
Comparative Metabolomics Study of the Impact of Articaine and Lidocaine on the Metabolism of SH-SY5Y Neuronal Cells

 ,
,  ,
,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Cell Viability
2.2. 1H NMR-Metabolomics
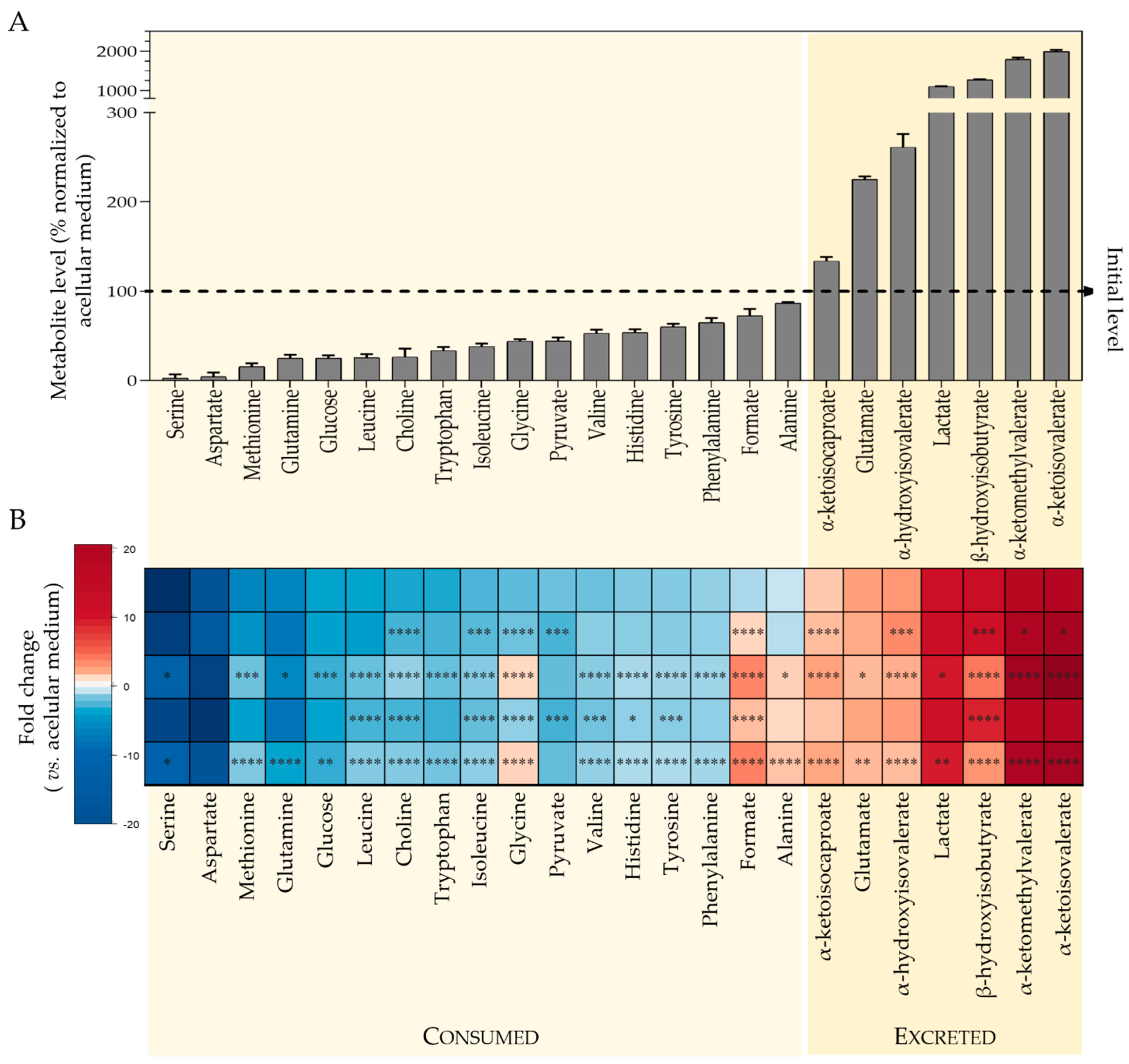
2.2.1. Metabolic Profiling of Culture Medium
2.2.2. Metabolic Profiling of Cell Aqueous Extracts
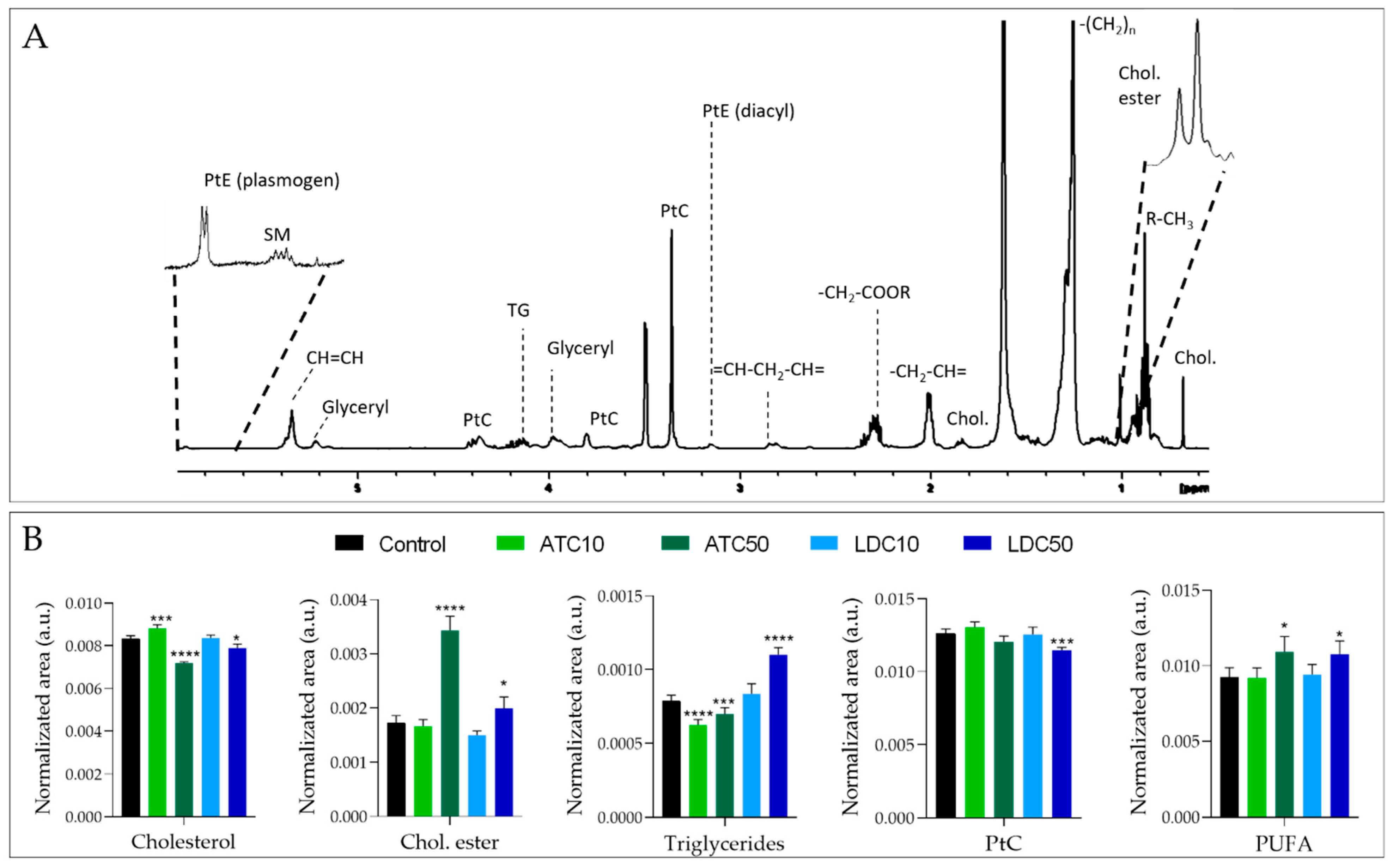
2.2.3. Metabolic Profiling of Cell Lipid Extracts
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture and Viability Assay
4.3. 1H NMR Metabolomics
4.3.1. Exposure of SH-SY5Y to the Local Anesthetics
4.3.2. Sample Extraction
4.3.3. NMR Data Acquisition and Processing
4.3.4. Multivariate Analysis and Spectral Integration
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seward, G.R. Handbook of Local Anesthesia; Elsevier: Amsterdam, The Netherlands, 1981; Volume 9, ISBN 9780323074131. [Google Scholar]
- Katyal, V. The Efficacy and Safety of Articaine versus Lignocaine in Dental Treatments: A Meta-Analysis. J. Dent. 2010, 38, 307–317. [Google Scholar] [CrossRef]
- Zhang, A.; Tang, H.; Liu, S.; Ma, C.; Ma, S.; Zhao, H. Anesthetic Efficiency of Articaine Versus Lidocaine in the Extraction of Lower Third Molars: A Meta-Analysis and Systematic Review. J. Oral Maxillofac. Surg. 2019, 77, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Kakroudi, S.H.A.; Mehta, S.; Millar, B.J. Articaine Hydrochloride: Is It the Solution? Dent. Update 2015, 42, 88–93. [Google Scholar] [CrossRef]
