
The Metabolomic Profile in Amyotrophic Lateral Sclerosis Changes According to the Progression of the Disease: An Exploratory Study
 , , , , , ,
, , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Sample Collection and Preparation
2.3. NMR Spectroscopy and Processing
2.4. Mass Spectrometry Sample Preparation and Processing
2.5. Statistical Analysis
3. Results
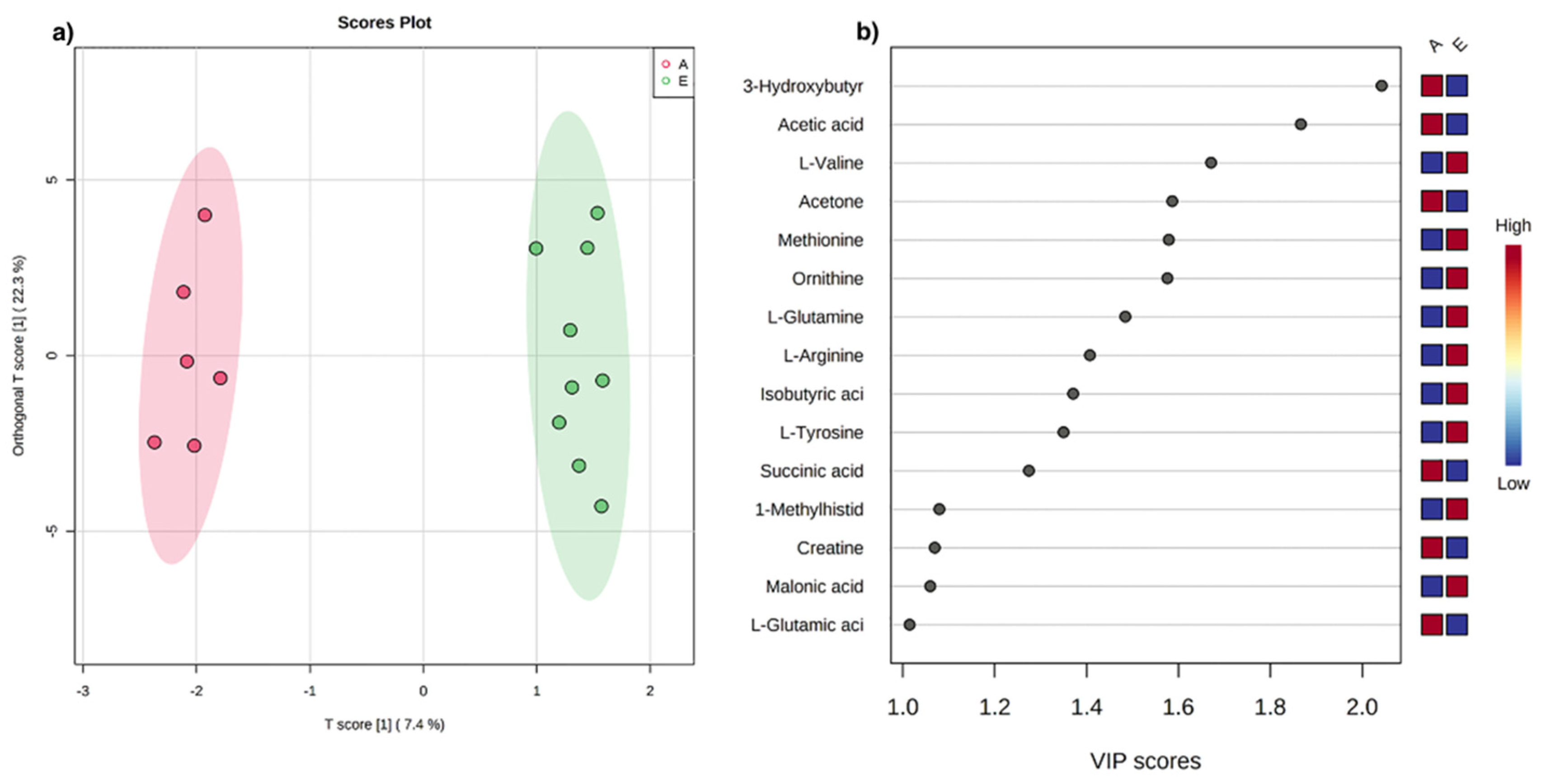
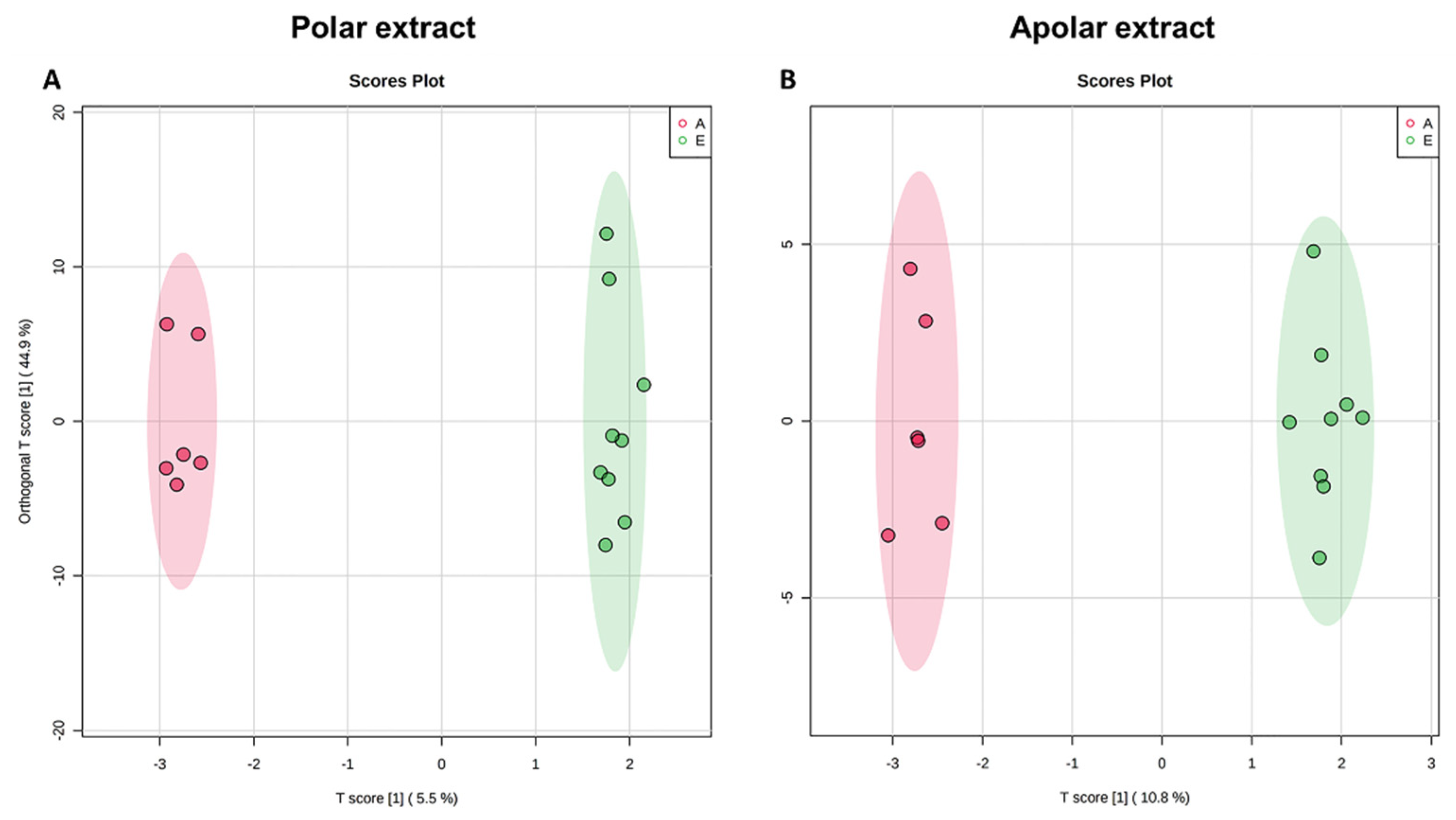
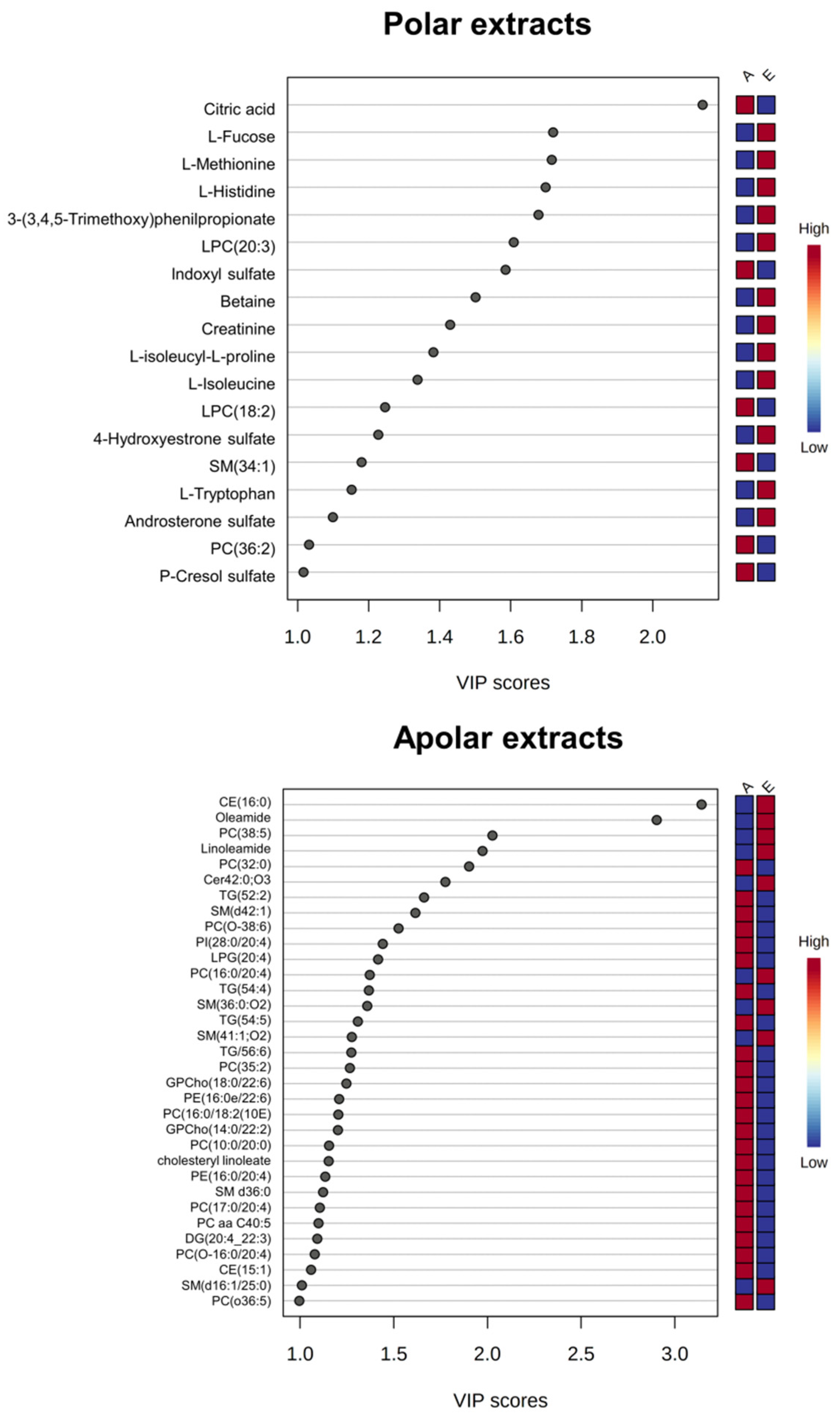
3.1. Univariate and Multivariate Data Analysis
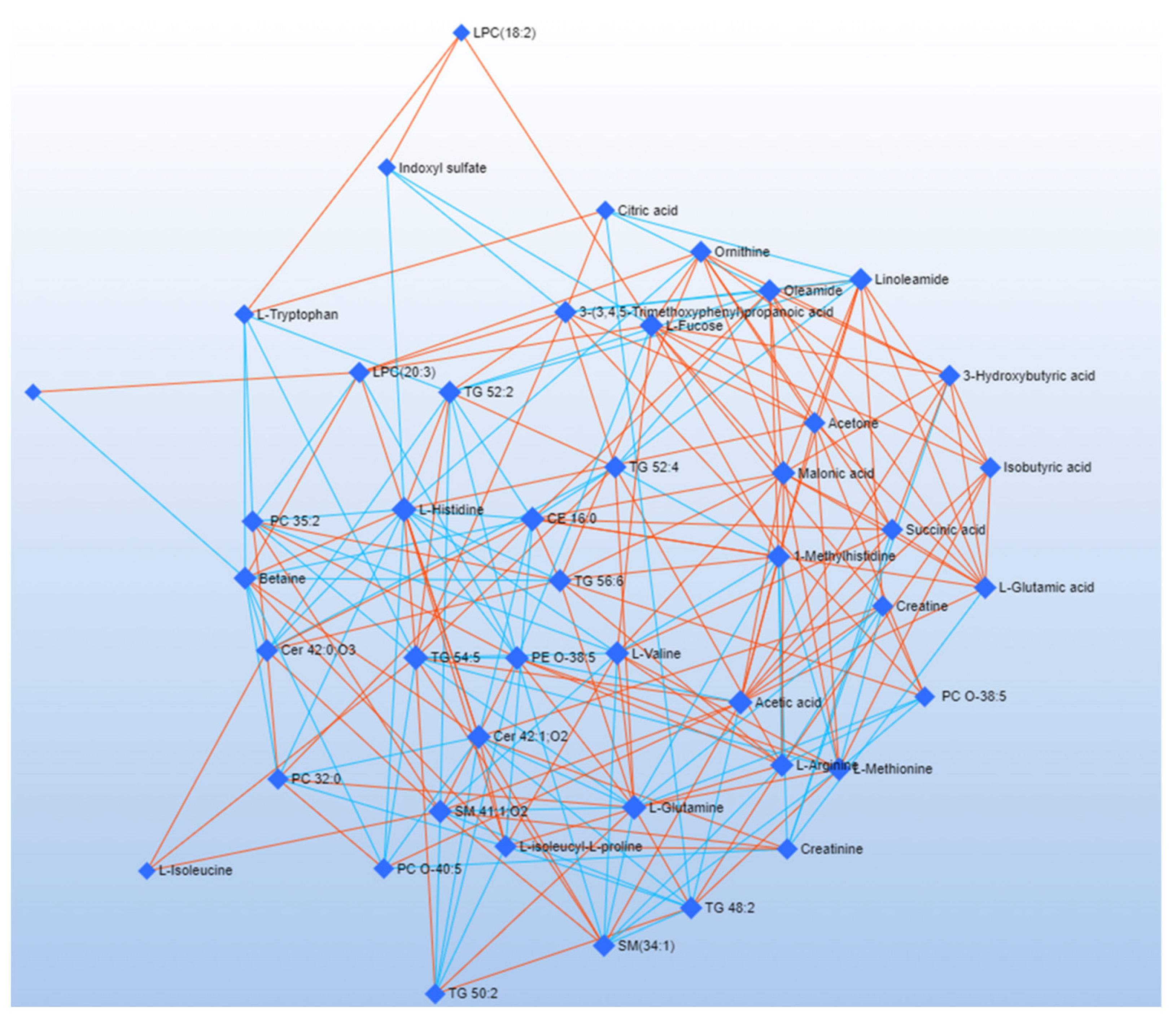
3.2. Debiased Sparse Partial Correlation (DSPC) Algorithm
3.3. Combined Pathway Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aydemir, D.; Ulusu, N.N. Importance of the serum biochemical parameters as potential biomarkers for rapid diagnosis and evaluating preclinical stage of ALS. Med. Hypotheses 2020, 141, 109736. [Google Scholar] [CrossRef]
- Foerster, B.R.; Pomper, M.G.; Callaghan, B.C.; Petrou, M.; Edden, R.; Mohamed, M.; Welsh, R.C.; Carlos, R.C.; Barker, P.B.; Feldman, E. An Imbalance Between Excitatory and Inhibitory Neurotransmitters in Amyotrophic Lateral Sclerosis Revealed by Use of 3-T Proton Magnetic Resonance Spectroscopy. JAMA Neurol. 2013, 70, 1009–1016. [Google Scholar] [CrossRef]
- McCombe, P.A.; Henderson, R.D. Effects of gender in amyotrophic lateral sclerosis. Gend. Med. 2010, 7, 557–570. [Google Scholar] [CrossRef]
- Takeda, T.; Kitagawa, K.; Arai, K. Phenotypic variability and its pathological basis in amyotrophic lateral sclerosis. Neuropathology 2020, 40, 40–56. [Google Scholar] [CrossRef]
- Shefner, J.M.; Al-Chalabi, A.; Baker, M.R.; Cui, L.-Y.; de Carvalho, M.; Eisen, A.; Grosskreutz, J.; Hardiman, O.; Henderson, R.; Matamala, J.M.; et al. A proposal for new diagnostic criteria for ALS. Clin. Neurophysiol. 2020, 131, 1975–1978. [Google Scholar] [CrossRef]
- Balendra, R.; Jones, A.; Jivraj, N.; Steen, I.N.; Young, C.; Shaw, P.; Turner, M.; Leigh, P.N.; Al-Chalabi, A. Use of clinical staging in amyotrophic lateral sclerosis for phase 3 clinical trials. J. Neurol. Neurosurg. Psychiatry 2015, 86, 45–49. [Google Scholar] [CrossRef]
- Poletti, B.; Solca, F.; Carelli, L.; Madotto, F.; Lafronza, A.; Faini, A.; Monti, A.; Zago, S.; Calini, D.; Tiloca, C.; et al. The validation of the Italian Edinburgh Cognitive and Behavioural ALS Screen (ECAS). Amyotroph. Lateral Scler. Front. Degener. 2016, 17, 489–498. [Google Scholar] [CrossRef]
- Cedarbaum, J.M.; Stambler, N.; Malta, E.; Fuller, C.; Hilt, D.; Thurmond, B.; Nakanishi, A. The ALSFRS-R: A revised ALS functional rating scale that incorporates assessments of respiratory function. BDNF ALS Study Group (Phase III). J. Neurol. Sci. 1999, 169, 13–21. [Google Scholar] [CrossRef]
