
A Multi-Matrix Metabolomic Approach in Ringed Seals and Beluga Whales to Evaluate Contaminant and Climate-Related Stressors
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Biological Variables
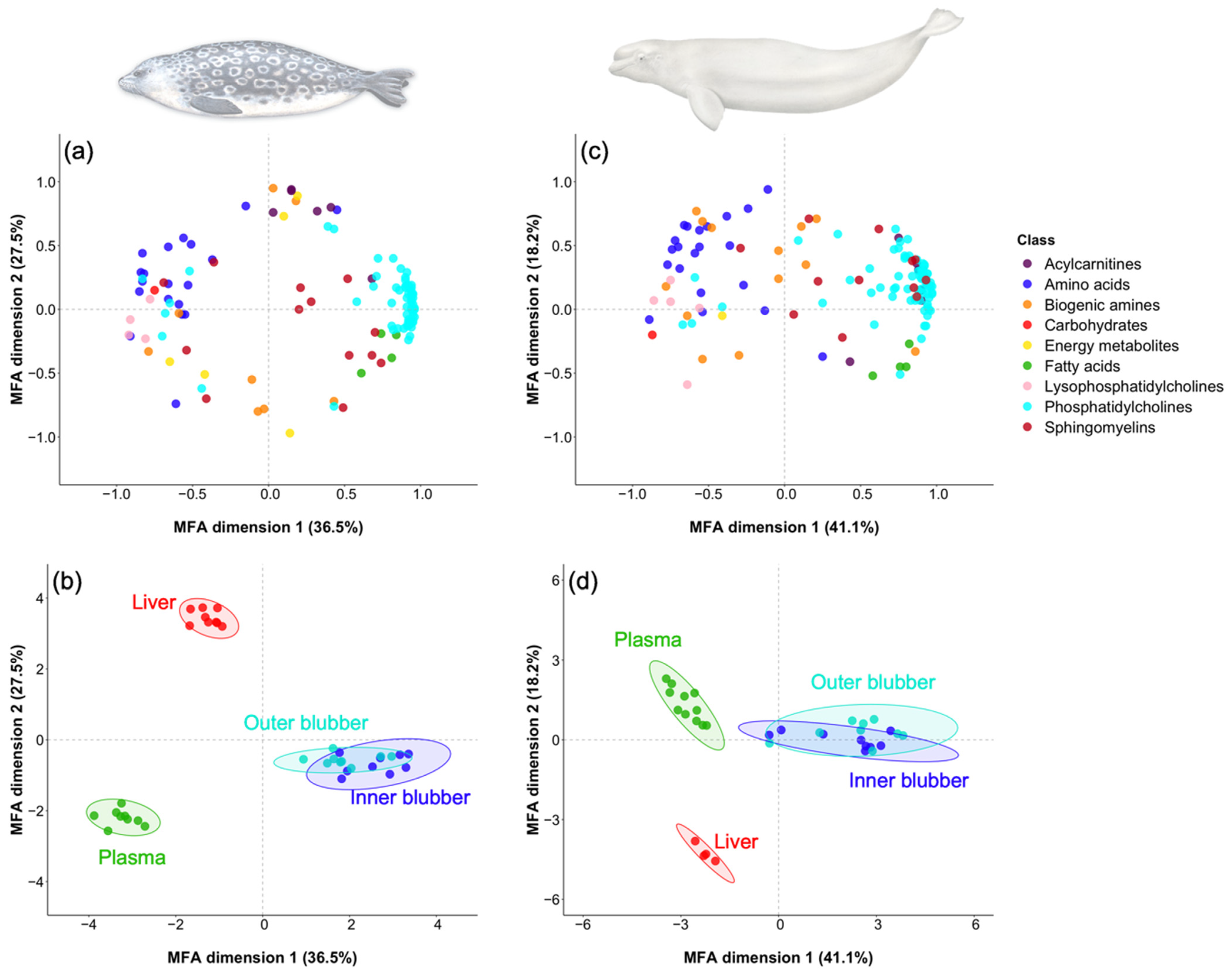
2.2. Metabolites
2.2.1. Metabolite Profiles
2.2.2. Metabolite Concentrations
2.3. Contaminants
2.4. Correlations between Biological and Contaminant Variables and Metabolite Percent Contributions
3. Discussion
3.1. Inter-Tissue and Inter-Species Metabolite Profiles
3.2. Contaminants and Effect Thresholds
3.3. Correlations between Metabolite Profiles, Biological Variables and Contaminants
3.4. Tissue Selection Guidance for Marine Mammal Health Monitoring
4. Materials and Methods
4.1. Field Sampling
4.2. Metabolomic Analysis
4.3. Chemical Analyses
4.3.1. Organohalogens
4.3.2. Total Mercury
4.4. Stable Isotope Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reijnders, P.J.H.; Borrell, A.; van Franeker, J.A.; Aguilar, À. Pollution. In Encyclopedia of Marine Mammals, 3rd ed.; Würsig, B., Thewissen, J.G.M., Kovacs, K.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 746–753. ISBN 978-0-12-804327-1. [Google Scholar]
- Houde, M.; Hoekstra, P.F.; Solomon, K.R.; Muir, D.C.G. Organohalogen contaminants in delphinoid cetaceans. Rev. Environ. Contam. Toxicol. 2005, 184, 1–57. [Google Scholar] [CrossRef] [PubMed]
- de Wit, C.A.; Herzke, D.; Vorkamp, K. Brominated flame retardants in the Arctic environment—Trends and new candidates. Sci. Total Environ. 2010, 408, 2885–2918. [Google Scholar] [CrossRef] [PubMed]
- Simond, A.E.; Houde, M.; Lesage, V.; Michaud, R.; Verreault, J. Metabolomic profiles of the endangered St. Lawrence Estuary beluga population and associations with organohalogen contaminants. Sci. Total Environ. 2020, 717, 137204. [Google Scholar] [CrossRef] [PubMed]
- Lebeuf, M.; Measures, L.; Noël, M.; Raach, M.; Trottier, S. A twenty-one year temporal trend of persistent organic pollutants in St. Lawrence Estuary beluga, Canada. Sci. Total Environ. 2014, 485, 377–386. [Google Scholar] [CrossRef]
- Kershaw, J.L.; Hall, A.J. Mercury in cetaceans: Exposure, bioaccumulation and toxicity. Sci. Total Environ. 2019, 694, 133683. [Google Scholar] [CrossRef]
- Reijnders, P.J.H.; Simmonds, M. Global temporal trends of organochlorines and heavy metals in pinnipeds. In Toxicology of Marine Mammals; Vos, J.G., Bossart, G.D., Fournier, M., O’Shea, T.J., Eds.; CRC Press: London, UK; New York, NY, USA, 2002; pp. 490–506. ISBN 0203165578. [Google Scholar]
- Brown, T.M.; Ross, P.S.; Reimer, K.J.; Veldhoen, N.; Dangerfield, N.J.; Fisk, A.T.; Helbing, C.C. PCB related effects thresholds as derived through gene transcript profiles in locally contaminated ringed seals (Pusa hispida). Environ. Sci. Technol. 2014, 48, 12952–12961. [Google Scholar] [CrossRef]
- Simond, A.E.; Houde, M.; Lesage, V.; Michaud, R.; Zbinden, D.; Verreault, J. Associations between organohalogen exposure and thyroid- and steroid-related gene responses in St. Lawrence Estuary belugas and minke whales. Mar. Pollut. Bull. 2019, 145, 174–184. [Google Scholar] [CrossRef]
- Routti, H.; Arukwe, A.; Jenssen, B.M.; Letcher, R.J.; Nyman, M.; Bäckman, C.; Gabrielsen, G.W. Comparative endocrine disruptive effects of contaminants in ringed seals (Phoca hispida) from Svalbard and the Baltic Sea. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 152, 306–312. [Google Scholar] [CrossRef]
- Troisi, G.M.; Barton, S.J.; Liori, O.; Nyman, M. Polychlorinated Biphenyls (PCBs) and Sex Hormone Concentrations in Ringed and Grey Seals: A Possible Link to Endocrine Disruption? Arch. Environ. Contam. Toxicol. 2020, 78, 513–524. [Google Scholar] [CrossRef]
- Clarkson, T.W.; Magos, L. The toxicology of mercury and its chemical compounds. Crit. Rev. Toxicol. 2006, 36, 609–662. [Google Scholar] [CrossRef]
- Tan, S.W.; Meiller, J.C.; Mahaffey, K.R. The endocrine effects of mercury in humans and wildlife. Crit. Rev. Toxicol. 2009, 39, 228–269. [Google Scholar] [CrossRef] [PubMed]
- Desforges, J.-P.W.; Ross, P.S.; Loseto, L.L. Transplacental transfer of polychlorinated biphenyls and polybrominated diphenyl ethers in arctic beluga whales (Delphinapterus leucas). Environ. Toxicol. Chem. 2012, 31, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Lebeuf, M.; Raach, M.; Measures, L.; Ménard, N.; Hammill, M.O. Temporal Trends of PBDEs in Adult and Newborn Beluga (Delphinapterus leucas) from the St. Lawrence Estuary; Canadian Science Advisory Secretariat: Ottawa, ON, Canada, 2014. [Google Scholar]
- Cullon, D.L. Biomagnification and Fate of Persistent Organic Pollutants (POPs) in Marine Mammal Food Webs in the Northeastern Pacific Ocean. Ph.D. Thesis, University of Victoria, Victoria, BC, Canada, 2010. [Google Scholar]
- Brown, T.M.; Ross, P.S.; Reimer, K.J. Transplacental Transfer of Polychlorinated Biphenyls, Polybrominated Diphenylethers, and Organochlorine Pesticides in Ringed Seals (Pusa hispida). Arch. Environ. Contam. Toxicol. 2016, 70, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Rea, L.D.; Castellini, J.M.; Correa, L.; Fadely, B.S.; O’Hara, T.M. Maternal Steller sea lion diets elevate fetal mercury concentrations in an area of population decline. Sci. Total Environ. 2013, 454–455, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Habran, S.; Pomeroy, P.P.; Debier, C.; Das, K. Changes in trace elements during lactation in a marine top predator, the grey seal. Aquat. Toxicol. 2013, 126, 455–466. [Google Scholar] [CrossRef]
