
Proteobacteria and Firmicutes Secreted Factors Exert Distinct Effects on Pseudomonas aeruginosa Infection under Normoxia or Mild Hypoxia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
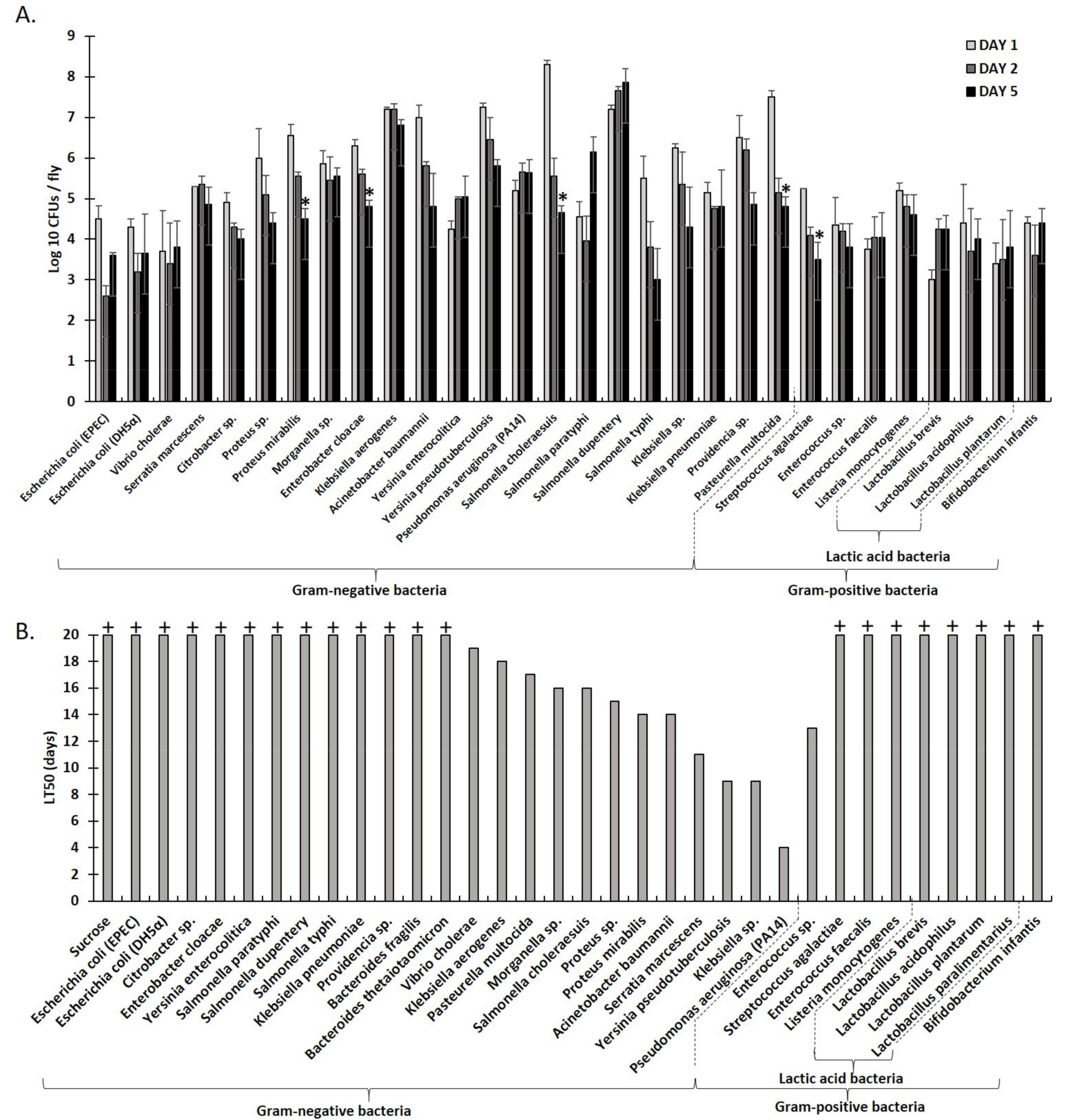
2.1. An Array of Culturable Strains of Bacteria Found in Humans Can Stably Associate with Drosophila via Feeding