- Siciliano, M.; Trojano, L.; Trojsi, F.; Greco, R.; Santoro, M.; Basile, G.; Piscopo, F.; D’Iorio, A.; Patrone, M.; Femiano, C.; et al. Edinburgh Cognitive and Behavioural ALS Screen (ECAS)-Italian version: Regression based norms and equivalent scores. Neurol. Sci. 2017, 38, 1059–1068. [Google Scholar] [CrossRef]
- Kim, H.-J.; De Leon, M.; Wang, X.; Kim, H.Y.; Lee, Y.-J.; Kim, Y.-H.; Kim, S.H. Relationship between Clinical Parameters and Brain Structure in Sporadic Amyotrophic Lateral Sclerosis Patients According to Onset Type: A Voxel-Based Morphometric Study. PLoS ONE 2017, 12, e0168424. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.-S.; Yuan, M. Considerations on the concept, definition, and diagnosis of amyotrophic lateral sclerosis. Neural Regen. Res. 2021, 16, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, P.; Rucco, R.; Jacini, F.; Trojsi, F.; Lardone, A.; Baselice, F.; Femiano, C.; Santangelo, G.; Granata, C.; Vettoliere, A.; et al. Brain functional networks become more connected as amyotrophic lateral sclerosis progresses: A source level magnetoencephalographic study. NeuroImage Clin. 2018, 20, 564–571. [Google Scholar] [CrossRef] [PubMed]
- A McCombe, P.; D Henderson, R. The role of immune and inflammatory mechanisms in ALS. Curr. Mol. Med. 2011, 11, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Robelin, L.; De Aguilar, J.L.G. Blood Biomarkers for Amyotrophic Lateral Sclerosis: Myth or Reality? BioMed Res. Int. 2014, 2014, 525097. [Google Scholar] [CrossRef]
- Polverino, A.; Rucco, R.; Stillitano, I.; Bonavita, S.; Grimaldi, M.; Minino, R.; Pesoli, M.; Trojsi, F.; D’Ursi, A.M.; Sorrentino, G.; et al. In Amyotrophic Lateral Sclerosis Blood Cytokines Are Altered, but Do Not Correlate with Changes in Brain Topology. Brain Connect. 2020, 10, 411–421. [Google Scholar] [CrossRef]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef]
- Menyhárt, O.; Győrffy, B. Multi-omics approaches in cancer research with applications in tumor subtyping, prognosis, and diagnosis. Comput. Struct. Biotechnol. J. 2021, 19, 949–960. [Google Scholar] [CrossRef]
- Bhinderwala, F.; Wase, N.; DiRusso, C.; Powers, R. Combining Mass Spectrometry and NMR Improves Metabolite Detection and Annotation. J. Proteome Res. 2018, 17, 4017–4022. [Google Scholar] [CrossRef]
- Germeys, C.; Vandoorne, T.; Bercier, V.; Bosch, L.V.D. Existing and Emerging Metabolomic Tools for ALS Research. Genes 2019, 10, 1011. [Google Scholar] [CrossRef]
- Lanznaster, D.; de Assis, D.R.; Corcia, P.; Pradat, P.-F.; Blasco, H. Metabolomics Biomarkers: A Strategy Toward Therapeutics Improvement in ALS. Front. Neurol. 2018, 9, 1126. [Google Scholar] [CrossRef] [Green Version]