- Endo, T.; Kimura, O.; Hisamichi, Y.; Minoshima, Y.; Haraguchi, K.; Kakumoto, C.; Kobayashi, M. Distribution of total mercury, methyl mercury and selenium in pod of killer whales (Orcinus orca) stranded in the northern area of Japan: Comparison of mature females with calves. Environ. Pollut. 2006, 144, 145–150. [Google Scholar] [CrossRef]
- Endo, T.; Kimura, O.; Hisamichi, Y.; Minoshima, Y.; Haraguchi, K. Age-dependent accumulation of heavy metals in a pod of killer whales (Orcinus orca) stranded in the northern area of Japan. Chemosphere 2007, 67, 51–59. [Google Scholar] [CrossRef]
- Vorkamp, K.; Balmer, J.; Hung, H.; Letcher, R.J.; Rigét, F.F. A review of chlorinated paraffin contamination in Arctic ecosystems. Emerg. Contam. 2019, 5, 219–231. [Google Scholar] [CrossRef]
- Law, R.J. An overview of time trends in organic contaminant concentrations in marine mammals: Going up or down? Mar. Pollut. Bull. 2014, 82, 7–10. [Google Scholar] [CrossRef]
- Zeng, L.; Lam, J.C.W.; Wang, Y.; Jiang, G.; Lam, P.K.S. Temporal Trends and Pattern Changes of Short- and Medium-Chain Chlorinated Paraffins in Marine Mammals from the South China Sea over the Past Decade. Environ. Sci. Technol. 2015, 49, 11348–11355. [Google Scholar] [CrossRef]
- Brown, T.M.; Macdonald, R.W.; Muir, D.C.G.; Letcher, R.J. The distribution and trends of persistent organic pollutants and mercury in marine mammals from Canada’s Eastern Arctic. Sci. Total Environ. 2018, 618, 500–517. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, T.J.; Tanabe, S. Persistent ocean contaminants and marine mammals: A retrospective overview. In World; Vos, J.G., Bossart, G.D., Fournier, M., O’Shea, T.J., Eds.; CRC Press: London, UK; New York, NY, USA, 2002; pp. 98–134. ISBN 0203165578. [Google Scholar]
- Krey, A.; Ostertag, S.K.; Chan, H.M. Assessment of neurotoxic effects of mercury in beluga whales (Delphinapterus leucas), ringed seals (Pusa hispida), and polar bears (Ursus maritimus) from the Canadian Arctic. Sci. Total Environ. 2015, 509–510, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Noël, M.; Loseto, L.L.; Stern, G.A.; Nöel, M.; Loseto, L.L.; Stern, G.A. Legacy contaminants in the Eastern Beaufort Sea beluga whales (Delphinapterus leucas): Are temporal trends reflecting regulations? Arct. Sci. 2018, 4, 373–387. [Google Scholar] [CrossRef]
- Desforges, J.-P.W.; Ross, P.S.; Dangerfield, N.; Palace, V.P.; Whiticar, M.; Loseto, L.L. Vitamin A and E profiles as biomarkers of PCB exposure in beluga whales (Delphinapterus leucas) from the western Canadian Arctic. Aquat. Toxicol. 2013, 142–143, 317–328. [Google Scholar] [CrossRef]
- Noël, M.; Loseto, L.L.; Helbing, C.C.; Veldhoen, N.; Dangerfield, N.J.; Ross, P.S. PCBs Are Associated With Altered Gene Transcript Profiles in Arctic Beluga Whales (Delphinapterus leucas). Environ. Sci. Technol. 2014, 48, 2942–2951. [Google Scholar] [CrossRef]
- Wang, F.; Outridge, P.M.; Feng, X.; Meng, B.; Heimbürger-Boavida, L.E.; Mason, R.P. How closely do mercury trends in fish and other aquatic wildlife track those in the atmosphere?-Implications for evaluating the effectiveness of the Minamata Convention. Sci. Total Environ. 2019, 674, 58–70. [Google Scholar] [CrossRef]
- Alava, J.J.; Cheung, W.W.L.; Ross, P.S.; Sumaila, U.R. Climate change-contaminant interactions in marine food webs: Toward a conceptual framework. Glob. Chang. Biol. 2017, 23, 3984–4001. [Google Scholar] [CrossRef]
- McKinney, M.A.; Pedro, S.; Dietz, R.; Sonne, C.; Fisk, A.T.; Roy, D.; Jenssen, B.M.; Letcher, R.J. A review of ecological impacts of global climate change on persistent organic pollutant and mercury pathways and exposures in arctic marine ecosystems. Curr. Zool. 2015, 61, 617–628. [Google Scholar] [CrossRef]
- Steiner, N.S.; Bowman, J.; Campbell, K.; Chierici, M.; Eronen-Rasimus, E.; Falardeau, M.; Flores, H.; Fransson, A.; Herr, H.; Insley, S.J.; et al. Climate change impacts on sea-ice ecosystems and associated ecosystem services. Elementa 2021, 9, 7. [Google Scholar] [CrossRef]
- Wassmann, P.; Duarte, C.M.; Agustí, S.; Sejr, M.K. Footprints of climate change in the Arctic marine ecosystem. Glob. Chang. Biol. 2011, 17, 1235–1249. [Google Scholar] [CrossRef]
- Tynan, C.T.; Demaster, D.P. Observations and Predictions of Arctic Climatic Change: Potential Effects on Marine Mammals. ARCTIC 1997, 50, 308–322. [Google Scholar] [CrossRef]
- Kovacs, K.M.; Lydersen, C.; Overland, J.E.; Moore, S.E. Impacts of changing sea-ice conditions on Arctic marine mammals. Mar. Biodivers. 2011, 41, 181–194. [Google Scholar] [CrossRef]
- Species at Risk Public Registry Ringed Seal (Pusa hispida). Available online: https://species-registry.canada.ca/index-en.html#/species/347-1059 (accessed on 11 September 2021).