- Bartlett, G.; Mansoor, J. Articaine Buccal Infiltration vs Lidocaine Inferior Dental Block—A Review of the Literature. BDJ 2016, 220, 117–120. [Google Scholar] [CrossRef]
- Malamed, S.F. Sedation-E-Book: A Guide to Patient Management; Elsevier: Maryland Heights, MI, USA, 2009; ISBN 0323075967. [Google Scholar]
- de Araújo, D.R.; Ribeiro, L.N.D.M.; de Paula, E. Lipid-Based Carriers for the Delivery of Local Anesthetics. Expert Opin. Drug Deliv. 2019, 16, 701–714. [Google Scholar] [CrossRef]
- Prates, É.T.; Rodrigues da Silva, G.H.; Souza, T.F.; Skaf, M.S.; Pickholz, M.; de Paula, E. Articaine Interaction with Phospholipid Bilayers. J. Mol. Struct. 2020, 1222, 128854. [Google Scholar] [CrossRef]
- Fraceto, L.F.; Spisni, A.; Schreier, S.; de Paula, E. Differential Effects of Uncharged Aminoamide Local Anesthetics on Phospholipid Bilayers, as Monitored by 1H-NMR Measurements. Biophys. Chem. 2005, 115, 11–18. [Google Scholar] [CrossRef]
- Onal, O.; Saltali, A.O.; Apiliogullari, S. Comments on “Local Anesthetic Systemic Toxicity”. Aesthetic Surg. J. 2016, 36, NP37. [Google Scholar] [CrossRef] [Green Version]
- Malamed, S.F.; Gagnon, S.; Leblanc, D. Articaine Hydrochloride: A Study of the Safety of a New Amide Local Anesthetic. J. Am. Dent. Assoc. 2001, 132, 177–185. [Google Scholar] [CrossRef]