- Blasco, H.; Patin, F.; Hounoum, B.M.; Gordon, P.H.; Vourc’H, P.; Andres, C.R.; Corcia, P. Metabolomics in amyotrophic lateral sclerosis: How far can it take us? Eur. J. Neurol. 2016, 23, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.; Larkin, J.R.; Claridge, T.D.W.; Talbot, K.; Sibson, N.R.; Turner, M.R. The longitudinal cerebrospinal fluid metabolomic profile of amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Front. Degener. 2015, 16, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bala, L.; Kalita, J.; Misra, U.; Singh, R.; Khetrapal, C.; Babu, G.N. Metabolomic analysis of serum by (1) H NMR spectroscopy in amyotrophic lateral sclerosis. Clin. Chim. Acta 2010, 411, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wuolikainen, A.; Trupp, M.; Jonsson, P.; Marklund, S.L.; Andersen, P.M.; Forsgren, L.; Öhman, A. NMR analysis of the CSF and plasma metabolome of rigorously matched amyotrophic lateral sclerosis, Parkinson’s disease and control subjects. Metabolomics 2016, 12, 101. [Google Scholar] [CrossRef]
- A Goutman, S.; Boss, J.; Guo, K.; Alakwaa, F.M.; Patterson, A.; Kim, S.; Savelieff, M.G.; Hur, J.; Feldman, E.L. Untargeted metabolomics yields insight into ALS disease mechanisms. J. Neurol. Neurosurg. Psychiatry 2020, 91, 1329–1338. [Google Scholar] [CrossRef]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef]
- Marino, C.; Grimaldi, M.; Sabatini, P.; Amato, P.; Pallavicino, A.; Ricciardelli, C.; D’Ursi, A. Fibromyalgia and Depression in Women: An 1H-NMR Metabolomic Study. Metabolites 2021, 11, 429. [Google Scholar] [CrossRef]
- Castaldo, G.; Pagano, I.; Grimaldi, M.; Marino, C.; Molettieri, P.; Santoro, A.; Stillitano, I.; Romano, R.; Montoro, P.; D’Ursi, A.M.; et al. Effect of Very-Low-Calorie Ketogenic Diet on Psoriasis Patients: A Nuclear Magnetic Resonance-Based Metabolomic Study. J. Proteome Res. 2021, 20, 1509–1521. [Google Scholar] [CrossRef]
- Grimaldi, M.; Palisi, A.; Marino, C.; Montoro, P.; Capasso, A.; Novi, S.; Tecce, M.F.; D’Ursi, A.M. NMR-based metabolomic profile of hypercholesterolemic human sera: Relationship with in vitro gene expression? PLoS ONE 2020, 15, e0231506. [Google Scholar] [CrossRef]
- Ravanbakhsh, S.; Liu, P.; Bjordahl, T.C.; Mandal, R.; Grant, J.R.; Wilson, M.; Eisner, R.; Sinelnikov, I.; Hu, X.; Luchinat, C. Accurate, fully-automated NMR spectral profiling for metabolomics. PLoS ONE 2015, 10, e0124219. [Google Scholar]
- Sommella, E.; Carrizzo, A.; Merciai, F.; Di Sarno, V.; Carbone, D.; De Lucia, M.; Musella, S.; Vecchione, C.; Campiglia, P.s. Analysis of the metabolic switch induced by the spirulina peptide SP6 in high fat diet ApoE-/-mice model: A direct infusion FT-ICR-MS based approach. J. Pharm. Biomed. Anal. 2021, 195, 113865. [Google Scholar] [CrossRef]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, D521–D526. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Metabolomic data processing, analysis, and interpretation using MetaboAnalyst. Curr. Protoc. Bioinform. 2011, 34, 14.10.1–14.10.48. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic Quotient Normalization as Robust Method to Account for Dilution of Complex Biological Mixtures. Application in 1H NMR Metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef] [PubMed]