- Ferguson, S.H.; Stirling, I.; McLoughlin, P. Climate change and ringed seal (Phoca hispida) recruitment in western Hudson Bay. Mar. Mammal Sci. 2005, 21, 121–135. [Google Scholar] [CrossRef]
- Harwood, L.A.; Smith, T.G.; Melling, H. Variation in reproduction and body condition of the ringed seal (Phoca hispida) in western Prince Albert Sound, NT, Canada, as assessed through a harvest-based sampling program. Arctic 2000, 53, 422–431. [Google Scholar] [CrossRef]
- Loseto, L.L.; Stern, G.A.; Macdonald, R.W. Distant drivers or local signals: Where do mercury trends in western Arctic belugas originate? Sci. Total Environ. 2015, 509–510, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Choy, E.S.; Giraldo, C.; Rosenberg, B.; Roth, J.D.; Ehrman, A.D.; Majewski, A.; Swanson, H.; Power, M.; Reist, J.D.; Loseto, L.L. Variation in the diet of beluga whales in response to changes in prey availability: Insights on changes in the Beaufort Sea ecosystem. Mar. Ecol. Prog. Ser. 2020, 647, 195–210. [Google Scholar] [CrossRef]
- Harwood, L.A.; Iacozza, J.; Auld, J.C.; Norton, P.; Loseto, L.L. Belugas in the Mackenzie River estuary, NT, Canada: Habitat use and hot spots in the Tarium Niryutait Marine Protected Area. Ocean Coast. Manag. 2014, 100, 128–138. [Google Scholar] [CrossRef]
- Loseto, L.L.; Hoover, C.; Ostertag, S.K.; Whalen, D.; Pearce, T.; Paulic, J.; Iacozza, J.; MacPhee, S. Beluga whales (Delphinapterus leucas), environmental change and marine protected areas in the Western Canadian Arctic. Estuar. Coast. Shelf Sci. 2018, 212, 128–137. [Google Scholar] [CrossRef]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2009, 5, 3. [Google Scholar] [CrossRef]
- Tartu, S.; Lille-Langoy, R.; Størseth, T.R.; Bourgeon, S.; Brunsvik, A.; Aars, J.; Goksøyr, A.; Jenssen, B.M.; Polder, A.; Thiemann, G.W.; et al. Multiple-stressor effects in an apex predator: Combined influence of pollutants and sea ice decline on lipid metabolism in polar bears. Sci. Rep. 2017, 7, 16487. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.M.; Ekman, D.R.; Teng, Q.; Ankley, G.T.; Berninger, J.P.; Cavallin, J.E.; Jensen, K.M.; Kahl, M.D.; Schroeder, A.L.; Villeneuve, D.L.; et al. Linking field-based metabolomics and chemical analyses to prioritize contaminants of emerging concern in the Great Lakes basin. Environ. Toxicol. Chem. 2016, 35, 2493–2502. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.Y.; Benskin, J.P.; Veldhoen, N.; Chandramouli, B.; Butler, H.; Helbing, C.C.; Cosgrove, J.R. A multi-omic approach to elucidate low-dose effects of xenobiotics in zebrafish (Danio rerio) larvae. Aquat. Toxicol. 2017, 182, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.M.; Hammond, S.A.; Behsaz, B.; Veldhoen, N.; Birol, I.; Helbing, C.C. De novo assembly of the ringed seal (Pusa hispida) blubber transcriptome: A tool that enables identification of molecular health indicators associated with PCB exposure. Aquat. Toxicol. 2017, 185, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Kovacevic, V.; Simpson, M.J. Fundamentals of Environmental Metabolomics; Elsevier Inc.: Amsterdam, The Netherlands, 2020; ISBN 9780128181966. [Google Scholar]
- Morris, A.D.; Letcher, R.J.; Dyck, M.; Chandramouli, B.; Cosgrove, J. Concentrations of legacy and new contaminants are related to metabolite profiles in Hudson Bay polar bears. Environ. Res. 2019, 168, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Green, C.J.; Hodson, L. The influence of dietary fat on liver fat accumulation. Nutrients 2014, 6, 5018–5033. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Von Ruesten, A.; Drogan, D.; Schulze, M.B.; Prehn, C.; Adamski, J.; Pischon, T.; Boeing, H. Variation of serum metabolites related to habitual diet: A targeted metabolomic approach in EPIC-potsdam. Eur. J. Clin. Nutr. 2013, 67, 1100–1108. [Google Scholar] [CrossRef]
- Ma, S.; Yim, S.H.; Lee, S.G.; Kim, E.B.; Lee, S.R.; Chang, K.T.; Buffenstein, R.; Lewis, K.N.; Park, T.J.; Miller, R.A.; et al. Organization of the Mammalian Metabolome according to Organ Function, Lineage Specialization, and Longevity. Cell Metab. 2015, 22, 332–343. [Google Scholar] [CrossRef] [PubMed]
- ten Hove, M.; Pater, L.; Storm, G.; Weiskirchen, S.; Weiskirchen, R.; Lammers, T.; Bansal, R. The hepatic lipidome: From basic science to clinical translation. Adv. Drug Deliv. Rev. 2020, 159, 180–197. [Google Scholar] [CrossRef]
- Davis, R.W. Marine Mammals-Adaptations for an Aquatic Life; Springer International Publishing: Cham, Switzerland, 2019; ISBN 978-3-319-98278-6. [Google Scholar]
- Hayamizu, K. Amino Acids and Energy Metabolism: An Overview. In Sustained Energy for Enhanced Human Functions and Activity; Elsevier: Amsterdam, The Netherlands, 2017; pp. 339–349. ISBN 9780128093320. [Google Scholar]
- Brown, T.M.; Iverson, S.J.; Fisk, A.T.; Macdonald, R.W.; Helbing, C.C.; Reimer, K.J. Local contamination, and not feeding preferences, explains elevated PCB concentrations in Labrador ringed seals (Pusa hispida). Sci. Total Environ. 2015, 515–516, 188–197. [Google Scholar] [CrossRef]
- Xiong, D. Bioaccumulation and Biomagnification of Perfluoroalkyl Substances (PFAS) in a Subarctic Ringed Seal Food Web in Lake Melville, Northern Labrador, Canada. Ph.D. Thesis, University of Toronto, Toronto, ON, Canada, 2021. [Google Scholar]
