
Untargeted Metabolomics Profiling Reveals Perturbations in Arginine-NO Metabolism in Middle Eastern Patients with Coronary Heart Disease

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
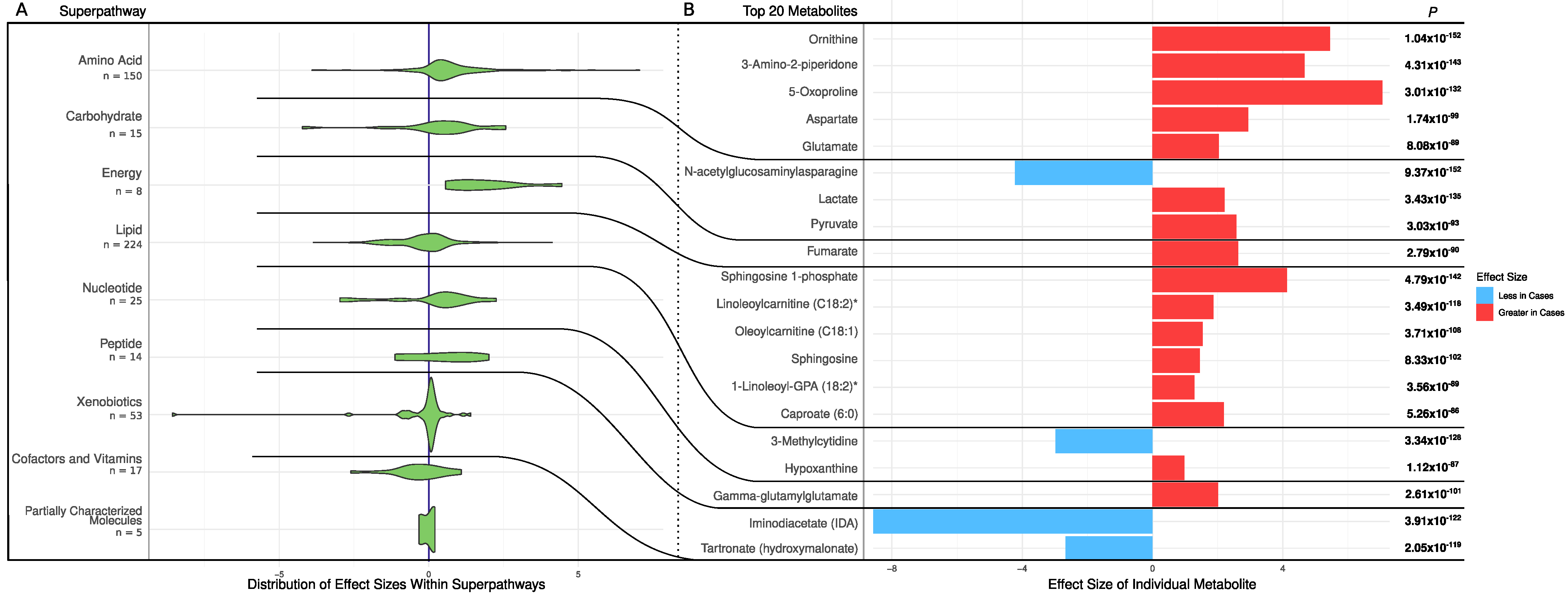
2.1. Univariate Analysis
2.2. Pathway Enrichment Analysis
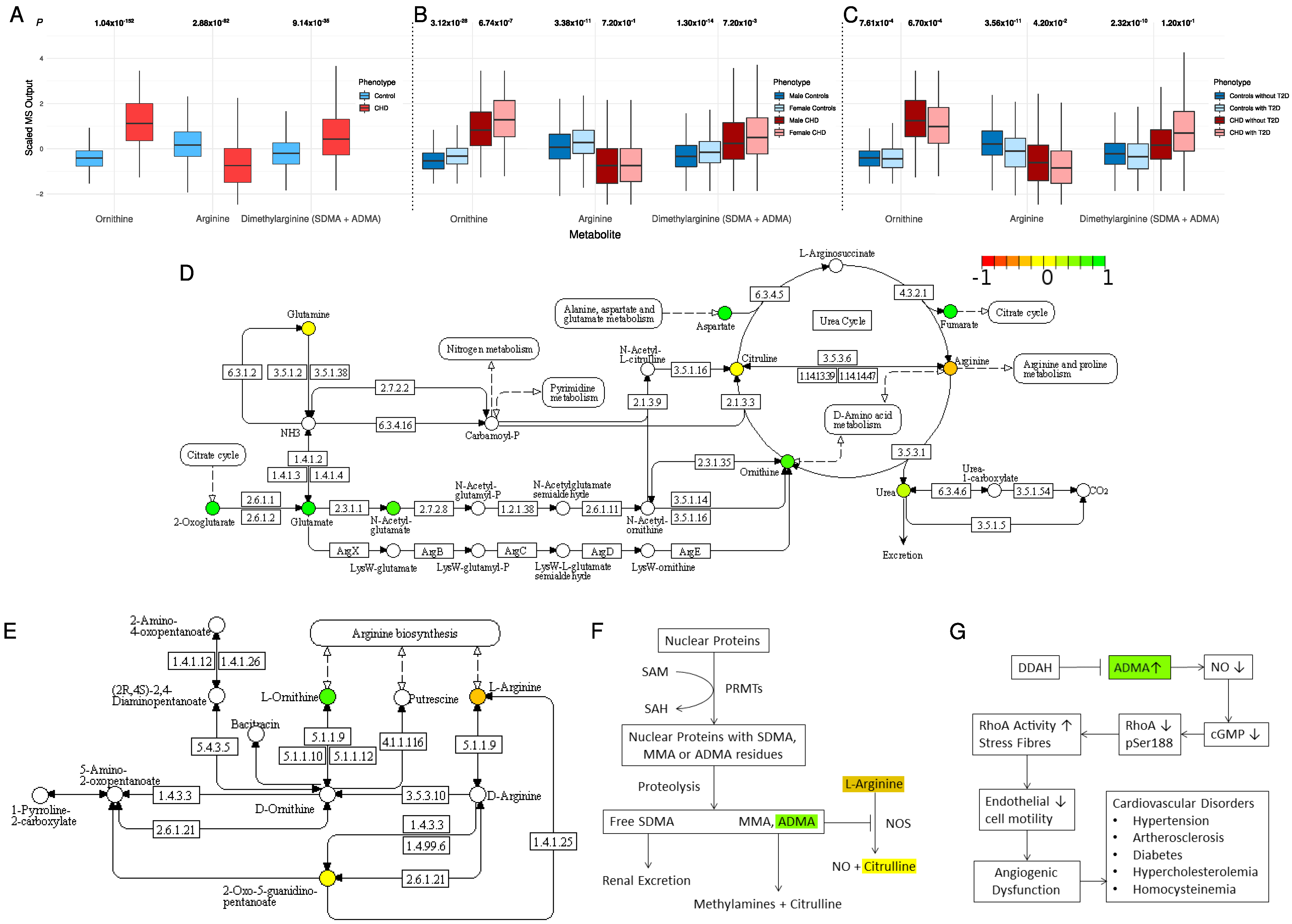
2.3. Alternation in Arginine and Ornithine Metabolism and Synthesis
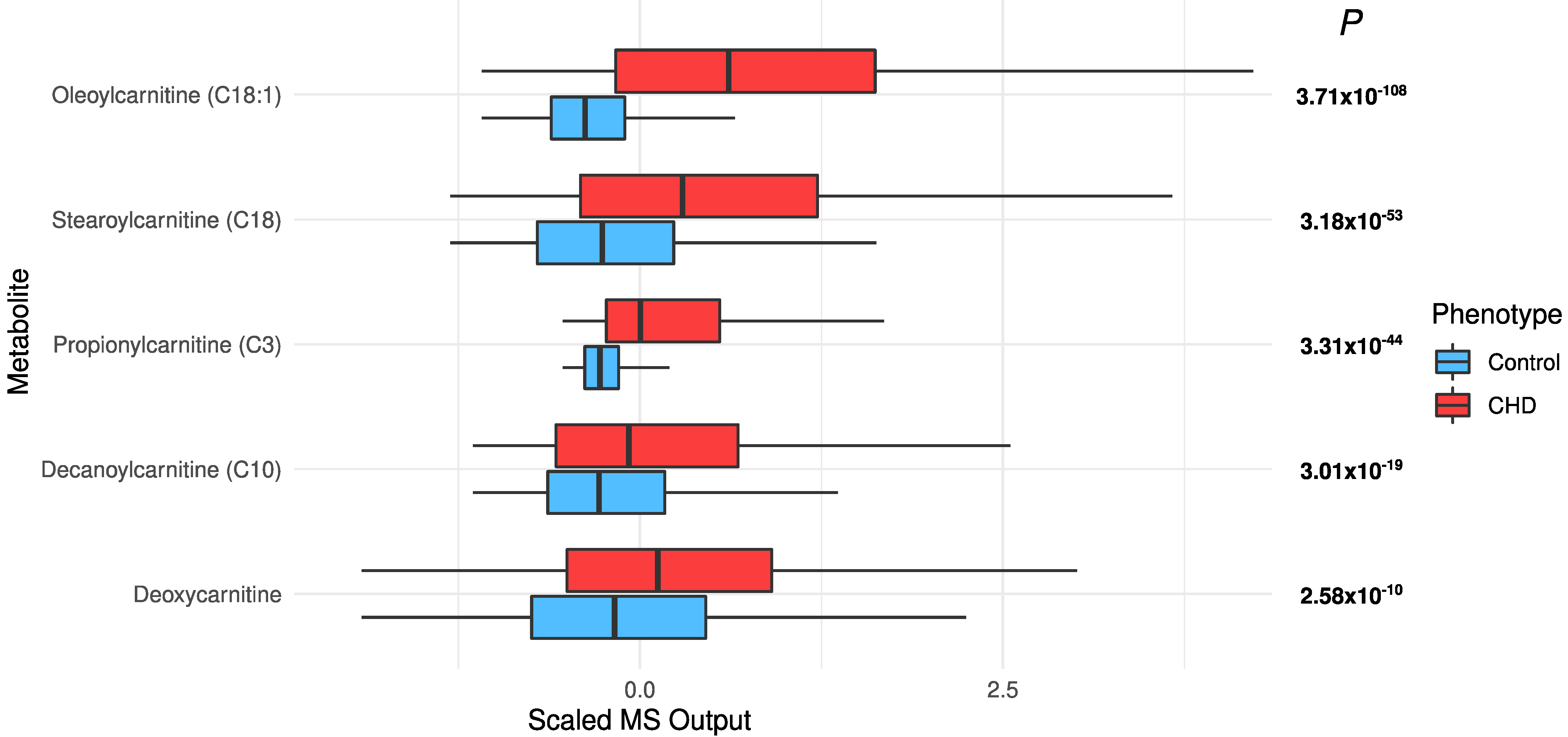
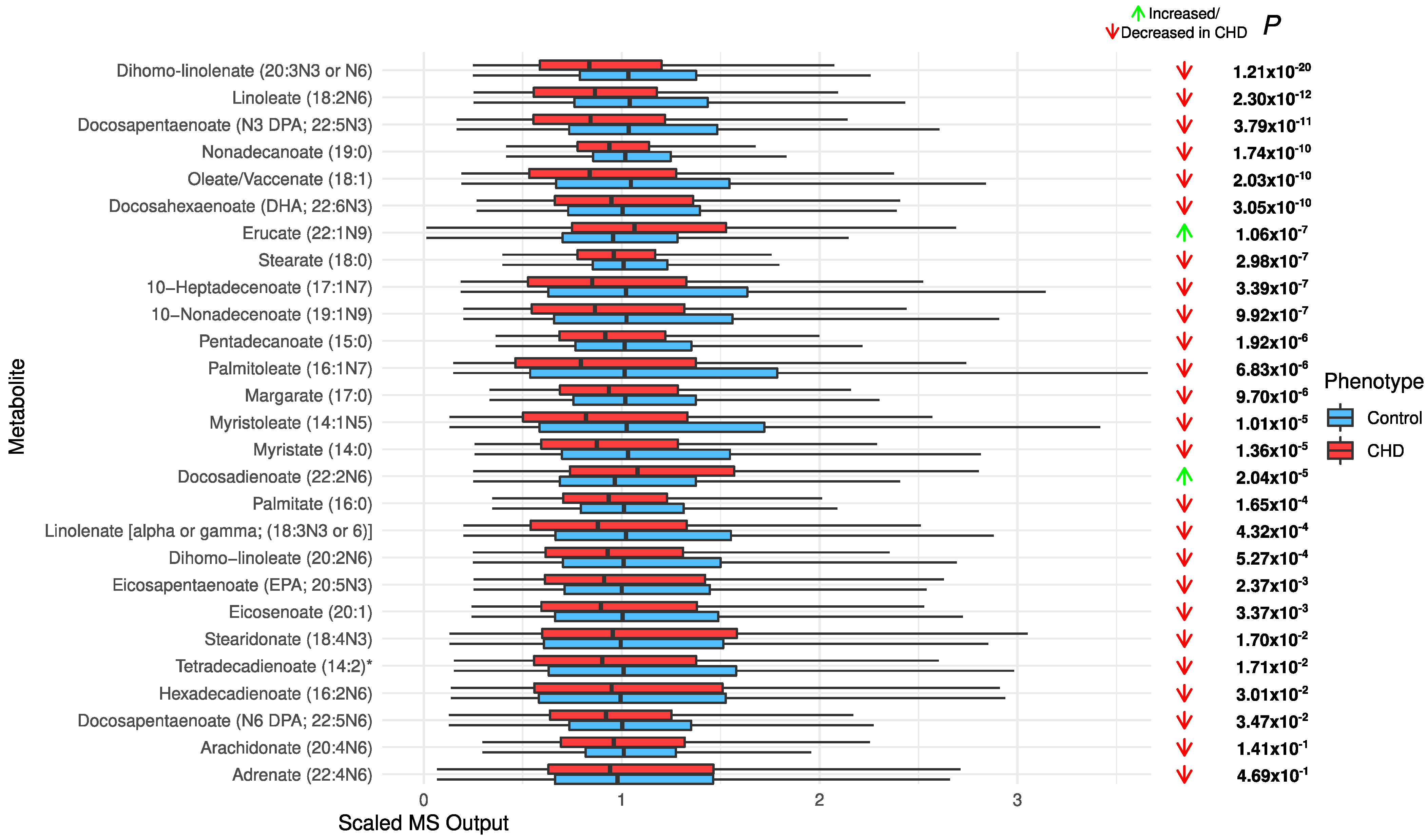
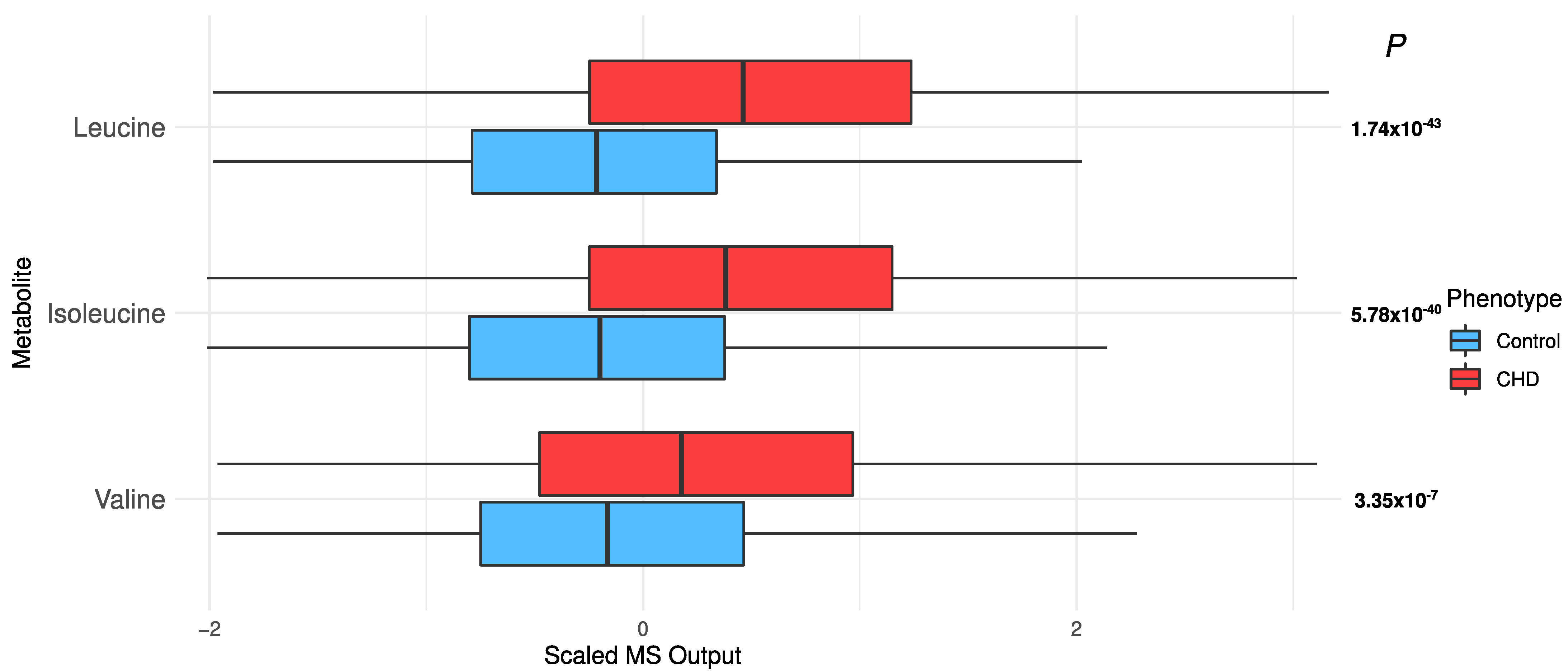
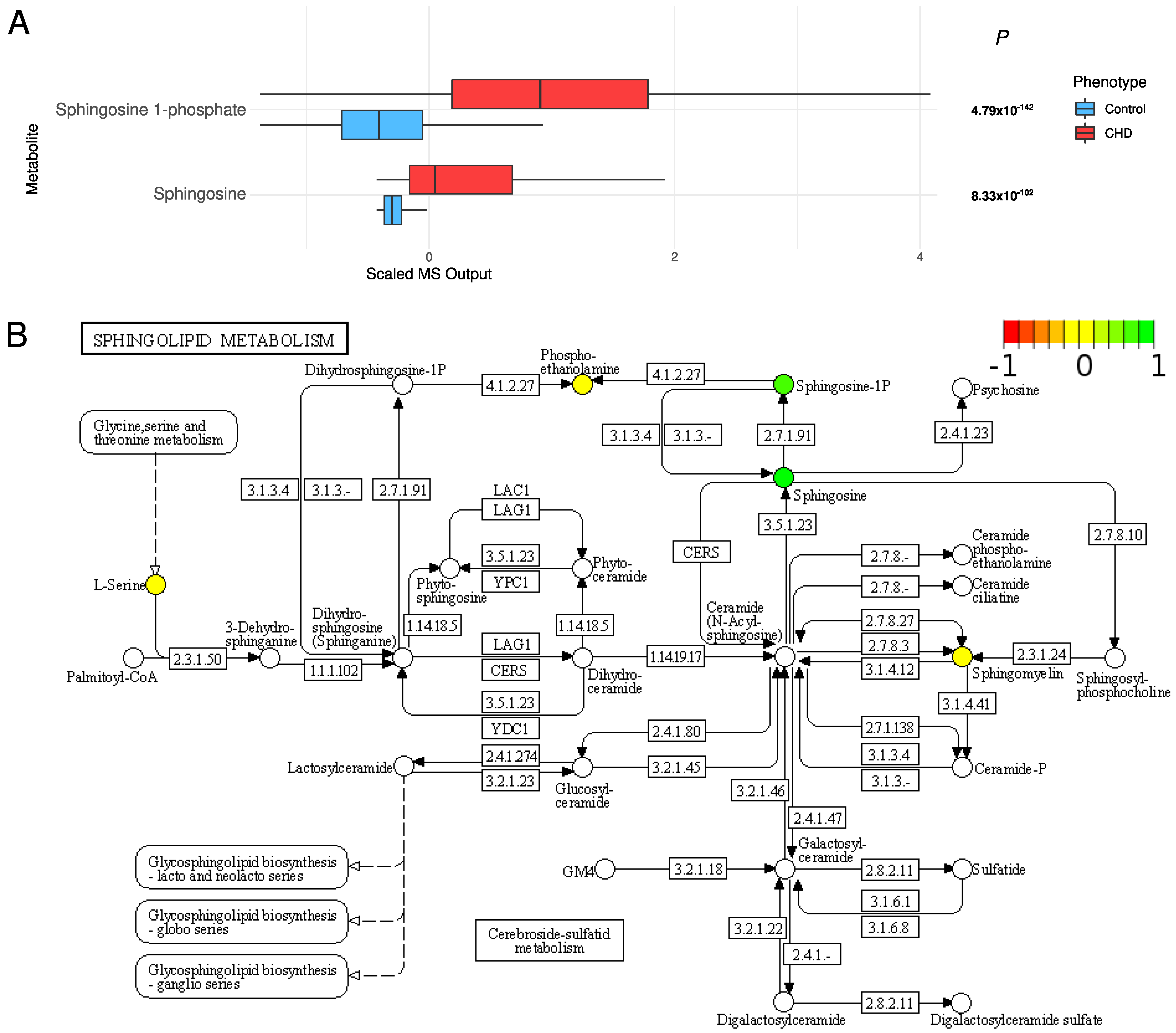
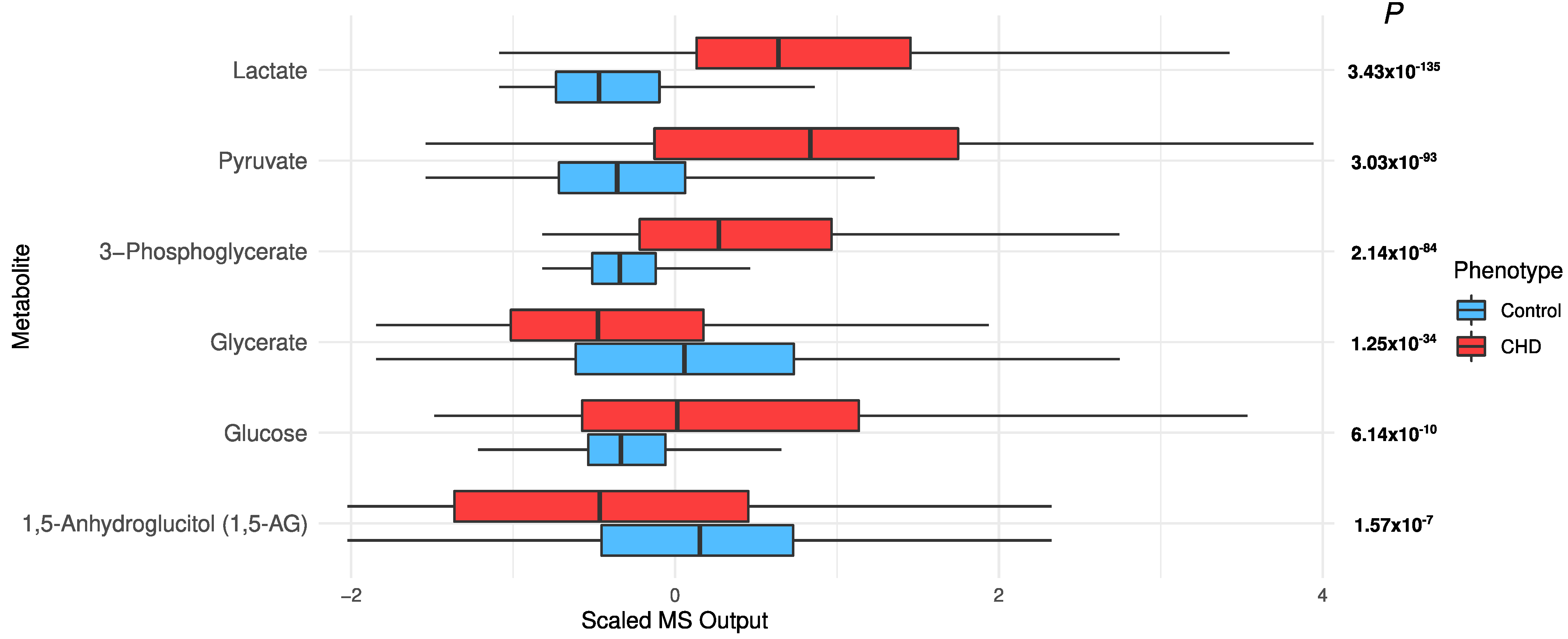
2.4. Changes in Acylcarnitines, Branched Chain Amino Acids, Sphingolipids, and Sugar Metabolisms
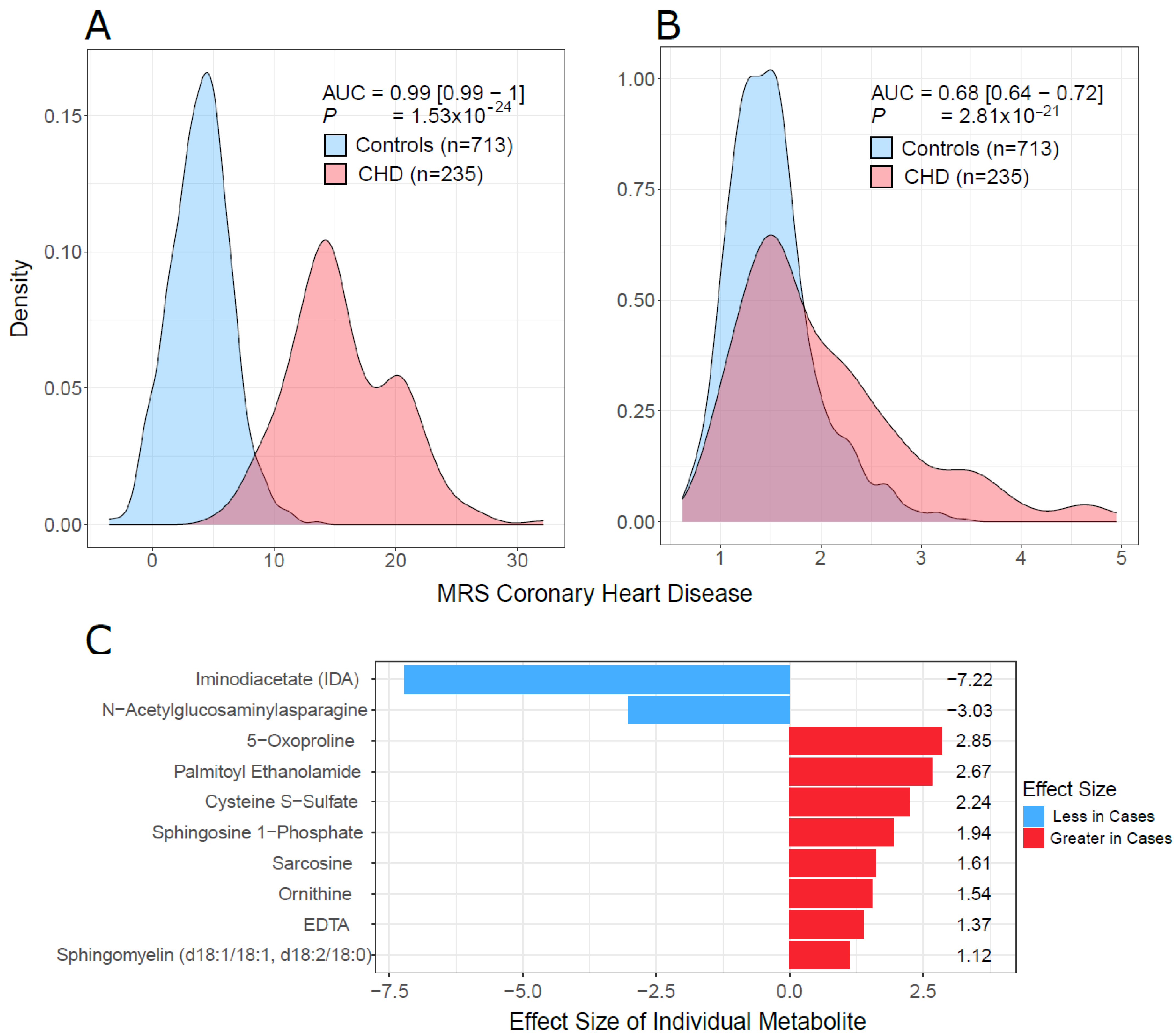
2.5. Multivariate Analysis and Predictive Model
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Metabolomic Profiling and Quality Control
4.3. Univariate Statistical Analysis
4.4. Pathway Enrichment Analysis
4.5. Multivariate Analysis and Predictive Modeling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2021 Update: A Report From the American Heart Association. Circulation 2021, 143, e254–e743. [Google Scholar] [CrossRef]
- Hajar, R. Risk Factors for Coronary Artery Disease: Historical Perspectives. Heart Views 2017, 18, 109–114. [Google Scholar] [CrossRef]
- Menotti, A.; Puddu, P.E. Epidemiology of Heart Disease of Uncertain Etiology: A Population Study and Review of the Problem. Medicina 2019, 55, 687. [Google Scholar] [CrossRef] [Green Version]
- Rana, J.S.; Nieuwdorp, M.; Jukema, J.W.; Kastelein, J.J. Cardiovascular metabolic syndrome—An interplay of, obesity, inflammation, diabetes and coronary heart disease. Diabetes Obes. Metab. 2007, 9, 218–232. [Google Scholar] [CrossRef]
- Shah, S.H.; Hauser, E.R.; Bain, J.R.; Muehlbauer, M.J.; Haynes, C.; Stevens, R.D.; Wenner, B.R.; Dowdy, Z.E.; Granger, C.B.; Ginsburg, G.S.; et al. High heritability of metabolomic profiles in families burdened with premature cardiovascular disease. Mol. Syst. Biol. 2009, 5, 258. [Google Scholar] [CrossRef]