- Garlsto, G.A.; Gaffen, A.S.; Lawrence, H.P.; Tenenbaum, H.C.; Haas, D.A. Occurrence of Paresthesia after Dental Local Anesthetic Administration in the United States. J. Am. Dent. Assoc. 2010, 141, 836–844. [Google Scholar] [CrossRef] [Green Version]
- Kingon, A.; Sambrook, P.; Goss, A. Higher Concentration Local Anaesthetics Causing Prolonged Anaesthesia. Do They? A Literature Review and Case Reports. Aust. Dent. J. 2011, 56, 348–351. [Google Scholar] [CrossRef]
- Hillerup, S.; Jensen, R. Nerve Injury Caused by Mandibular Block Analgesia. Int. J. Oral Maxillofac. Surg. 2006, 35, 437–443. [Google Scholar] [CrossRef]
- Ramadurai, N.; Gurunathan, D.; Samuel, A.V.; Subramanian, E.; Rodrigues, S.J.L. Effectiveness of 2% Articaine as an Anesthetic Agent in Children: Randomized Controlled Trial. Clin. Oral Investig. 2019, 23, 3543–3550. [Google Scholar] [CrossRef]
- Hopman, A.J.G.; Baart, J.A.; Brand, H.S. Articaine and Neurotoxicity—A Review. Br. Dent. J. 2017, 223, 501–506. [Google Scholar] [CrossRef]
- Stirrup, P.; Crean, S. Does Articaine, Rather than Lidocaine, Increase the Risk of Nerve Damage When Administered for Inferior Alveolar Nerve Blocks in Patients Undergoing Local Anaesthesia for Dental Treatment? A Mini Systematic Review of the Literature. Br. Dent. J. 2019, 226, 213–223. [Google Scholar] [CrossRef]
- Malamed, S. Articaine 30 Years Later. Available online: https://www.oralhealthgroup.com/features/1003919408/ (accessed on 13 April 2020).