- Strandberg, U.; Käkelä, A.; Lydersen, C.; Kovacs, K.M.; Grahl-Nielsen, O.; Hyvärinen, H.; Käkelä, R. Stratification, Composition, and Function of Marine Mammal Blubber: The Ecology of Fatty Acids in Marine Mammals. Physiol. Biochem. Zool. 2008, 81, 473–485. [Google Scholar] [CrossRef]
- Iverson, S.J.; Koopman, H.N. Blubber. In Encyclopedia of Marine Mammals, 3rd ed.; Würsig, B., Thewissen, J.G.M., Kovacs, K.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 107–110. ISBN 978-0-12-804327-1. [Google Scholar]
- Costa, D.P.; Maresh, J.L. Energetics. In Encyclopedia of Marine Mammals; Elsevier: Amsterdam, The Netherlands, 2018; pp. 329–335. [Google Scholar]
- Smith, H.R. Fatty Acid Variation in Beluga (Delphinapterus leucas) blubber: Implications for Estimating Diet Using Fatty Acids. Ph.D. Thesis, University of Washington, Seattle, WA, USA, 2009. [Google Scholar]
- Casares, D.; Escrib, P.V.; Casares, D.; Escribá, P.V.; Rosselló, C.A. Membrane Lipid Composition Effect on Membrane and Organelle Structure, Function and Compartmentalization and Therapeutic Avenues. Int. J. Mol. Sci. 2019, 20, 2167. [Google Scholar] [CrossRef] [PubMed]
- Storrie, L.; Hussey, N.E.; MacPhee, S.A.; O’Corry-Crowe, G.; Iacozza, J.; Barber, D.G.; Nunes, A.; Loseto, L.L. Year-Round Dive Characteristics of Male Beluga Whales From the Eastern Beaufort Sea Population Indicate Seasonal Shifts in Foraging Strategies. Front. Mar. Sci. 2022, 8, 1–22. [Google Scholar] [CrossRef]
- Hammill, M.O. Ringed Seal. In Encyclopedia of Marine Mammals; Elsevier: Amsterdam, The Netherlands, 2009; pp. 972–974. [Google Scholar]
- Ryg, M.; Smith, T.G.; Øritsland, N.A. Seasonal changes in body mass and body composition of ringed seals (Phoca hispida) on Svalbard. Can. J. Zool. 1990, 68, 470–475. [Google Scholar] [CrossRef]
- Bhagavan, N.V.; Ha, C.-E. Carbohydrate Metabolism II. In Essentials of Medical Biochemistry; Elsevier: Amsterdam, The Netherlands, 2015; pp. 205–225. ISBN 9780124166875. [Google Scholar]
- Lauderdale, L.K.; Walsh, M.T.; Mitchell, K.A.; Granger, D.A.; Mellen, J.D.; Miller, L.J. Health reference intervals and values for common bottlenose dolphins (Tursiops truncatus), Indo-Pacific bottlenose dolphins (Tursiops aduncus), Pacific white-sided dolphins (Lagenorhynchus obliquidens), and beluga whales (Delphinapterus leucas). PLoS ONE 2021, 16, e0250332. [Google Scholar] [CrossRef]
- Norman, S.A.; Goertz, C.E.C.; Burek, K.A.; Quakenbush, L.T.; Cornick, L.A.; Romano, T.A.; Spoon, T.; Miller, W.; Beckett, L.A.; Hobbs, R.C. Seasonal hematology and serum chemistry of wild beluga whales (Delphinapterus leucas) in Bristol Bay, Alaska, USA. J. Wildl. Dis. 2012, 48, 21–32. [Google Scholar] [CrossRef]
- Facey, A.; Irving, R.; Dilworth, L. Overview of Lactate Metabolism and the Implications for Athletes. Am. J. Sport. Sci. Med. 2013, 1, 42–46. [Google Scholar] [CrossRef]
- Duncan, W.P.; Júnior, J.N.A.; Mendonça, W.C.S.; Santa Cruz, I.F.; Samonek, J.F.; Morais, E.J.F.; Marcon, J.L.; Da Silveira, R. Physiological stress response in free-living Amazonian caimans following experimental capture. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2021, 337, 282–292. [Google Scholar] [CrossRef]
- Fuller, L.; Stell, E.; Leary, C.; Parsons, G. Circulating adrenocorticotropic hormone levels, lactate levels, hematocrit and osmolality in relation to capture stress in Atlantic sharpnose sharks, Rhizoprionodon terraenovae. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2020, 243, 110655. [Google Scholar] [CrossRef]
- Lanyon, J.M.; Wong, A.; Long, T.; Woolford, L. Serum biochemistry reference intervals of live wild dugongs (Dugong dugon) from urban coastal Australia. Vet. Clin. Pathol. 2015, 44, 234–242. [Google Scholar] [CrossRef]
- St. Aubin, D.J.; Forney, K.A.; Chivers, S.J.; Scott, M.D.; Danil, K.; Romano, T.A.; Wells, R.S.; Gulland, F.M.D. Hematological, serum, and plasma chemical constituents in pantropical spotted dolphins (Stenella attenuata) following chase, encirclement, and tagging. Mar. Mammal Sci. 2013, 29, 14–35. [Google Scholar] [CrossRef]
- Petricorena, Z.C. Chemical Composition of Fish and Fishery Products Proteins in Fish. In Handbook of Food Chemistry; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–28. ISBN 9783642416095. [Google Scholar]
- Wu, J.Y.; Prentice, H. Role of taurine in the central nervous system. J. Biomed. Sci. 2010, 17, S1. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.T.; Rodgers, Q.R. Taurine Levels in Cetaceans: A Preliminary Investigation. In Proceedings of the International Association for Aquatic Mammal Medicine, Orlando, FL, USA; 1995. [Google Scholar]
- Simond, A.E.; Houde, M.; Lesage, V.; Verreault, J. Temporal trends of PBDEs and emerging flame retardants in belugas from the St. Lawrence Estuary (Canada) and comparisons with minke whales and Canadian Arctic belugas. Environ. Res. 2017, 156, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Houde, M.; Wang, X.; Colson, T.L.L.; Gagnon, P.; Ferguson, S.H.; Ikonomou, M.G.; Dubetz, C.; Addison, R.F.; Muir, D.C.G. Trends of persistent organic pollutants in ringed seals (Phoca hispida) from the Canadian Arctic. Sci. Total Environ. 2019, 665, 1135–1146. [Google Scholar] [CrossRef]