- Kolwicz, S.C., Jr.; Purohit, S.; Tian, R. Cardiac metabolism and its interactions with contraction, growth, and survival of cardiomyocytes. Circ. Res. 2013, 113, 603–616. [Google Scholar] [CrossRef] [Green Version]
- Doenst, T.; Nguyen, T.D.; Abel, E.D. Cardiac metabolism in heart failure: Implications beyond ATP production. Circ. Res. 2013, 113, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Fox, C.S.; Coady, S.; Sorlie, P.D.; D’Agostino, R.B., Sr.; Pencina, M.J.; Vasan, R.S.; Meigs, J.B.; Levy, D.; Savage, P.J. Increasing cardiovascular disease burden due to diabetes mellitus: The Framingham Heart Study. Circulation 2007, 115, 1544–1550. [Google Scholar] [CrossRef]
- Krauss, R.M.; Winston, M.; Fletcher, B.J.; Grundy, S.M. Obesity: Impact on cardiovascular disease. Circulation 1998, 98, 1472–1476. [Google Scholar]
- Shah, S.H.; Sun, J.L.; Stevens, R.D.; Bain, J.R.; Muehlbauer, M.J.; Pieper, K.S.; Haynes, C.; Hauser, E.R.; Kraus, W.E.; Granger, C.B.; et al. Baseline metabolomic profiles predict cardiovascular events in patients at risk for coronary artery disease. Am. Heart J. 2012, 163, 844–850.e1. [Google Scholar] [CrossRef]
- Rizza, S.; Copetti, M.; Rossi, C.; Cianfarani, M.A.; Zucchelli, M.; Luzi, A.; Pecchioli, C.; Porzio, O.; Di Cola, G.; Urbani, A.; et al. Metabolomics signature improves the prediction of cardiovascular events in elderly subjects. Atherosclerosis 2014, 232, 260–264. [Google Scholar] [CrossRef]
- Vaarhorst, A.A.; Verhoeven, A.; Weller, C.M.; Bohringer, S.; Goraler, S.; Meissner, A.; Deelder, A.M.; Henneman, P.; Gorgels, A.P.; van den Brandt, P.A.; et al. A metabolomic profile is associated with the risk of incident coronary heart disease. Am. Heart J. 2014, 168, 45–52.e7. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, C.; Nambi, V.; Morrison, A.C.; Folsom, A.R.; Ballantyne, C.M.; Boerwinkle, E.; Yu, B. Metabolomic Pattern Predicts Incident Coronary Heart Disease. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1475–1482. [Google Scholar] [CrossRef]