- Mickiewicz, B.; Vogel, H.J.; Wong, H.R.; Winston, B.W. Metabolomics as a Novel Approach for Early Diagnosis of Pediatric Septic Shock and Its Mortality. Am. J. Respir. Crit. Care Med. 2013, 187, 967–976. [Google Scholar] [CrossRef]
- Wiklund, S.; Johansson, E.; Sjöström, L.; Mellerowicz, E.J.; Edlund, U.; Shockcor, J.P.; Gottfries, J.; Moritz, A.T.; Trygg, J. Visualization of GC/TOF-MS-Based Metabolomics Data for Identification of Biochemically Interesting Compounds Using OPLS Class Models. Anal. Chem. 2008, 80, 115–122. [Google Scholar] [CrossRef]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Vanderlooy, S.; Hüllermeier, E. A critical analysis of variants of the AUC. Mach. Learn. 2008, 72, 247–262. [Google Scholar] [CrossRef] [Green Version]
- Rosset, S. Model selection via the AUC. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; p. 89. [Google Scholar]
- Fabregat, A.; Sidiropoulos, K.; Garapati, P.; Gillespie, M.; Hausmann, K.; Haw, R.; Jassal, B.; Jupe, S.; Korninger, F.; McKay, S.; et al. The Reactome pathway Knowledgebase. Nucleic Acids Res. 2016, 44, D481–D487. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Sun, X.; Xie, H.; Zhang, T.; Zu, X.; Qiao, L.; Zhang, W. Plasma metabolomics coupled with MetaboAnalyst and Ingenuity Pathway Analysis characterizes linoleic acid metabolism disorder in patients with spleen-yang-deficiency syndrome. Eur. J. Integr. Med. 2018, 19, 72–79. [Google Scholar] [CrossRef]
- Grimaldi, M.; Marino, C.; Buonocore, M.; Santoro, A.; Sommella, E.; Merciai, F.; Salviati, E.; De Rosa, A.; Nuzzo, T.; Errico, F.; et al. Prenatal and Early Postnatal Cerebral d-Aspartate Depletion Influences l-Amino Acid Pathways, Bioenergetic processes, and Developmental Brain Metabolism. J. Proteome Res. 2021, 20, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Duren, W.; Evans, C.R.; Burant, C.F.; Michailidis, G.; Karnovsky, A. Sparse network modeling and Metscape-based visualization methods for the analysis of large-scale metabolomics data. Bioinformatics 2017, 33, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hernandez, J.M.; Van Mieghem, P. Betweenness centrality in a weighted network. Phys. Rev. E 2008, 77, 046105. [Google Scholar] [CrossRef] [PubMed]
- Barthelemy, M. Betweenness centrality in large complex networks. Eur. Phys. J. B 2004, 38, 163–168. [Google Scholar] [CrossRef]
- Good, P.J.I. Robustness of Pearson correlation. Interstat 2009, 15, 1–6. [Google Scholar]
- Kellogg, J.J.; Kvalheim, O.M.; Cech, N.B. Composite score analysis for unsupervised comparison and network visualization of metabolomics data. Anal. Chim. Acta 2020, 1095, 38–47. [Google Scholar] [CrossRef]