- Verlinde, M.; Hollmann, M.W.; Stevens, M.F.; Hermanns, H.; Werdehausen, R.; Lirk, P. Local Anesthetic-Induced Neurotoxicity. Int. J. Mol. Sci. 2016, 17, 339. [Google Scholar] [CrossRef]
- Kim, E.-J.; Kim, H.Y.; Ahn, J.-H. Neurotoxicity of Local Anesthetics in Dentistry. J. Dent. Anesth. Pain Med. 2020, 20, 55. [Google Scholar] [CrossRef]
- Johnson, M.E.; Uhl, C.B.; Spittler, K.-H.; Wang, H.; Gores, G.J. Mitochondrial Injury and Caspase Activation by the Local Anesthetic Lidocaine. Anesthesiology 2004, 101, 1184–1194. [Google Scholar] [CrossRef]
- Okamoto, A.; Tanaka, M.; Sumi, C.; Oku, K.; Kusunoki, M.; Nishi, K.; Matsuo, Y.; Takenaga, K.; Shingu, K.; Hirota, K. The Antioxidant N-Acetyl Cysteine Suppresses Lidocaine-Induced Intracellular Reactive Oxygen Species Production and Cell Death in Neuronal SH-SY5Y Cells. BMC Anesthesiol. 2016, 16, 104. [Google Scholar] [CrossRef] [Green Version]
- Werdehausen, R.; Fazeli, S.; Braun, S.; Hermanns, H.; Essmann, F.; Hollmann, M.W.; Bauer, I.; Stevens, M.F. Apoptosis Induction by Different Local Anaesthetics in a Neuroblastoma Cell Line. Br. J. Anaesth. 2009, 103, 711–718. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, A.; Sumi, C.; Tanaka, H.; Kusunoki, M.; Iwai, T.; Nishi, K.; Matsuo, Y.; Harada, H.; Takenaga, K.; Bono, H.; et al. HIF-1-Mediated Suppression of Mitochondria Electron Transport Chain Function Confers Resistance to Lidocaine-Induced Cell Death. Sci. Rep. 2017, 7, 3816. [Google Scholar] [CrossRef] [Green Version]
- Haller, I.; Hausott, B.; Tomaselli, B.; Keller, C.; Klimaschewski, L.; Gerner, P.; Lirk, P. Neurotoxicity of Lidocaine Involves Specific Activation of the P38 Mitogen-Activated Protein Kinase, but not Extracellular Signal-Regulated or c-Jun N-Terminal Kinases, and Is Mediated by Arachidonic Acid Metabolites. Anesthesiology 2006, 105, 1024–1033. [Google Scholar] [CrossRef]
- Albalawi, F.; Lim, J.C.; DiRenzo, K.V.; Hersh, E.V.; Mitchell, C.H. Effects of Lidocaine and Articaine on Neuronal Survival and Recovery. Anesth. Prog. 2018, 65, 82–88. [Google Scholar] [CrossRef]
- Malet, A.; Faure, M.O.; Deletage, N.; Pereira, B.; Haas, J.; Lambert, G. The Comparative Cytotoxic Effects of Different Local Anesthetics on a Human Neuroblastoma Cell Line. Anesth. Analg. 2015, 120, 589–596. [Google Scholar] [CrossRef]
- Kovalevich, J.; Langford, D. Considerations for the Use of SH-SY5Y Neuroblastoma Cells in Neurobiology. Methods Mol. Biol. 2013, 1078, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Schultz, L.; Zurich, M.G.; Culot, M.; da Costa, A.; Landry, C.; Bellwon, P.; Kristl, T.; Hörmann, K.; Ruzek, S.; Aiche, S.; et al. Evaluation of Drug-Induced Neurotoxicity Based on Metabolomics, Proteomics and Electrical Activity Measurements in Complementary CNS in vitro Models. Toxicol Vitr. 2015, 30, 138–165. [Google Scholar] [CrossRef]
- Boone, C.H.T.; Grove, R.A.; Adamcova, D.; Seravalli, J.; Adamec, J. Oxidative Stress, Metabolomics Profiling, and Mechanism of Local Anesthetic Induced Cell Death in Yeast. Redox Biol. 2017, 12, 139–149. [Google Scholar] [CrossRef]
- Chamaraux-Tran, T.N.; Muller, M.; Pottecher, J.; Diemunsch, P.A.; Tomasetto, C.; Namer, I.J.; Dali-Youcef, N. Metabolomic Impact of Lidocaine on a Triple Negative Breast Cancer Cell Line. Front. Pharmacol. 2022, 13, 821779. [Google Scholar] [CrossRef]