- El-Menyar, A.; Al Suwaidi, J.; Badii, R.; Mir, F.; Dalenberg, A.K.; Kullo, I.J. Discovering Novel Biochemical and Genetic Markers for Coronary Heart Disease in Qatari Individuals: The Initiative Qatar Cardiovascular Biorepository. Heart Views 2020, 21, 6–16. [Google Scholar] [CrossRef]
- Salman, A.; Baber, R.; Hannigan, L.; Habermann, J.K.; Henderson, M.K.; Mayrhofer, M.T.; Afifi, N. Qatar Biobank Milestones in Building a Successful Biobank. Biopreserv. Biobank. 2019, 17, 485–486. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.; Kim, O.Y.; Jo, G.; Shin, M.J. Alterations in Circulating Amino Acid Metabolite Ratio Associated with Arginase Activity Are Potential Indicators of Metabolic Syndrome: The Korean Genome and Epidemiology Study. Nutrients 2017, 9, 740. [Google Scholar] [CrossRef] [Green Version]
- Sourij, H.; Meinitzer, A.; Pilz, S.; Grammer, T.B.; Winkelmann, B.R.; Boehm, B.O.; Marz, W. Arginine bioavailability ratios are associated with cardiovascular mortality in patients referred to coronary angiography. Atherosclerosis 2011, 218, 220–225. [Google Scholar] [CrossRef]
- Kovamees, O.; Shemyakin, A.; Pernow, J. Amino acid metabolism reflecting arginase activity is increased in patients with type 2 diabetes and associated with endothelial dysfunction. Diabetes Vasc. Dis. Res. 2016, 13, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Pernow, J.; Jung, C. Arginase as a potential target in the treatment of cardiovascular disease: Reversal of arginine steal? Cardiovasc. Res. 2013, 98, 334–343. [Google Scholar] [CrossRef] [Green Version]
- Durante, W.; Johnson, F.K.; Johnson, R.A. Arginase: A critical regulator of nitric oxide synthesis and vascular function. Clin. Exp. Pharmacol. Physiol. 2007, 34, 906–911. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, R.B.