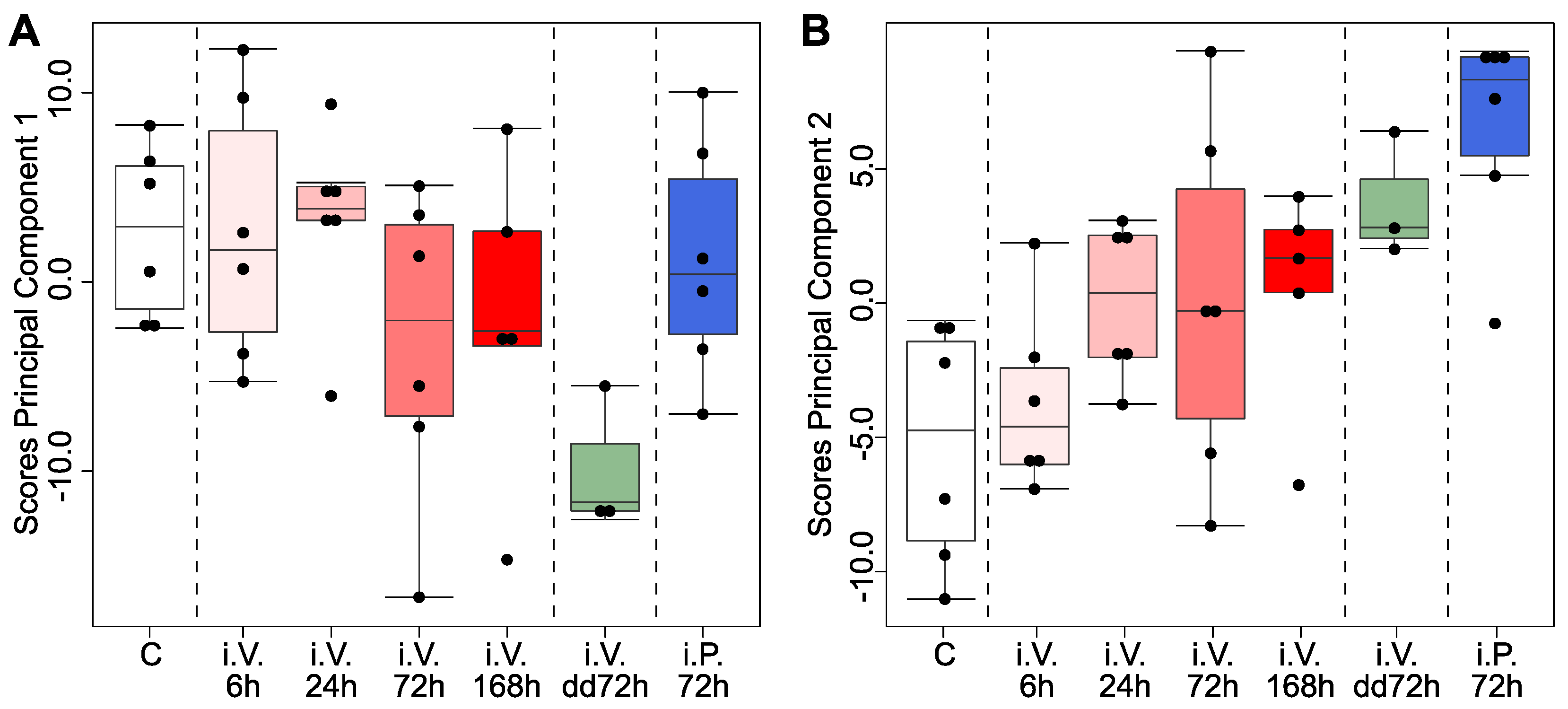
2.1. Principal Component Analysis Allowed the Identification of Lipids That Were Statistically Independent of Ad Libitum Feeding in the Jejunum (but Not in the Liver)
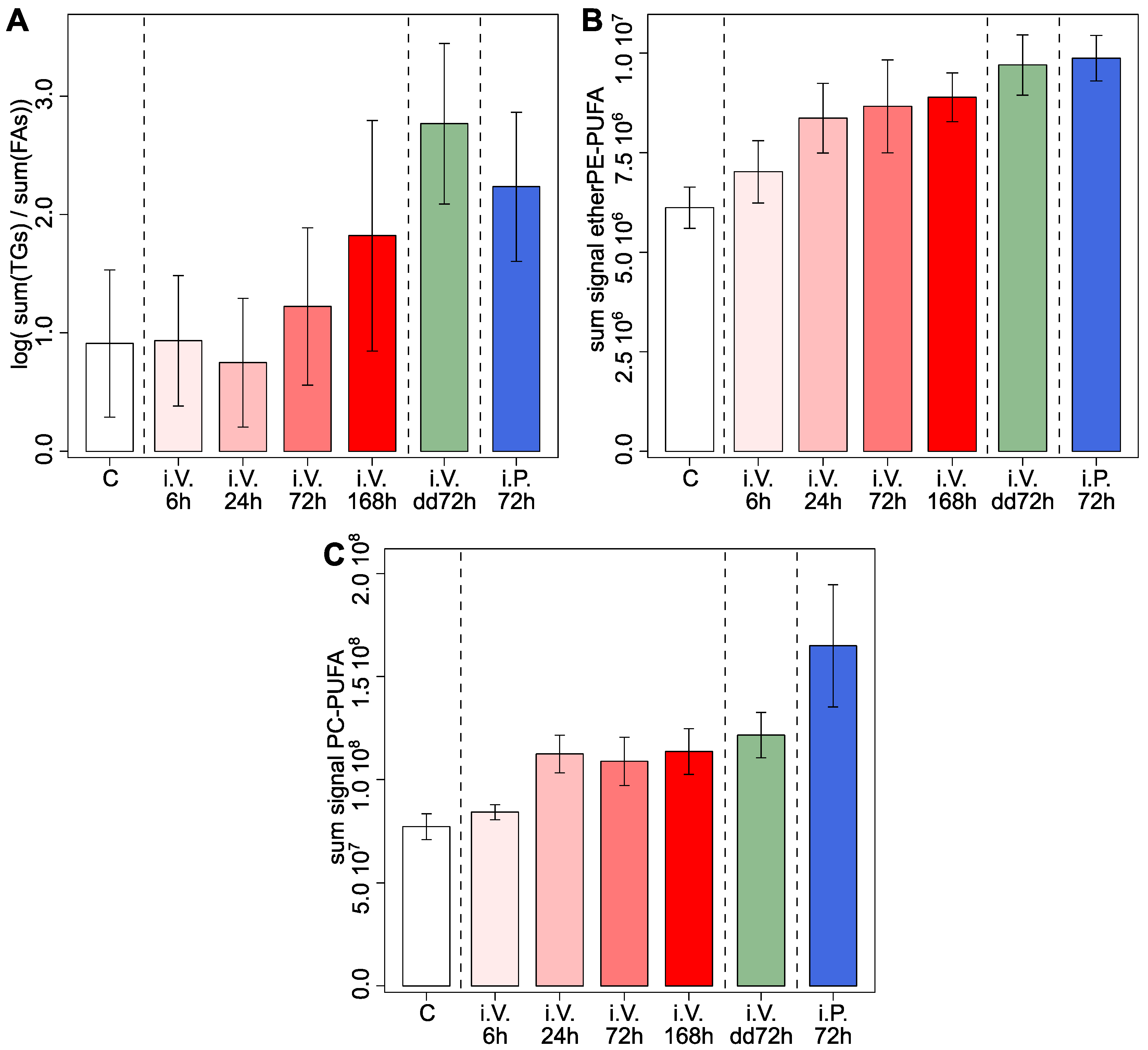
2.2. Doxorubicin Affects Feeding Biomarkers in the Jejunum and Increases Lipids with PUFAs, Which Is Consistent with Previous Results in Cell Cultures
3. Concluding Remarks
4. Materials and Methods
4.1. Animal Treatment, Tissue Isolation, and Tissue Homogenization
4.2. Lipidomics Analysis
4.3. Data Pre-Treatment, Analysis, and Intepretation
4.4. Software and Graphical Resources
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mu, H.; Høy, C.-E. The Digestion of Dietary Triacylglycerols. Prog. Lipid Res. 2004, 43, 105–133. [Google Scholar] [CrossRef]
- Balgoma, D.; Guitton, Y.; Evans, J.J.; Le Bizec, B.; Dervilly-Pinel, G.; Meynier, A. Modeling the Fragmentation Patterns of Triacylglycerides in Mass Spectrometry Allows the Quantification of the Regioisomers with a Minimal Number of Standards. Anal. Chim. Acta 2019, 1057, 60–69. [Google Scholar] [CrossRef]
- Nilsson, Å.; Duan, R.-D. Pancreatic and Mucosal Enzymes in Choline Phospholipid Digestion. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G425–G445. [Google Scholar] [CrossRef] [PubMed]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. (Eds.) An Introduction to Statistical Learning: With Applications in R; Springer Texts in Statistics; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-7137-0. [Google Scholar]
- Shlens, J. A Tutorial on Principal Component Analysis. arXiv 2014, arXiv:1404.1100. [Google Scholar]