- Gong, Q.; Wen, X.; Li, H.; He, J.; Wang, Y.; Wu, H.; Wang, H.; Wang, X. Up-Regulation of Cav3.1 Expression in SH-SY5Y Cells Induced by Lidocaine Hydrochloride. Artif. Cells Nanomed. Biotechnol. 2018, 46, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, Q.; Lu, J.; Cao, J.; Wang, X.Y.; Chen, Y. Lidocaine Promotes Autophagy of SH-SY5Y Cells through Inhibiting PI3K/AKT/MTOR Pathway by Upregulating MiR-145. Toxicol. Res. 2020, 9, 467–473. [Google Scholar] [CrossRef]
- Werdehausen, R.; Braun, S.; Fazeli, S.; Hermanns, H.; Hollmann, M.W.; Bauer, I.; Stevens, M.F. Lipophilicity but not Stereospecificity Is a Major Determinant of Local Anaesthetic-Induced Cytotoxicity in Human T-Lymphoma Cells. Eur. J. Anaesthesiol. 2012, 29, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Smith, J.; Nie, H.; Wang, Z.; Erwin, P.J.; van Wijnen, A.J.; Qu, W. Cytotoxicity of Local Anesthetics in Mesenchymal Stem Cells. Am. J. Phys. Med. Rehabil. 2018, 97, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Baroni, D.B.; Franz-Montan, M.; Cogo, K.; Berto, L.A.; Volpato, M.C.; Novaes, P.D.; Groppo, F.C. Effect of Articaine on Mental Nerve Anterior Portion: Histological Analysis in Rats. Acta Odontol. Scand. 2012, 71, 82–87. [Google Scholar] [CrossRef]
- de Paula, E.; Schreier, S. Use of a Novel Method for Determination of Partition Coefficients to Compare the Effect of Local Anesthetics on Membrane Structure. Biochim. Biophys. Acta 1995, 1240, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Jha, M.K.; Morrison, B.M. Glia-Neuron Energy Metabolism in Health and Diseases: New Insights into the Role of Nervous System Metabolic Transporters. Exp. Neurol. 2018, 309, 23–31. [Google Scholar] [CrossRef]
- Simpson, I.A.; Dwyer, D.; Malide, D.; Moley, K.H.; Travis, A.; Vannucci, S.J. The Facilitative Glucose Transporter GLUT3: 20 Years of Distinction. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E242. [Google Scholar] [CrossRef] [Green Version]
- Yellen, G. Fueling Thought: Management of Glycolysis and Oxidative Phosphorylation in Neuronal Metabolism. J. Cell Biol. 2018, 217, 2235–2246. [Google Scholar] [CrossRef]
- Alherz, M.; Lee, D.; Alshangiti, A.; Roddy, D.; O’Keeffe, G.; White, R.; Barry, D. The Growth Response to Beta-Hydroxybutyrate in SH-SY5Y Neuroblastoma Cells Is Suppressed by Glucose and Pyruvate Supplementation. Neurochem. Res. 2021, 46, 701–709. [Google Scholar] [CrossRef]
- Uesono, Y.; Toh-e, A.; Kikuchi, Y.; Araki, T.; Hachiya, T.; Watanabe, C.; Noguchi, K.; Terashima, I. Local Anesthetics and Antipsychotic Phenothiazines Interact Nonspecifically with Membranes and Inhibit Hexose Transporters in Yeast. Genetics 2016, 202, 997–1012. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, D.; Beitner, R. Detachment of the Glycolytic Enzymes, Phosphofructokinase and Aldolase, from Cytoskeleton of Melanoma Cells, Induced by Local Anesthetics. Mol. Genet. Metab. 2000, 69, 159–164. [Google Scholar] [CrossRef]
- Boone, C.H.T.; Grove, R.A.; Adamcova, D.; Braga, C.P.; Adamec, J. Revealing Oxidative Damage to Enzymes of Carbohydrate Metabolism in Yeast: An Integration of 2D DIGE, Quantitative Proteomics, and Bioinformatics. Proteomics 2016, 16, 1889–1903. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.E.; Suh, P.G.; Kim, J.I. O-GlcNAcylation in Health and Neurodegenerative Diseases. Exp. Mol. Med. 2021, 53, 1674–1682. [Google Scholar] [CrossRef] [PubMed]