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.W. Arginase: An old enzyme with new tricks. Trends Pharmacol. Sci. 2015, 36, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Romero, M.J.; Platt, D.H.; Tawfik, H.E.; Labazi, M.; El-Remessy, A.B.; Bartoli, M.; Caldwell, R.B.; Caldwell, R.W. Diabetes-induced coronary vascular dysfunction involves increased arginase activity. Circ. Res. 2008, 102, 95–102. [Google Scholar] [CrossRef]
- Pernow, J.; Kiss, A.; Tratsiakovich, Y.; Climent, B. Tissue-specific up-regulation of arginase I and II induced by p38 MAPK mediates endothelial dysfunction in type 1 diabetes mellitus. Br. J. Pharmacol. 2015, 172, 4684–4698. [Google Scholar] [CrossRef]
- Yao, L.; Chandra, S.; Toque, H.A.; Bhatta, A.; Rojas, M.; Caldwell, R.B.; Caldwell, R.W. Prevention of diabetes-induced arginase activation and vascular dysfunction by Rho kinase (ROCK) knockout. Cardiovasc. Res. 2013, 97, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Johnson, F.K.; Johnson, R.A.; Peyton, K.J.; Durante, W. Arginase inhibition restores arteriolar endothelial function in Dahl rats with salt-induced hypertension. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2005, 288, R1057–R1062. [Google Scholar] [CrossRef]
- Cannon, R.O., 3rd. Role of nitric oxide in cardiovascular disease: Focus on the endothelium. Clin. Chem. 1998, 44, 1809–1819. [Google Scholar]
- Shah, S.H.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Crosslin, D.R.; Haynes, C.; Dungan, J.; Newby, L.K.; Hauser, E.R.; Ginsburg, G.S.; et al. Association of a peripheral blood metabolic profile with coronary artery disease and risk of subsequent cardiovascular events. Circ. Cardiovasc. Genet. 2010, 3, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Kalim, S.; Clish, C.B.; Wenger, J.; Elmariah, S.; Yeh, R.W.; Deferio, J.J.; Pierce, K.; Deik, A.; Gerszten, R.E.; Thadhani, R.; et al. A plasma long-chain acylcarnitine predicts cardiovascular mortality in incident dialysis patients. J. Am. Heart Assoc. 2013, 2, e000542. [Google Scholar] [CrossRef] [Green Version]