- Rodionova, O.; Kucheryavskiy, S.; Pomerantsev, A. Efficient Tools for Principal Component Analysis of Complex Data—A Tutorial. Chemom. Intell. Lab. Syst. 2021, 213, 104304. [Google Scholar] [CrossRef]
- Balgoma, D.; Kullenberg, F.; Calitz, C.; Kopsida, M.; Heindryckx, F.; Lennernäs, H.; Hedeland, M. Anthracyclins Increase PUFAs: Potential Implications in ER Stress and Cell Death. Cells 2021, 10, 1163. [Google Scholar] [CrossRef]
- Zou, Y.; Henry, W.S.; Ricq, E.L.; Graham, E.T.; Phadnis, V.V.; Maretich, P.; Paradkar, S.; Boehnke, N.; Deik, A.A.; Reinhardt, F.; et al. Plasticity of Ether Lipids Promotes Ferroptosis Susceptibility and Evasion. Nature 2020, 585, 603–608. [Google Scholar] [CrossRef]
- Balgoma, D.; Hedeland, M. Etherglycerophospholipids and Ferroptosis: Structure, Regulation, and Location. Trends Endocrinol. Metab. 2021, 32, 960–962. [Google Scholar] [CrossRef]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascón, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Balgoma, D.; Zelleroth, S.; Grönbladh, A.; Hallberg, M.; Pettersson, C.; Hedeland, M. Anabolic Androgenic Steroids Exert a Selective Remodeling of the Plasma Lipidome That Mirrors the Decrease of the de Novo Lipogenesis in the Liver. Metabolomics 2020, 16, 12. [Google Scholar] [CrossRef] [Green Version]
- Pulfer, M.; Murphy, R.C. Electrospray Mass Spectrometry of Phospholipids. Mass Spectrom. Rev. 2003, 22, 332–364. [Google Scholar] [CrossRef]
- McAnoy, A.M.; Wu, C.C.; Murphy, R.C. Direct Qualitative Analysis of Triacylglycerols by Electrospray Mass Spectrometry Using a Linear Ion Trap. J. Am. Soc. Mass Spectrom. 2005, 16, 1498–1509. [Google Scholar] [CrossRef] [PubMed]