- Westerhuis, J.A.; Van Velzen, E.J.J.; Hoefsloot, H.C.J.; Smilde, A.K. Multivariate paired data analysis: Multilevel PLSDA versus OPLSDA. Metabolomics 2010, 6, 119–128. [Google Scholar] [CrossRef]
- De Souza, L.P.; Alseekh, S.; Brotman, Y.; Fernie, A.R. Network-based strategies in metabolomics data analysis and interpretation: From molecular networking to biological interpretation. Expert Rev. Proteom. 2020, 17, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Zhang, W.; Fan, Z.; Sun, G.; Likhodi, S.; Randell, E.; Zhai, G. Metabolomics differential correlation network analysis of osteoarthritis. In Proceedings of the Biocomputing 2016: Proceedings of the Pacific Symposium, Kohala Coast, HI, USA, 4–8 January 2016; pp. 120–131. [Google Scholar]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradat, P.-F.; Bruneteau, G.; Gordon, P.; Dupuis, L.; Bonnefont-Rousselot, D.; Simon, D.; Salachas, F.; Corcia, P.; Frochot, V.; Lacorte, J.-M.; et al. Impaired glucose tolerance in patients with amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. 2010, 11, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Harno, K.; Rissanen, A.; Palo, J. Glucose tolerance in amyotrophic lateral sclerosis. Acta Neurol. Scand. 1984, 70, 451–455. [Google Scholar] [CrossRef]
- Masrori, P.; Van Damme, P. Amyotrophic lateral sclerosis: A clinical review. Eur. J. Neurol. 2020, 27, 1918–1929. [Google Scholar] [CrossRef]
- Carrera-Juliá, S.; Moreno, M.L.; Barrios, C.; de la Rubia Ortí, J.E.; Drehmer, E. Antioxidant Alternatives in the Treatment of Amyotrophic Lateral Sclerosis: A Comprehensive Review. Front. Physiol. 2020, 11, 63. [Google Scholar] [CrossRef]
- Ruiz-Perez, D.; Guan, H.; Madhivanan, P.; Mathee, K.; Narasimhan, G. So you think you can PLS-DA? BMC Bioinform. 2020, 21, 2. [Google Scholar] [CrossRef]
- Dupuis, L.; Pradat, P.-F.; Ludolph, A.C.; Loeffler, J.-P. Energy metabolism in amyotrophic lateral sclerosis. Lancet Neurol. 2011, 10, 75–82. [Google Scholar] [CrossRef]
- Nakaya, T.; Maragkakis, M.J.S.r. Amyotrophic Lateral Sclerosis associated FUS mutation shortens mitochondria and induces neurotoxicity. Sci. Rep. 2018, 8, 15575. [Google Scholar] [CrossRef]
- Filho, A.D.B.C.; Pinto, I.F.D.; Dantas, L.S.; Xavier, A.M.; Inague, A.; Faria, R.L.; Medeiros, M.H.G.; Glezer, I.; Yoshinaga, M.Y.; Miyamoto, S. Alterations in lipid metabolism of spinal cord linked to amyotrophic lateral sclerosis. Sci. Rep. 2019, 9, 11642. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, Y.; Zhao, M.; Zheng, L.; Fan, D. Changes in the concentrations of trimethylamine N-oxide (TMAO) and its precursors in patients with amyotrophic lateral sclerosis. Sci. Rep. 2020, 10, 15198. [Google Scholar] [CrossRef] [PubMed]