- Brittain, E.L.; Talati, M.; Fessel, J.P.; Zhu, H.; Penner, N.; Calcutt, M.W.; West, J.D.; Funke, M.; Lewis, G.D.; Gerszten, R.E.; et al. Fatty Acid Metabolic Defects and Right Ventricular Lipotoxicity in Human Pulmonary Arterial Hypertension. Circulation 2016, 133, 1936–1944. [Google Scholar] [CrossRef]
- Koves, T.R.; Ussher, J.R.; Noland, R.C.; Slentz, D.; Mosedale, M.; Ilkayeva, O.; Bain, J.; Stevens, R.; Dyck, J.R.; Newgard, C.B.; et al. Mitochondrial overload and incomplete fatty acid oxidation contribute to skeletal muscle insulin resistance. Cell Metab. 2008, 7, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Schooneman, M.G.; Vaz, F.M.; Houten, S.M.; Soeters, M.R. Acylcarnitines: Reflecting or inflicting insulin resistance? Diabetes 2013, 62, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Szpetnar, M.; Pasternak, K.; Boguszewska, A. Branched chain amino acids (BCAAs) in heart diseases (ischaemic heart disease and myocardial infarction). Ann. Univ. Mariae Curie-Sklodowska Med. 2004, 59, 91–95. [Google Scholar]
- Sun, H.; Wang, Y. Branched chain amino acid metabolic reprogramming in heart failure. Biochim. Biophys. Acta 2016, 1862, 2270–2275. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, F.; Xia, Y.; Zhao, S.; Yan, W.; Wang, H.; Lee, Y.; Li, C.; Zhang, L.; Lian, K.; et al. Defective branched chain amino acid catabolism contributes to cardiac dysfunction and remodeling following myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H1160–H1169. [Google Scholar] [CrossRef]
- Uddin, G.M.; Zhang, L.; Shah, S.; Fukushima, A.; Wagg, C.S.; Gopal, K.; Al Batran, R.; Pherwani, S.; Ho, K.L.; Boisvenue, J.; et al. Impaired branched chain amino acid oxidation contributes to cardiac insulin resistance in heart failure. Cardiovasc. Diabetol. 2019, 18, 86. [Google Scholar] [CrossRef]
- Poss, A.M.; Maschek, J.A.; Cox, J.E.; Hauner, B.J.; Hopkins, P.N.; Hunt, S.C.; Holland, W.L.; Summers, S.A.; Playdon, M.C. Machine learning reveals serum sphingolipids as cholesterol-independent biomarkers of coronary artery disease. J. Clin. Investig. 2020, 130, 1363–1376. [Google Scholar] [CrossRef] [Green Version]