- Houde, M.; Wang, X.; Ferguson, S.H.; Gagnon, P.; Brown, T.M.; Tanabe, S.; Kunito, T.; Kwan, M.; Muir, D.C.G. Spatial and temporal trends of alternative flame retardants and polybrominated diphenyl ethers in ringed seals (Phoca hispida) across the Canadian Arctic. Environ. Pollut. 2017, 223, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Frouin, H.; Lebeuf, M.; Hammill, M.O.; Sjare, B.; Fournier, M. PBDEs in serum and blubber of harbor, grey and harp seal pups from Eastern Canada. Chemosphere 2011, 82, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.S.; Noël, M.; Lambourn, D.; Dangerfield, N.; Calambokidis, J.; Jeffries, S. Declining concentrations of persistent PCBs, PBDEs, PCDEs, and PCNs in harbor seals (Phoca vitulina) from the Salish Sea. Prog. Oceanogr. 2013, 115, 160–170. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, L.; Zhou, F.; Shaw, S.; Roos, A.; Berger, M.; Bäcklin, B.M.; Huang, Y.; Zheng, X.; Wang, X.; et al. Hepatic Fatty Acid Profiles Associated with Exposure to Emerging and Legacy Halogenated Contaminants in Two Harbor Seal Populations across the North Atlantic. Environ. Sci. Technol. 2022, 56, 1830–1840. [Google Scholar] [CrossRef]
- Vorkamp, K.; Rigét, F.F. A review of new and current-use contaminants in the Arctic environment: Evidence of long-range transport and indications of bioaccumulation. Chemosphere 2014, 111, 379–395. [Google Scholar] [CrossRef]
- de Wit, C.A.; Kierkegaard, A.; Ricklund, N.; Sellström, U. Emerging Brominated Flame Retardants in the environment. In Brominated Flame Retardants; Eljarrat, E., Barceló, D., Eds.; Springer: Berlin/Heidelberg, Germany; London, UK; New York, NY, USA, 2011; Volume 16, pp. 241–286. ISBN 978-3-642-19269-2. [Google Scholar]
- Houde, M.; Taranu, Z.E.; Wang, X.; Young, B.; Gagnon, P.; Ferguson, S.H.; Kwan, M.; Muir, D.C.G. Mercury in Ringed Seals (Pusa hispida) from the Canadian Arctic in Relation to Time and Climate Parameters. Environ. Toxicol. Chem. 2020, 39, 2462–2474. [Google Scholar] [CrossRef]
- Tomy, G.T. Time-Trend Studies on New and Emerging Persistent Halogenated Compounds in Beluga Whales from Hendrickson Island (NWT) and Sanikiluaq (Nunavut). In Synopsis of Research Conducted under the 2013–2014 Northern Contaminants Program; Aboriginal Affairs and Northern Development Canada: Gatineau, QC, Canada, 2014; pp. 209–215. [Google Scholar]
- Desforges, J.-P.W.; Sonne, C.; Levin, M.; Siebert, U.; De Guise, S.; Dietz, R. Immunotoxic effects of environmental pollutants in marine mammals. Environ. Int. 2016, 86, 126–139. [Google Scholar] [CrossRef]
- Jepson, P.D.; Deaville, R.; Barber, J.L.; Aguilar, À.; Borrell, A.; Murphy, S.; Barry, J.; Brownlow, A.; Barnett, J.E.F.; Berrow, S.; et al. PCB pollution continues to impact populations of orcas and other dolphins in European waters. Sci. Rep. 2016, 6, 18573. [Google Scholar] [CrossRef]
- Zhang, R.; Guo, J.; Wu, F.; Mu, Y.; Giesy, J.P.; Chang, H.; Zhao, X.; Feng, C. Toxicity Reference Values for Polybrominated Diphenyl Ethers: Risk Assessment for Predatory Birds and Mammals from Two Chinese Lakes. In Reviews of Environmental Contamination and Toxicology; Whitacre, D., Ed.; Springer: Cham, Switzerland, 2014; pp. 111–137. [Google Scholar]
- Scheuhammer, A.; Braune, B.; Chan, H.M.; Frouin, H.; Krey, A.; Letcher, R.; Loseto, L.; Noël, M.; Ostertag, S.; Ross, P.; et al. Recent progress on our understanding of the biological effects of mercury in fish and wildlife in the Canadian Arctic. Sci. Total Environ. 2015, 509–510, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Newsome, S.D.; Clementz, M.T.; Koch, P.L. Using stable isotope biogeochemistry to study marine mammal ecology. Mar. Mammal Sci. 2010, 26, 509–572. [Google Scholar] [CrossRef]
- Castrillon, J.; Bengtson Nash, S. Evaluating cetacean body condition; a review of traditional approaches and new developments. Ecol. Evol. 2020, 10, 6144–6162. [Google Scholar] [CrossRef]
- MacMillan, K.; Hoover, C.; Iacozza, J.; Peyton, J.; Loseto, L. Body condition indicators: Assessing the influence of harvest location and potential thresholds for application in beluga monitoring. Ecol. Indic. 2019, 104, 145–155. [Google Scholar] [CrossRef]
- Quehenberger, O.; Dennis, E.A. The Human Plasma Lipidome. N. Engl. J. Med. 2012, 366, 668–669. [Google Scholar] [CrossRef] [PubMed]
- Pietiläinen, K.H.; Róg, T.; Seppänen-Laakso, T.; Virtue, S.; Gopalacharyulu, P.; Tang, J.; Rodriguez-Cuenca, S.; Maciejewski, A.; Naukkarinen, J.; Ruskeepää, A.L.; et al. Association of Lipidome Remodeling in the Adipocyte Membrane with Acquired Obesity in Humans. PLoS Biol. 2011, 9, e1000623. [Google Scholar] [CrossRef]
- Bernier-Graveline, A.; Lesage, V.; Cabrol, J.; Lair, S.; Michaud, R.; Rosabal, M.; Verreault, J. Lipid metabolites as indicators of body condition in highly contaminant-exposed belugas from the endangered St. Lawrence Estuary population (Canada). Environ. Res. 2021, 192, 110272. [Google Scholar] [CrossRef]