- McKenna, M.C.; Waagepetersen, H.S.; Schousboe, A.; Sonnewald, U. Neuronal and Astrocytic Shuttle Mechanisms for Cytosolic-Mitochondrial Transfer of Reducing Equivalents: Current Evidence and Pharmacological Tools. Biochem. Pharmacol. 2006, 71, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Maugard, M.; Vigneron, P.A.; Bolaños, J.P.; Bonvento, G. L-Serine Links Metabolism with Neurotransmission. Prog. Neurobiol. 2020, 197, 101896. [Google Scholar] [CrossRef]
- Meiser, J.; Tumanov, S.; Maddocks, O.; Labuschagne, C.F.; Athineos, D.; van den Broek, N.; Mackay, G.M.; Gottlieb, E.; Blyth, K.; Vousden, K.; et al. Serine One-Carbon Catabolism with Formate Overflow. Sci. Adv. 2016, 2, e1601273. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.V.; Markussen, K.H.; Jakobsen, E.; Schousboe, A.; Waagepetersen, H.S.; Rosenberg, P.A.; Aldana, B.I. Glutamate Metabolism and Recycling at the Excitatory Synapse in Health and Neurodegeneration. Neuropharmacology 2021, 196, 108719. [Google Scholar] [CrossRef]
- Sperringer, J.E.; Addington, A.; Hutson, S.M. Branched-Chain Amino Acids and Brain Metabolism. Neurochem. Res. 2017, 42, 1697–1709. [Google Scholar] [CrossRef]
- Gondáš, E.; Král’Ová Trančíková, A.; Baranovičová, E.; Šofranko, J.; Hatok, J.; Kowtharapu, B.S.; Galanda, T.; Dobrota, D.; Kubatka, P.; Busselberg, D.; et al. Expression of 3-Methylcrotonyl-CoA Carboxylase in Brain Tumors and Capability to Catabolize Leucine by Human Neural Cancer Cells. Cancers 2022, 14, 585. [Google Scholar] [CrossRef]
- Engskog, M.K.R.; Ersson, L.; Haglöf, J.; Arvidsson, T.; Pettersson, C.; Brittebo, E. β-N-Methylamino-l-Alanine (BMAA) Perturbs Alanine, Aspartate and Glutamate Metabolism Pathways in Human Neuroblastoma Cells as Determined by Metabolic Profiling. Amino Acids 2017, 49, 905–919. [Google Scholar] [CrossRef]
- Gunda, V.; Pathania, A.S.; Chava, S.; Prathipati, P.; Chaturvedi, N.K.; Coulter, D.W.; Pandey, M.K.; Durden, D.L.; Challagundla, K.B. Amino Acids Regulate Cisplatin Insensitivity in Neuroblastoma. Cancers 2020, 12, 2576. [Google Scholar] [CrossRef]
- Delic, V.; Griffin, J.W.D.; Zivkovic, S.; Zhang, Y.; Phan, T.A.; Gong, H.; Chaput, D.; Reynes, C.; Dinh, V.B.; Cruz, J.; et al. Individual Amino Acid Supplementation Can Improve Energy Metabolism and Decrease ROS Production in Neuronal Cells Overexpressing Alpha-Synuclein. NeuroMol. Med. 2017, 19, 322–344. [Google Scholar] [CrossRef] [PubMed]
- la Monaca, E.; Fodale, V. Effects of Anesthetics on Mitochondrial Signaling and Function. Curr. Drug Saf. 2012, 7, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Amo, T.; Oji, Y.; Saiki, S.; Hattori, N. Metabolomic Analysis Revealed Mitochondrial Dysfunction and Aberrant Choline Metabolism in MPP+-Exposed SH-SY5Y Cells. Biochem. Biophys. Res. Commun. 2019, 519, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Vettore, L.A.; Westbrook, R.L.; Tennant, D.A. Proline Metabolism and Redox; Maintaining a Balance in Health and Disease. Amino Acids 2021, 53, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Pandhare, J.; Donald, S.P.; Cooper, S.K.; Phang, J.M. Regulation and Function of Proline Oxidase under Nutrient Stress. J. Cell. Biochem. 2009, 107, 759–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Glunde, K.; Bhujwalla, Z.M.; Raman, V.; Sharma, A.; Phang, J.M. Proline Oxidase Promotes Tumor Cell Survival in Hypoxic Tumor Microenvironments. Cancer Res. 2012, 72, 3677–3686. [Google Scholar] [CrossRef] [Green Version]