- Lim, G.B. Sphingolipids are biomarkers of coronary disease. Nat. Rev. Cardiol. 2020, 17, 200. [Google Scholar] [CrossRef]
- Borodzicz, S.; Czarzasta, K.; Kuch, M.; Cudnoch-Jedrzejewska, A. Sphingolipids in cardiovascular diseases and metabolic disorders. Lipids Health Dis. 2015, 14, 55. [Google Scholar] [CrossRef] [Green Version]
- Kovilakath, A.; Jamil, M.; Cowart, L.A. Sphingolipids in the Heart: From Cradle to Grave. Front. Endocrinol. 2020, 11, 652. [Google Scholar] [CrossRef]
- Wende, A.R.; Brahma, M.K.; McGinnis, G.R.; Young, M.E. Metabolic Origins of Heart Failure. JACC Basic Transl. Sci. 2017, 2, 297–310. [Google Scholar] [CrossRef]
- Shorewala, V. Early detection of coronary heart disease using ensemble techniques. Inform. Med. Unlocked 2021, 26, 100655. [Google Scholar]
- Wei, R.; Wang, J.; Su, M.; Jia, E.; Chen, S.; Chen, T.; Ni, Y. Missing Value Imputation Approach for Mass Spectrometry-based Metabolomics Data. Sci. Rep. 2018, 8, 663. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.; Chong, J.; Li, S.; Xia, J. MetaboAnalystR 3.0: Toward an Optimized Workflow for Global Metabolomics. Metabolites 2020, 10, 186. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. MSEA: A web-based tool to identify biologically meaningful patterns in quantitative metabolomic data. Nucleic Acids Res. 2010, 38, W71–W77. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and Regression by RandomForest. Forest 2001, 23. [Google Scholar]
- Kuhn, M. Caret: Classification and Regression Training. R Package Version 6 2021. [Google Scholar]
- Sing, T.; Sander, O.; Beerenwinkel, N.; Lengauer, T. ROCR: Visualizing classifier performance in R. Bioinformatics 2005, 21, 3940–3941. [Google Scholar] [CrossRef]
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Pathway | C | H | ER | Rank | Pathway | C | H | ER |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Malate-Aspartate Shuttle | 10 | 3 | 937.76 | 24 | Oxidation of Branched Chain Fatty Acids | 26 | 4 | 429.34 |