- Maia, M.L.; Sousa, S.; Pestana, D.; Faria, A.; Teixeira, D.; Delerue-Matos, C.; Domingues, V.F.; Calhau, C. Impact of brominated flame retardants on lipid metabolism: An in vitro approach. Environ. Pollut. 2022, 294, 118639. [Google Scholar] [CrossRef]
- Lockhart, W.L.; Stern, G.A.; Wagemann, R.; Hunt, R.V.; Metner, D.A.; DeLaronde, J.; Dunn, B.; Stewart, R.E.A.; Hyatt, C.K.; Harwood, L.; et al. Concentrations of mercury in tissues of beluga whales (Delphinapterus leucas) from several communities in the Canadian Arctic from 1981 to 2002. Sci. Total Environ. 2005, 351–352, 391–412. [Google Scholar] [CrossRef]
- Aubail, A.; Méndez-Fernandez, P.; Bustamante, P.; Churlaud, C.; Ferreira, M.; Vingada, J.V.; Caurant, F. Use of skin and blubber tissues of small cetaceans to assess the trace element content of internal organs. Mar. Pollut. Bull. 2013, 76, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Bouquegneau, J.-M.; Pillet, S.; Das, K.; Debacker, V. Heavy metals in marine mammals. In Toxicology of Marine Mammals; Vos, J.V., Bossart, G.D., Fournier, M., O’Shea, T., Eds.; Taylor and Francis Publishers: Washington, DC, USA, 2002; pp. 135–167. ISBN 0-203-16557-8. [Google Scholar]
- Valvi, D.; Walker, D.I.; Inge, T.; Bartell, S.M.; Jenkins, T.; Helmrath, M.; Ziegler, T.R.; La Merrill, M.A.; Eckel, S.P.; Conti, D.; et al. Environmental chemical burden in metabolic tissues and systemic biological pathways in adolescent bariatric surgery patients: A pilot untargeted metabolomic approach. Environ. Int. 2020, 143, 105957. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Cao, L.; Li, F.; Lian, P.; Zhao, J. Multiple biomarkers of the cytotoxicity induced by BDE-47 in human embryonic kidney cells. Chemosphere 2015, 126, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Iverson, S.J.; Field, C.; Don Bowen, W.; Blanchard, W. Quantitative fatty acid signature analysis: A new method of estimating predator diets. Ecol. Monogr. 2004, 74, 211–235. [Google Scholar] [CrossRef]
- Krahn, M.M.; Herman, D.P.; Ylitalo, G.M.; Sloan, C.A.; Burrows, D.G.; Hobbs, R.C.; Mahoney, B.A.; Yanagida, G.K.; Calambokidis, J.; Moore, S.E. Stratification of lipids, fatty acids and organochlorine contaminants in blubber of white whales and killer whales. J. Cetacean Res. Manag. 2004, 6, 175–189. [Google Scholar]
- Raach, M.; Lebeuf, M.; Pelletier, E. PBDEs and PCBs in the liver of the St Lawrence Estuary beluga (Delphinapterus leucas): A comparison of levels and temporal trends with the blubber. J. Environ. Monit. 2011, 13, 649–656. [Google Scholar] [CrossRef]
- Ewald, J.D.; Kirk, J.L.; Li, M.; Sunderland, E.M. Organ-specific differences in mercury speciation and accumulation across ringed seal (Phoca hispida) life stages. Sci. Total Environ. 2019, 650, 2013–2020. [Google Scholar] [CrossRef]
- Ikemoto, T.; Kunito, T.; Tanaka, H.; Baba, N.; Miyazaki, N.; Tanabe, S. Detoxification Mechanism of Heavy Metals in Marine Mammals and Seabirds: Interaction of Selenium with Mercury, Silver, Copper, Zinc, and Cadmium in Liver. Arch. Environ. Contam. Toxicol. 2004, 47, 402–413. [Google Scholar] [CrossRef]
- Milo, R.; Jorgensen, P.; Springer, M. How Quickly Do Different Cells in the Body Replace Themselves? Available online: http://book.bionumbers.org/how-quickly-do-different-cells-in-the-body-replace-themselves/ (accessed on 11 September 2021).
- Harwood, L.A.; Norton, P.; Day, B.; Hall, P.A. The Harvest of Beluga Whales in Canada’s Western Arctic: Hunter-Based Monitoring of the Size and Composition of the Catch. Arctic 2002, 55, 10–20. [Google Scholar] [CrossRef]
- Stewart, R.E.A.; Stewart, B.E.; Stirling, I.; Street, E. Counts of growth layer groups in cementum and dentine in ringed seals (Phoca hipsida). Mar. Mammal Sci. 1996, 12, 383–401. [Google Scholar] [CrossRef]
- Stewart, R.E.A.; Campana, S.E.; Jones, C.M.; Stewart, B.E. Bomb radiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates. Can. J. Zool. 2006, 84, 1840–1852. [Google Scholar] [CrossRef] [Green Version]
- Pleskach, K.; Hoang, W.; Chu, M.; Halldorson, T.; Loseto, L.; Ferguson, S.H.; Tomy, G.T. Use of mass spectrometry to measure aspartic acid racemization for ageing beluga whales. Mar. Mammal Sci. 2016, 32, 1370–1380. [Google Scholar] [CrossRef]
- Benskin, J.P.; Ikonomou, M.G.; Liu, J.; Veldhoen, N.; Dubetz, C.; Helbing, C.C.; Cosgrove, J.R. Distinctive metabolite profiles in in-migrating Sockeye salmon suggest sex-linked endocrine perturbation. Environ. Sci. Technol. 2014, 48, 11670–11678. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.Y.; Benskin, J.P.; Chandramouli, B.; Butler, H.; Helbing, C.C.; Cosgrove, J.R. Xenobiotics Produce Distinct Metabolomic Responses in Zebrafish Larvae (Danio rerio). Environ. Sci. Technol. 2016, 50, 6526–6535. [Google Scholar] [CrossRef]
- Environmental Protection Agency. Method 1668C-Chlorinated Biphenyl Congeners in Water, Soil, Sediment, Biosolids, and Tissue by HRGC/HRMS; Environmental Protection Agency: Washington, DC, USA, 2010.
- Environmental Protection Agency. Method 1614A-Brominated Diphenyl Ethers in Water, Soil, Sediment, and Tissue by HRGC/HRMS; Environmental Protection Agency: Washington, DC, USA, 2010.