- Hooten, K.G.; Beers, D.R.; Zhao, W.; Appel, S.H. Protective and Toxic Neuroinflammation in Amyotrophic Lateral Sclerosis. Neurotherapeutics 2015, 12, 364–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasco, H.; Mavel, S.; Corcia, P.; Gordon, P. The Glutamate Hypothesis in ALS: Pathophysiology and Drug Development. Curr. Med. Chem. 2014, 21, 3551–3575. [Google Scholar] [CrossRef]
- Karbowska, M.; Hermanowicz, J.M.; Tankiewicz-Kwedlo, A.; Kalaska, B.; Kaminski, T.W.; Nosek, K.; Wisniewska, R.J.; Pawlak, D. Neurobehavioral effects of uremic toxin–indoxyl sulfate in the rat model. Sci. Rep. 2020, 10, 9483. [Google Scholar] [CrossRef]
- Ferreira, G.K.; Scaini, G.; Carvalho-Silva, M.; Gomes, L.M.; Borges, L.S.; Vieira, J.S.; Constantino, L.S.; Ferreira, G.C.; Schuck, P.F.; Streck, E.L. Effect of l-Tyrosine In Vitro and In Vivo on Energy Metabolism Parameters in Brain and Liver of Young Rats. Neurotox. Res. 2013, 23, 327–335. [Google Scholar] [CrossRef]
- Whitehouse, P.J.; Wamsley, J.K.; Zarbin, M.A.; Price, D.L.; Tourtellotte, W.W.; Kuhar, M.J. Amyotrophic lateral sclerosis: Alterations in neurotransmitter receptors. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 1983, 14, 8–16. [Google Scholar] [CrossRef]
- Heath, P.R.; Shaw, P. Update on the glutamatergic neurotransmitter system and the role of excitotoxicity in amyotrophic lateral sclerosis. Muscle Nerve 2002, 26, 438–458. [Google Scholar] [CrossRef]
- van Kruining, D.; Luo, Q.; van Echten-Deckert, G.; Mielke, M.M.; Bowman, A.; Ellis, S.; Gil Oliveira, T.; Martinez-Martinez, P. Sphingolipids as prognostic biomarkers of neurodegeneration, neuroinflammation, and psychiatric diseases and their emerging role in lipidomic investigation methods. Adv. Drug Deliv. Rev. 2020, 159, 232–244. [Google Scholar] [CrossRef]
- Area-Gomez, E.; Larrea, D.; Yun, T.; Xu, Y.; Hupf, J.; Zandkarimi, F.; Chan, R.B.; Mitsumoto, H. Lipidomics study of plasma from patients suggest that ALS and PLS are part of a continuum of motor neuron disorders. Sci. Rep. 2021, 11, 13562. [Google Scholar] [CrossRef]
- Wheeler, D.; Bandaru, V.V.R.; Calabresi, P.A.; Nath, A.; Haughey, N.J. A defect of sphingolipid metabolism modifies the properties of normal appearing white matter in multiple sclerosis. Brain 2008, 131, 3092–3102. [Google Scholar] [CrossRef]
- Chung, S.Y.; Moriyama, T.; Uezu, E.; Uezu, K.; Hirata, R.; Yohena, N.; Masuda, Y.; Kokubu, T.; Yamamoto, S. Administration of phosphatidylcholine increases brain acetylcholine concentration and improves memory in mice with dementia. J. Nutr. 1995, 125, 1484–1489. [Google Scholar] [PubMed]
- Syme, C.; Pelletier, S.; Shin, J.; Abrahamowicz, M.; Leonard, G.; Perron, M.; Richer, L.; Veillette, S.; Gaudet, D.; Pike, B.; et al. Visceral fat-related systemic inflammation and the adolescent brain: A mediating role of circulating glycerophosphocholines. Int. J. Obes. 2019, 43, 1223–1230. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