| 2 | D-Arginine and D-Ornithine Metabolism | 11 | 3 | 760 | 25 | Mitochondrial Electron Transport Chain | 19 | 4 | 422.54 |
| 3 | Glucose-Alanine Cycle | 13 | 5 | 740.04 | 26 | Propanoate Metabolism | 42 | 5 | 419.62 |
| 4 | Gluconeogenesis | 35 | 6 | 730.95 | 27 | Tryptophan Metabolism | 60 | 12 | 386.48 |
| 5 | Warburg Effect | 58 | 12 | 695.11 | 28 | Ammonia Recycling | 32 | 12 | 386.28 |
| 6 | Citric Acid Cycle | 32 | 7 | 683.41 | 29 | Glycine and Serine Metabolism | 59 | 21 | 379.37 |
| 7 | Glutathione Metabolism | 21 | 6 | 659.62 | 30 | Nicotinate and Nicotinamide Metabolism | 37 | 9 | 363.32 |
| 8 | Lysine Degradation | 30 | 4 | 638.04 | 31 | Carnitine Synthesis | 22 | 8 | 346.34 |
| 9 | Pyruvate Metabolism | 48 | 5 | 620.85 | 32 | Betaine Metabolism | 21 | 6 | 342.96 |
| 10 | Tyrosine Metabolism | 72 | 9 | 608.32 | 33 | Sphingolipid Metabolism | 40 | 10 | 341.81 |
| 11 | Urea Cycle | 29 | 12 | 578.21 | 34 | Valine, Leucine and Isoleucine Degradation | 60 | 8 | 328.06 |
| 12 | Transfer of Acetyl Groups into Mitochondria | 22 | 5 | 573.51 | 35 | Histidine Metabolism | 43 | 10 | 305.92 |
| 13 | Glycolysis | 25 | 4 | 572.2 | 36 | Arachidonic Acid Metabolism | 69 | 4 | 288.72 |
| 14 | Phytanic Acid Peroxisomal Oxidation | 26 | 2 | 571.65 | 37 | Starch and Sucrose Metabolism | 31 | 5 | 265.43 |
| 15 | Phenylalanine and Tyrosine Metabolism | 28 | 7 | 544.89 | 38 | Steroidogenesis | 43 | 3 | 261.33 |
| 16 | Alanine Metabolism | 17 | 7 | 531.58 | 39 | Methionine Metabolism | 43 | 15 | 243.09 |
| 17 | Cysteine Metabolism | 26 | 6 | 514.23 | 40 | Glycerolipid Metabolism | 25 | 6 | 240.15 |
| 18 | Amino Sugar Metabolism | 33 | 6 | 459.18 | 41 | Pyrimidine Metabolism | 59 | 9 | 155.08 |
| 19 | Glutamate Metabolism | 49 | 12 | 457.04 | 42 | Fatty Acid Biosynthesis | 35 | 8 | 133.24 |
| 20 | Arginine and Proline Metabolism | 53 | 16 | 441.27 | 43 | Bile Acid Biosynthesis | 65 | 11 | 107.6 |