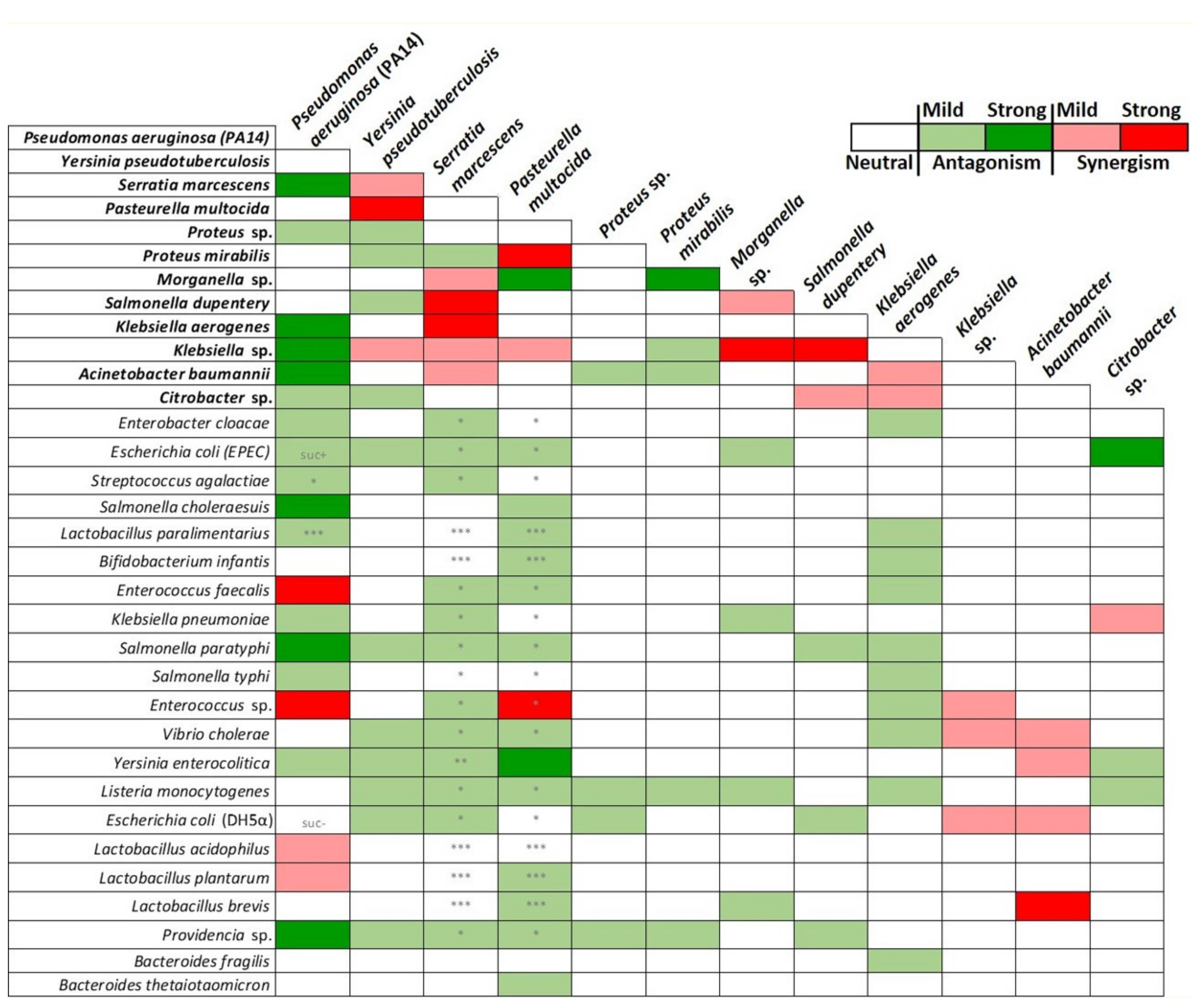
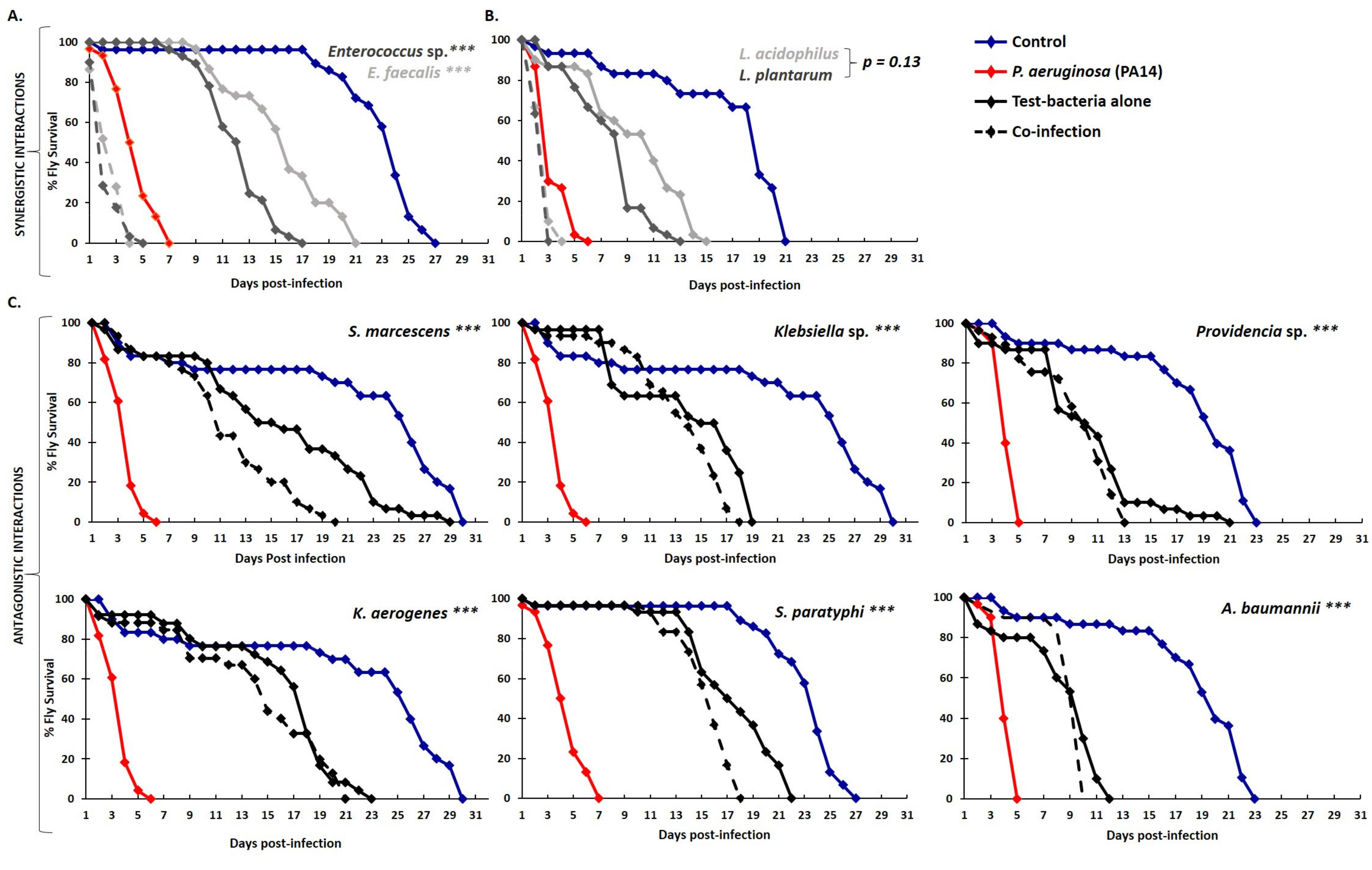
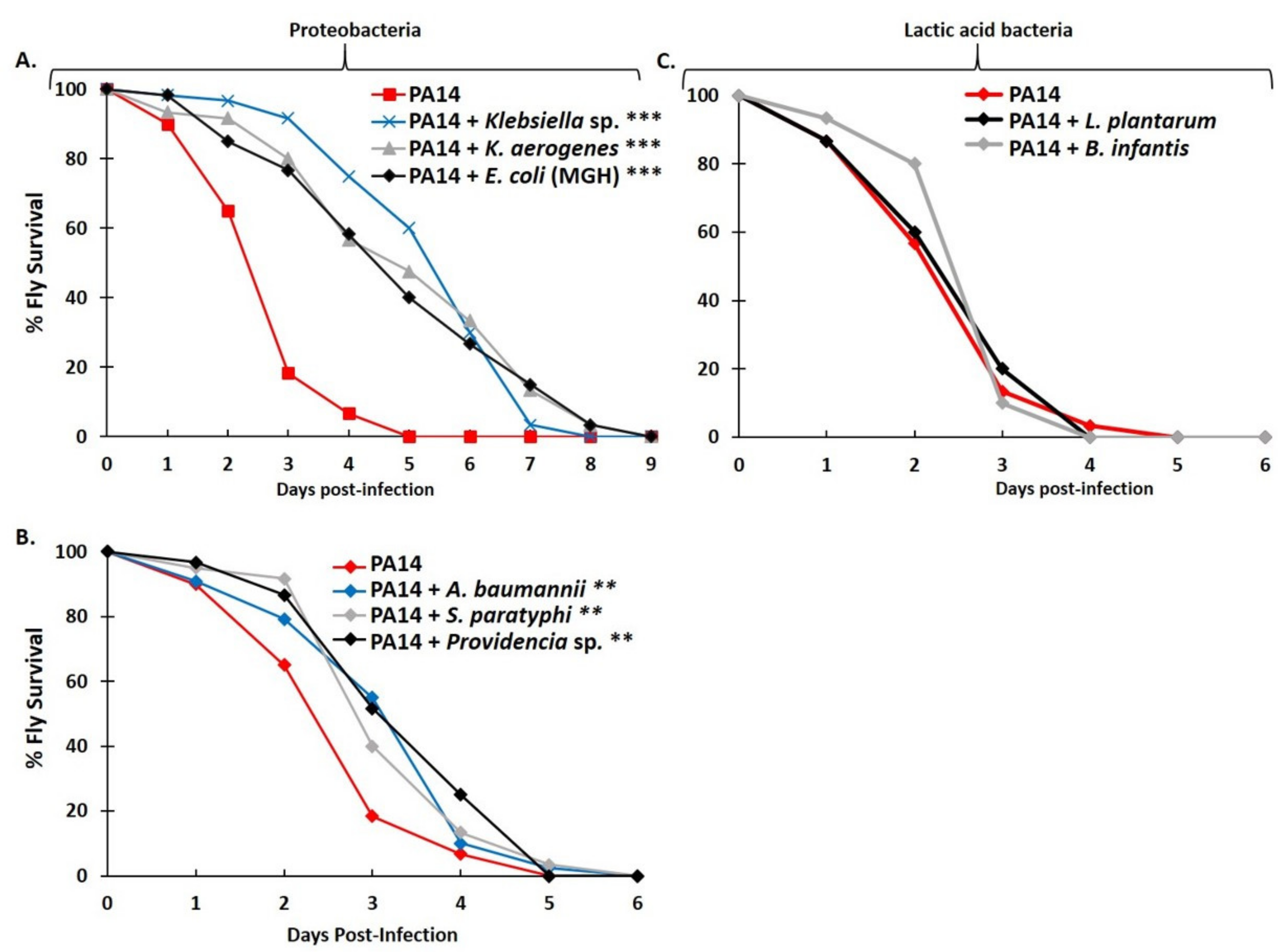
2.2. Graded Classification of