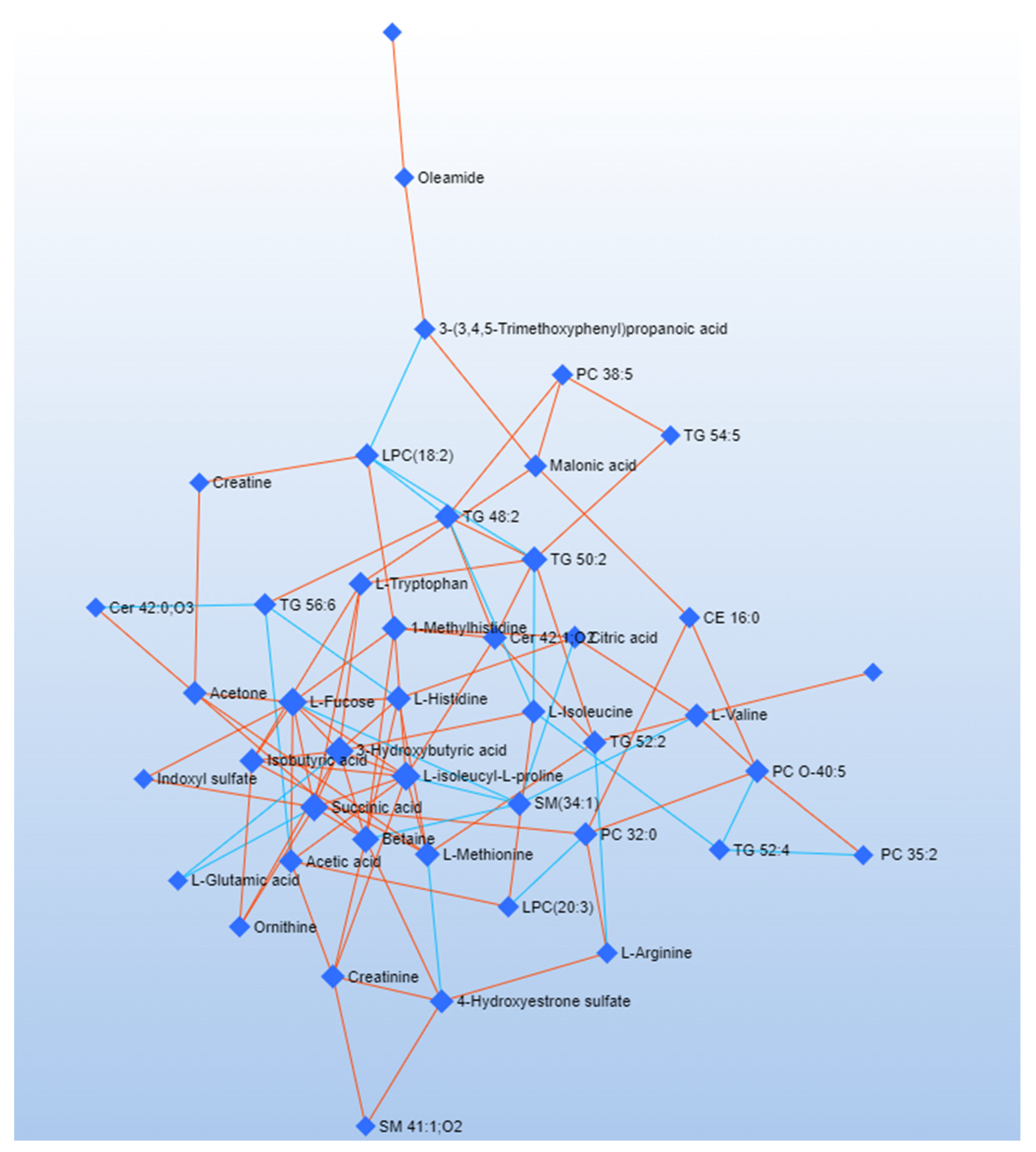{kind=link}
| Parameters | ALS “Advanced” Patients Mean (SD) (n = 6) | ALS “Early” Patients Mean (SD) (n = 9) |
|---|---|---|
| Demographic and clinical measures | ||
| Age | 64.66 (12.20) | 63,92 (10.56) |
| Male/Female | 4/2 | 6/3 |
| Education | 9.17 (4.49) | 8.76 (3.67) |
| Disease duration (months) | 54.33 (43.01) | 38.68 (16. 22) |
| ALSFRS-R score | 32.0 (8.85) | 41.62 (3.60) |
| UMN score | 8.33 (6.77) | 5.34 (2.61) |
| Site of onset | 1 bulbar | 1 bulbar |
| 5 Spinal | 8 Spinal | |
| Phenotype | 2 predominant LMN. | 7 predominant LMN. |
| 1 predominant U.M.N. | 1 predominant U.M.N. | |
| 2 Classic | 1 Classic | |
| Riluzole | 6/6 | 9/9 |
| Neuropsychological parameters | ||
| ECAS test (total score) | 93.67 (14.14) | 93.45 (14.82) |
| Network Analysis Related to Early Patients | ||
|---|---|---|
| Label | Degree | Betweenness |
| L-Histidine | 16 | 59.91 |
| TG 54:5 | 15 | 58.3 |
| L-Glutamine | 15 | 32.01 |
| Acetic acid | 14 | 45.95 |
| CE 16:0 | 14 | 40.1 |
| Cer 42:1;O2 | 13 | 33.92 |
| PE O-38:5 | 13 | 31.09 |
| L-Valine | 13 | 27.77 |
| Malonic acid | 13 | 23.08 |
| L-Fucose | 12 | 51.26 |
| SM 41:1;O2 | 12 | 32.34 |
| 1-Methylhistidine | 12 | 24.22 |
| L-Arginine | 12 | 20.99 |
| L-Methionine | 12 | 18.84 |
| Betaine | 11 | 38.48 |
| TG 52:2 | 11 | 25.04 |
| TG 56:6 | 11 | 24.16 |
| Linoleamide | 11 | 20.78 |
| Oleamide | 11 | 20.36 |
| 3-(3,4,5-Trimethoxyphenyl)propanoic acid | 10 | 32.28 |
| Cer 42:0;O3 | 10 | 19.77 |
| TG 48:2 | 10 | 19.28 |
| L-isoleucyl-L-proline | 10 | 15.43 |
| TG 52:4 | 10 | 13.13 |
| L-Glutamic acid | 10 | 10.19 |
| SM(34:1) | 10 | 4.89 |
| Ornithine | 9 | 15.13 |
| Acetone | 9 | 14.81 |
| PC 35:2 | 9 | 11.77 |
| 3-Hydroxybutyric acid | 9 | 7.81 |
| Succinic acid | 9 | 6.81 |
| LPC(20:3) | 8 | 31.8 |
| Creatinine | 8 | 13.73 |
| Creatine | 8 | 10.52 |
| Isobutyric acid | 8 | 7.92 |
| PC 32:0 | 8 | 3.67 |
| PC O-40:5 | 7 | 8.09 |
| PC O-38:5 | 7 | 6.29 |
| TG 50:2 | 7 | 2.66 |
| L-Tryptophan | 6 | 20.66 |
| Citric acid | 5 | 9.23 |
| Indoxyl sulfate | 4 | 6.16 |
| LPC(18:2) | 3 | 3.01 |
| L-Isoleucine | 3 | 0.35 |
| 4-Hydroxyestrone sulfate | 2 | 0 |
| Network analysis related to advanced patients | ||
| Label | Degree | Betweenness |
| 3-Hydroxybutyric acid | 10 | 88.58 |
| L-Isoleucyl-L-proline | 10 | 75.85 |
| L-Fucose | 10 | 70.7 |
| Succinic acid | 9 | 89.14 |
| Betaine | 8 | 74.67 |
| TG 50:2 | 7 | 83.66 |
| 1-Methylhistidine | 6 | 65.9 |
| TG 48:2 | 6 | 64.41 |
| L-Methionine | 6 | 37.13 |
| L-Histidine | 6 | 32.1 |
| Isobutyric acid | 6 | 19.31 |
| LPC(18:2) | 5 | 100.12 |
| L-Valine | 5 | 75.3 |
| L-Isoleucine | 5 | 72.02 |
| PC 32:0 | 5 | 65.74 |
| L-Tryptophan | 5 | 62.42 |
| TG 52:2 | 5 | 58.58 |
| PC O-40:5 | 5 | 54.62 |
| Acetone | 5 | 45.33 |
| SM(34:1) | 5 | 33.5 |
| Acetic acid | 5 | 29.88 |
| 4-Hydroxyestrone sulfate | 5 | 29.83 |