| 21 | Beta-Alanine Metabolism | 34 | 8 | 441.12 | 44 | Galactose Metabolism | 38 | 7 | 133.77 |
| 22 | Aspartate Metabolism | 35 | 12 | 438.07 | 45 | Phosphatidylcholine Biosynthesis | 14 | 4 | 359.58 |
| 23 | Purine Metabolism | 74 | 12 | 433.8 |
| CHD | Controls | p * | |||||
|---|---|---|---|---|---|---|---|
| Females | Males | All | Females | Males | All | ||
| Participants, N (%) | 600 (61.9) | 370 (38.1) | 970 (100) | 1505 (50.5) | 1478 (49.6) | 2983 (100) | 7.9 × 10−10 |
| Age, Mean (SD) years | 53.4 (14.5) | 51.6 (14.9) | 52.7 (14.6) | 39.6 (11.4) | 40.1 (12.6) | 39.8 (12.0) | <2.2 × 10−16 |
| BMI, Mean (SD) kg.m−2 | 29.8 (5.0) | 31.8 (6.2) | 30.5 (5.5) | 28.6 (5.5) | 29.4 (6.3) | 29.0 (5.9) | 2.7 × 10−13 |
| Type 2 Diabetes #, N (%) | 302 (62.8) | 179 (37.2) | 481 (100) | 135 (49.6) | 137 (50.4) | 272 (100) | 5.9 × 10−4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, E.; El-Menyar, A.; Kunji, K.; Elsousy, R.; Mokhtar, H.R.B.; Ahmad, E.; Al-Nesf, M.; Beotra, A.; Al-Maadheed, M.; Mohamed-Ali, V.; et al. Untargeted Metabolomics Profiling Reveals Perturbations in Arginine-NO Metabolism in Middle Eastern Patients with Coronary Heart Disease. Metabolites 2022, 12, 517. https://doi.org/10.3390/metabo12060517
Ullah E, El-Menyar A, Kunji K, Elsousy R, Mokhtar HRB, Ahmad E, Al-Nesf M, Beotra A, Al-Maadheed M, Mohamed-Ali V, et al. Untargeted Metabolomics Profiling Reveals Perturbations in Arginine-NO Metabolism in Middle Eastern Patients with Coronary Heart Disease. Metabolites. 2022; 12(6):517. https://doi.org/10.3390/metabo12060517
Chicago/Turabian StyleUllah, Ehsan, Ayman El-Menyar, Khalid Kunji, Reem Elsousy, Haira R. B. Mokhtar, Eiman Ahmad, Maryam Al-Nesf, Alka Beotra, Mohammed Al-Maadheed, Vidya Mohamed-Ali, and et al. 2022. "Untargeted Metabolomics Profiling Reveals Perturbations in Arginine-NO Metabolism in Middle Eastern Patients with Coronary Heart Disease" Metabolites 12, no. 6: 517. https://doi.org/10.3390/metabo12060517