- Ikonomou, M.G.; Fraser, T.L.; Crewe, N.F.; Fischer, M.B.; Rogers, I.H.; He, T.; Sather, P.J.; Lamb, R.F. A Comprehensive Multiresidue Ultra-Trace Analytical Method, Based on HRGC/HRMS, for the Determination of PCDDs, PCDFs, PCBs, PBDEs, PCDEs, and Organochlorine Pesticides in Six Different Environmental Matrices. Can. Tech. Rep. Fish. Aquat. Sci. Rapp. Tech. Can. Sci. Halieut. Aquat. 2001, 2389, 101. [Google Scholar]
- Environment and Climate Change Canada. Northern Contaminants Program (NCP)/Arctic Monitoring Assessment Program (AMAP) Quality Assurance (QA)/Quality Control (QC) Interlaboratory Comparison Study Round 14; 2021. [Google Scholar]
- Vaissie, P.; Monge, A.; Husson, F. Factoshiny: Perform Factorial Analysis from “FactoMineR” with a Shiny Application; R Package Version 2.4; 2021. Available online: https://cran.r-project.org/web/packages/Factoshiny/index.html (accessed on 11 September 2021).
- Fang, H.; Hong, H.; Liu, Z.; Perkins, R.; Kelly, R.; Beresney, J.; Tong, W.; Fowler, B.A. Omics Biomarkers in Risk Assessment: A Bioinformatics Perspective. Comput. Toxicol. Methods Appl. Risk Assess. 2013, 195–213. [Google Scholar] [CrossRef]
- Pike, N. Using false discovery rates for multiple comparisons in ecology and evolution. Methods Ecol. Evol. 2011, 2, 278–282. [Google Scholar] [CrossRef]
- Josse, J.; Husson, F. missMDA: A package for handling missing values in multivariate data analysis. J. Stat. Softw. 2016, 70, 1–31. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Year) | Length (cm) | Axial Girth (cm) | Blubber Thickness (cm) | Muscle δ13C (‰) | Muscle δ15N (‰) | Liver δ13C (‰) | Liver δ15N (‰) | |
|---|---|---|---|---|---|---|---|---|
| Ringed seals | ||||||||
| 2019-MLV-04 | 10 | 152 | 115 | 5.50 | −19.1 | 15.5 | −18.7 | 15.2 |
| 2019-MLV-08 | 19 | 125 | 100 | 3.00 | −18.6 | 15.4 | −18.7 | 15.1 |
| 2019-MLV-16 | 6 | 130 | 99 | 3.50 | −19.2 | 15.1 | −18.3 | 15.5 |
| 2019-MLV-18 | 12 | 137 | 102 | 3.50 | −18.9 | 15.5 | −19.0 | 15.0 |
| 2019-MLV-28 | 6 | 138 | 106 | 4.00 | −18.4 | 16.6 | −18.4 | 16.9 |
| 2019-MLV-29 | 6 | 150 | 101 | 5.00 | −18.2 | 15.9 | −18.5 | 15.8 |
| 2019-MLV-30 | 26 | 132 | 100 | 4.00 | −19.1 | 14.7 | −18.9 | 15.2 |
| 2019-MLV-31 | 26 | 136 | 100 | 3.00 | −18.6 | 15.7 | −18.7 | 15.3 |
| 2019-MLV-33 | 7 | 135 | 96 | 3.00 | −19.1 | 15.2 | −18.6 | 15.0 |
| 2019-MLV-40 | 14 | 244 | 177 | 7.00 | −17.4 | 14.8 | −17.4 | 14.8 |
| Mean ± SD | 13.1 ± 8.42 | 137 ± 8.79 | 102 ± 5.51 | 3.83 ± 0.90 | −18.8 ± 0.36 | 15.5 ± 0.54 | −18.6 ± 0.21 | 15.4 ± 0.60 |
| Belugas | ||||||||
| 2009-HI-02 | 24 | 432 | 117 | 7.60 | −20.9 | 19.1 | −20.3 | 18.3 |
| 2009-HI-07 | 23 | 366 | 122 | 11.4 | −23.8 | 18.7 | −20.1 | 18.3 |
| 2009-HI-09 | 32 | 422 | 129 | 8.90 | −23.0 | 19.2 | −21.4 | 18.1 |
| 2009-HI-10 | 25 | 427 | 129 | 10.2 | −21.8 | 18.8 | −20.9 | 18.2 |
| 2017-HI-04 | 19 | 437 | 117 | 9.20 | −18.9 | 17.5 | −21.3 | 18.0 |
| 2017-HI-07 | 29 | 417 | 104 | 8.90 | −18.9 | 17.2 | −20.8 | 17.6 |
| 2017-HI-09 | 63 | 442 | 130 | 12.1 | −19.2 | 17.3 | −21.5 | 17.6 |
| 2017-HI-10 | - | 433 | 127 | 10.8 | −19.1 | 17.3 | −20.9 | 18.0 |
| 2017-HI-11 | 29 | 406 | 130 | 11.4 | −19.4 | 17.3 | −20.7 | 17.8 |
| 2017-HI-12 | 30 | 420 | 150 | 9.50 | −19.4 | 17.2 | −21.0 | 17.6 |
| 2017-HI-13 | 44 | 424 | 133 | 11.4 | −19.5 | 17.0 | −20.9 | 17.9 |
| 2017-HI-14 | - | 399 | 110 | 8.30 | −19.3 | 17.2 | −21.1 | 17.6 |
| 2017-HI-15 | 38 | 419 | 107 | - | −19.3 | 17.6 | −21.1 | 17.8 |
| Mean ± SD | 32.4 ± 12.3 | 419 ± 19.8 | 123 ± 12.5 | 9.97 ± 1.44 | −20.2 ± 1.65 | 17.8 ± 0.83 | −20.9 ± 0.39 | 17.9 ± 0.27 |
| ΣPCB a,b (ng/g lw) | ΣPBDE c,d (ng/g lw) | HBB e (pg/g lw) | Total Hg (ng/g ww) | |
|---|---|---|---|---|
| Ringed seals | ||||
| 2019-MLV-04 | 827 | 158 | 3.81 | 1602 |
| 2019-MLV-08 | 1876 | 274 | 11.6 | 399 |
| 2019-MLV-16 | 884 | 145 | 17.0 | 357 |
| 2019-MLV-18 | 877 | 247 | 41.1 | 487 |
| 2019-MLV-28 | 753 | 91 | 10.6 | 1369 |
| 2019-MLV-29 | 582 | 80 | 12.9 | 931 |
| 2019-MLV-30 | 4587 | 1082 | 4.48 | 636 |
| 2019-MLV-31 | 3586 | 314 | 11.4 | 320 |
| 2019-MLV-33 | 1060 | 139 | 7.27 | 263 |
| Geometric mean ± SD | 1279 ± 1439 | 201 ± 311 | 10.5 ± 11.2 | 581 ± 488 |
| Belugas | ||||
| 2009-HI-02 | 1769 | 41.7 | n/a | 630 |
| 2009-HI-07 | 1777 | 34.7 | n/a | 535 |
| 2009-HI-09 | 3465 | 51.2 | n/a | 915 |
| 2009-HI-10 | 2447 | 30.2 | n/a | 681 |
| 2017-HI-04 | 1907 | 29.3 | n/a | 914 |
| 2017-HI-07 | 2253 | 19.3 | n/a | 974 |
| 2017-HI-09 | 4964 | 19.3 | n/a | 1729 |