| Cer 42:1 | 5 | 28.68463 |
| Creatinine | 5 | 27.83 |
| Malonic acid | 4 | 65.18 |
| TG 56:6 | 4 | 33.45 |
| Citric acid | 4 | 18.85 |
| 3-(3-45-Trimethoxyphenyl-propanoic acid) | 3 | 88.1 |
| CE 16:0 | 3 | 28.59 |
| TG 52:4 | 3 | 21.78 |
| L-Arginine | 3 | 19.99 |
| LPC(20:3) | 3 | 15.73 |
| PC 38:5 | 3 | 11.94 |
| Ornithine | 3 | 0.5 |
| Oleamide | 2 | 42 |
| Creatine | 2 | 9.23 |
| Pathway | Software | p-Value | FDR. |
|---|---|---|---|
| Ammonia Recycling | Metaboanalyst | 7.27 × 10−10 | 0.000538 |
| Mitochondrial Beta-Oxidation of Long-Chain Saturated Fatty Acids | Metaboanalyst | 0.0125 | 0.056 |
| Carnitine Synthesis | Metaboanalyst | 0.0125 | 0.0564 |
| Beta Oxidation of Very Long Chain Fatty Acids | Metaboanalyst | 0.015 | 0.0564 |
| Oxidation of Branched Chain Fatty Acids | Metaboanalyst | 0.015 | 0.0564 |
| Amine ligand-binding receptors | Reactome | 1.30 × 10−3 | 1.39 × 10−6 |
| Serotonin receptors | Reactome | 1.10 × 10−5 | 5.89 × 10−6 |
| Defective SLC6A19 causes Hartnup disorder (HND) | Reactome | 3.80 × 10−10 | 0.008 |
| Na−/Cl− dependent neurotransmitter transporters | Reactome | 5.13 × 10−10 | 0.009 |
| Cytosolic tRNA aminoacylation | Reactome | 8.64 × 10−9 | 0.011 |
| Class A/1 (Rhodopsin-like receptors) | Reactome | 9.88 × 10−10 | 0.011 |
| Adrenoceptors | Reactome | 1.26 × 10−12 | 0.013 |
| Muscarinic acetylcholine receptors | Reactome | 1.47 × 10−11 | 0.014 |
| GPCR ligand binding | Reactome | 5.59 × 10−11 | 0.048 |
| Chemokine receptors bind chemokines | Reactome | 5.88 × 10−11 | 0.048 |
| Adrenaline signaling through Alpha-2 adrenergic receptor | Reactome | 0.001 | 0.105 |
| Adenylate cyclase inhibitory pathway | Reactome | 0.007 | 0.492 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marino, C.; Grimaldi, M.; Sommella, E.M.; Ciaglia, T.; Santoro, A.; Buonocore, M.; Salviati, E.; Trojsi, F.; Polverino, A.; Sorrentino, P.; et al. The Metabolomic Profile in Amyotrophic Lateral Sclerosis Changes According to the Progression of the Disease: An Exploratory Study. Metabolites 2022, 12, 837. https://doi.org/10.3390/metabo12090837
Marino C, Grimaldi M, Sommella EM, Ciaglia T, Santoro A, Buonocore M, Salviati E, Trojsi F, Polverino A, Sorrentino P, et al. The Metabolomic Profile in Amyotrophic Lateral Sclerosis Changes According to the Progression of the Disease: An Exploratory Study. Metabolites. 2022; 12(9):837. https://doi.org/10.3390/metabo12090837
Chicago/Turabian StyleMarino, Carmen, Manuela Grimaldi, Eduardo Maria Sommella, Tania Ciaglia, Angelo Santoro, Michela Buonocore, Emanuela Salviati, Francesca Trojsi, Arianna Polverino, Pierpaolo Sorrentino, and et al. 2022. "The Metabolomic Profile in Amyotrophic Lateral Sclerosis Changes According to the Progression of the Disease: An Exploratory Study" Metabolites 12, no. 9: 837. https://doi.org/10.3390/metabo12090837