- Betensky, R.A. The P-Value Requires Context, Not a Threshold. Am. Stat. 2019, 73, 115–117. [Google Scholar] [CrossRef]
- Wasserstein, R.L.; Lazar, N.A. The ASA’s Statement on p -Values: Context, Process, and Purpose. Am. Stat. 2016, 70, 129–133. [Google Scholar] [CrossRef]
- Wasserstein, R.L.; Schirm, A.L.; Lazar, N.A. Moving to a World Beyond “p < 0.05”. Am. Stat. 2019, 73, 1–19. [Google Scholar] [CrossRef]
- Gardner, M.J.; Altman, D.G. Confidence Intervals Rather than P Values: Estimation Rather than Hypothesis Testing. Br. Med. J. Clin. Res. Ed. 1986, 292, 746–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balgoma, D.; Kullenberg, F.; Peters, K.; Dahlgren, D.; Heindryckx, F.; Lennernäs, H.; Hedeland, M. Orthogonality in Principal Component Analysis Allows the Discovery of Lipids in the Jejunum That Are Independent of Ad Libitum Feeding. Metabolites 2022, 12, 866. https://doi.org/10.3390/metabo12090866
Balgoma D, Kullenberg F, Peters K, Dahlgren D, Heindryckx F, Lennernäs H, Hedeland M. Orthogonality in Principal Component Analysis Allows the Discovery of Lipids in the Jejunum That Are Independent of Ad Libitum Feeding. Metabolites. 2022; 12(9):866. https://doi.org/10.3390/metabo12090866
Chicago/Turabian StyleBalgoma, David, Fredrik Kullenberg, Karsten Peters, David Dahlgren, Femke Heindryckx, Hans Lennernäs, and Mikael Hedeland. 2022. "Orthogonality in Principal Component Analysis Allows the Discovery of Lipids in the Jejunum That Are Independent of Ad Libitum Feeding" Metabolites 12, no. 9: 866. https://doi.org/10.3390/metabo12090866