| 2017-HI-10 | 2347 | 12.4 | n/a | 934 |
| 2017-HI-11 | 2219 | 14.2 | n/a | 741 |
| 2017-HI-12 | 1831 | 12.2 | n/a | 744 |
| 2017-HI-13 | 1438 | 12.6 | n/a | 703 |
| 2017-HI-14 | 1435 | 10.8 | n/a | 660 |
| 2017-HI-15 | 2091 | 16.4 | n/a | 686 |
| Geometric mean ± SD | 2168 ± 955 | 20.6 ± 12.9 | n/a | 798 ± 300 |
| Explanatory Variable | Class | Metabolite | Tissue | Correlation Coefficient | q-Value |
|---|---|---|---|---|---|
| Muscle δ13C | Phosphatidylcholine | PC aa C32:1 | Plasma | −0.87 | 0.033 |
| Phosphatidylcholine | PC aa C32:2 | Plasma | −0.83 | 0.049 | |
| Phosphatidylcholine | PC aa C34:1 | Plasma | −0.86 | 0.033 | |
| Phosphatidylcholine | PC aa C34:2 | Plasma | −0.90 | 0.033 | |
| Phosphatidylcholine | PC aa C34:3 | Plasma | −0.83 | 0.049 | |
| Phosphatidylcholine | PC aa C36:2 | Plasma | −0.88 | 0.033 | |
| Phosphatidylcholine | PC aa C36:5 | Plasma | −0.88 | 0.033 | |
| Phosphatidylcholine | PC aa C36:6 | Plasma | −0.86 | 0.033 | |
| Girth | Phosphatidylcholine | PC ae C36:5 | Liver | −0.85 | 0.027 |
| Sphingomyelin | SM C22:3 | Liver | −0.94 | 0.003 | |
| HBB concentration | Amino acid | Glu | Inner blubber | −0.74 | 0.045 |
| Phosphatidylcholine | PC aa C34:1 | Inner blubber | −0.94 | 0.006 | |
| Phosphatidylcholine | PC aa C36:1 | Inner blubber | −0.90 | 0.013 | |
| Phosphatidylcholine | PC aa C36:3 | Inner blubber | −0.80 | 0.029 | |
| Phosphatidylcholine | PC aa C36:4 | Inner blubber | −0.80 | 0.029 | |
| Phosphatidylcholine | PC aa C38:3 | Inner blubber | −0.77 | 0.035 | |
| Phosphatidylcholine | PC aa C38:4 | Inner blubber | −0.85 | 0.017 | |
| Phosphatidylcholine | PC aa C38:5 | Inner blubber | −0.85 | 0.016 | |
| Phosphatidylcholine | PC ae C32:1 | Inner blubber | −0.71 | 0.050 | |
| Phosphatidylcholine | PC ae C34:1 | Inner blubber | −0.84 | 0.018 | |
| Phosphatidylcholine | PC ae C34:2 | Inner blubber | −0.78 | 0.033 | |
| Phosphatidylcholine | PC ae C34:3 | Inner blubber | −0.73 | 0.045 | |
| Phosphatidylcholine | PC ae C36:0 | Inner blubber | −0.80 | 0.029 | |
| Phosphatidylcholine | PC ae C36:1 | Inner blubber | −0.77 | 0.035 | |
| Phosphatidylcholine | PC ae C36:2 | Inner blubber | −0.88 | 0.016 | |
| Phosphatidylcholine | PC ae C36:3 | Inner blubber | −0.79 | 0.029 | |
| Phosphatidylcholine | PC ae C36:4 | Inner blubber | −0.85 | 0.016 | |
| Phosphatidylcholine | PC ae C36:5 | Inner blubber | −0.72 | 0.050 | |
| Phosphatidylcholine | PC ae C38:1 | Inner blubber | −0.87 | 0.016 | |
| Phosphatidylcholine | PC ae C40:2 | Inner blubber | -0.93 | 0.006 | |
| Sphingomyelin | SM (OH) C16:1 | Inner blubber | −0.71 | 0.050 | |
| Sphingomyelin | SM (OH) C22:1 | Inner blubber | −0.85 | 0.016 | |
| Sphingomyelin | SM (OH) C22:2 | Inner blubber | −0.75 | 0.041 | |
| Sphingomyelin | SM (OH) C24:1 | Inner blubber | −0.73 | 0.045 | |
| Sphingomyelin | SM C18:0 | Inner blubber | −0.77 | 0.035 | |
| Sphingomyelin | SM C24:1 | Inner blubber | −0.87 | 0.016 |
| Explanatory Variable | Class | Metabolite | Tissue | Correlation Coefficient | q-Value |
|---|---|---|---|---|---|
| Girth | Acylcarnitine | AC C9 | Plasma | 0.79 | 0.025 |
| Hg concentration | Acylcarnitine | AC C14:1 | Inner blubber | 0.82 | 0.011 |
| Acylcarnitine | AC C18:2 | Inner blubber | 0.85 | 0.010 | |
| Phosphatidylcholine | PC aa C36:0 | Inner blubber | 0.91 | 0.004 | |
| Phosphatidylcholine | PC aa C42:1 | Inner blubber | −0.76 | 0.024 | |
| Sphingomyelin | SM C24:1 | Inner blubber | 0.71 | 0.032 | |
| PBDE concentration | Amino acid | Asn | Outer blubber | −0.89 | 0.024 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simond, A.É.; Noël, M.; Loseto, L.; Houde, M.; Kirk, J.; Elliott, A.; Brown, T.M. A Multi-Matrix Metabolomic Approach in Ringed Seals and Beluga Whales to Evaluate Contaminant and Climate-Related Stressors. Metabolites 2022, 12, 813. https://doi.org/10.3390/metabo12090813
Simond AÉ, Noël M, Loseto L, Houde M, Kirk J, Elliott A, Brown TM. A Multi-Matrix Metabolomic Approach in Ringed Seals and Beluga Whales to Evaluate Contaminant and Climate-Related Stressors. Metabolites. 2022; 12(9):813. https://doi.org/10.3390/metabo12090813
Chicago/Turabian StyleSimond, Antoine É., Marie Noël, Lisa Loseto, Magali Houde, Jane Kirk, Ashley Elliott, and Tanya M. Brown. 2022. "A Multi-Matrix Metabolomic Approach in Ringed Seals and Beluga Whales to Evaluate Contaminant and Climate-Related Stressors" Metabolites 12, no. 9: 813. https://doi.org/10.3390/metabo12090813