- Chhetri, D.R. Myo-Inositol and Its Derivatives: Their Emerging Role in the Treatment of Human Diseases. Front. Pharmacol. 2019, 10, 1172. [Google Scholar] [CrossRef] [Green Version]
- Fisher, S.K.; Novak, J.E.; Agranoff, B.W. Inositol and Higher Inositol Phosphates in Neural Tissues: Homeostasis, Metabolism and Functional Significance. J. Neurochem. 2002, 82, 736–754. [Google Scholar] [CrossRef]
- Loveday, D.; Heacock, A.M.; Fisher, S.K. Activation of Muscarinic Cholinergic Receptors Enhances the Volume-Sensitive Efflux of Myo-Inositol from SH-SY5Y Neuroblastoma Cells. J. Neurochem. 2003, 87, 476–486. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Inazu, M.; Tajima, H.; Matsumiya, T. Functional Expression of Choline Transporter-like Protein 1 (CTL1) in Human Neuroblastoma Cells and Its Link to Acetylcholine Synthesis. Neurochem. Int. 2011, 58, 354–365. [Google Scholar] [CrossRef]
- Ralhan, I.; Chang, C.L.; Lippincott-Schwartz, J.; Ioannou, M.S. Lipid Droplets in the Nervous System. J. Cell Biol. 2021, 220, e202102136. [Google Scholar] [CrossRef] [PubMed]
- Islimye, E.; Girard, V.; Gould, A.P. Functions of Stress-Induced Lipid Droplets in the Nervous System. Front. Cell Dev. Biol. 2022, 10, 863907. [Google Scholar] [CrossRef]
- Kostidis, S.; Addie, R.D.; Morreau, H.; Mayboroda, O.A.; Giera, M. Quantitative NMR Analysis of Intra- and Extracellular Metabolism of Mammalian Cells: A Tutorial. Anal. Chim. Acta 2017, 980, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Carrola, J.; Bastos, V.; Jarak, I.; Oliveira-Silva, R.; Malheiro, E.; Daniel-da-Silva, A.L.; Oliveira, H.; Santos, C.; Gil, A.M.; Duarte, I.F. Metabolomics of Silver Nanoparticles Toxicity in HaCaT Cells: Structure–Activity Relationships and Role of Ionic Silver and Oxidative Stress. Nanotoxicology 2016, 10, 1105–1117. [Google Scholar] [CrossRef] [PubMed]
- Berben, L.; Sereika, S.M.; Engberg, S. Effect Size Estimation: Methods and Examples. Int. J. Nurs. Stud. 2012, 49, 1039–1047. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, G.H.R.; Mendes, L.F.; de Carvalho, F.V.; de Paula, E.; Duarte, I.F. Comparative Metabolomics Study of the Impact of Articaine and Lidocaine on the Metabolism of SH-SY5Y Neuronal Cells. Metabolites 2022, 12, 581. https://doi.org/10.3390/metabo12070581
da Silva GHR, Mendes LF, de Carvalho FV, de Paula E, Duarte IF. Comparative Metabolomics Study of the Impact of Articaine and Lidocaine on the Metabolism of SH-SY5Y Neuronal Cells. Metabolites. 2022; 12(7):581. https://doi.org/10.3390/metabo12070581
Chicago/Turabian Styleda Silva, Gustavo H. Rodrigues, Luís F. Mendes, Fabíola V. de Carvalho, Eneida de Paula, and Iola F. Duarte. 2022. "Comparative Metabolomics Study of the Impact of Articaine and Lidocaine on the Metabolism of SH-SY5Y Neuronal Cells" Metabolites 12, no. 7: 581. https://doi.org/10.3390/metabo12070581