Bacterial Interactions as Neutral, Synergistic, or Antagonistic, Based on Drosophila Survival Rate upon Feeding Infection
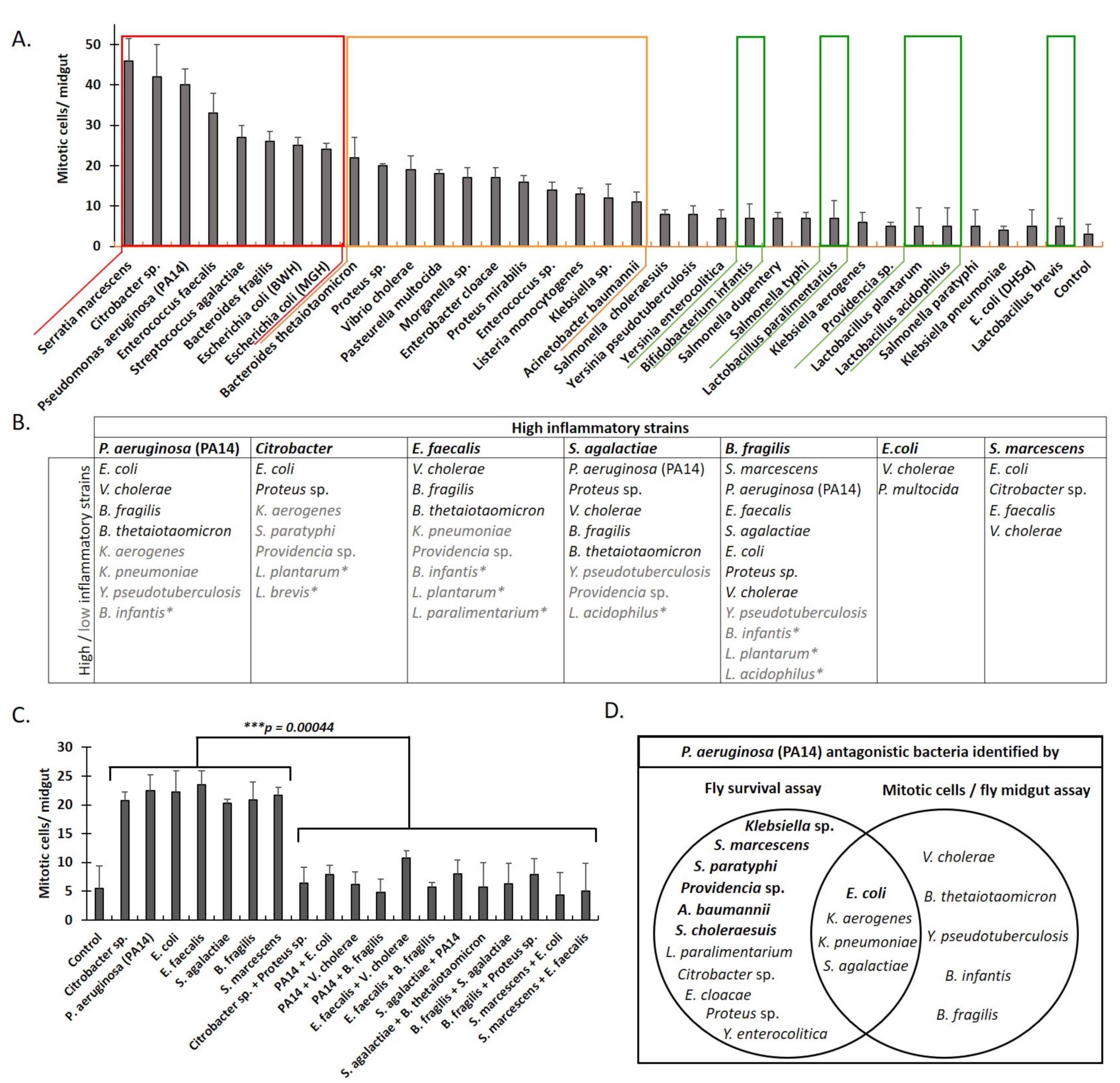
2.3. Classification of Interactions between Pathogenic Bacteria as Neutral, Synergistic, or Antagonistic, Based on Drosophila Midgut Cell Mitosis upon Feeding Infection
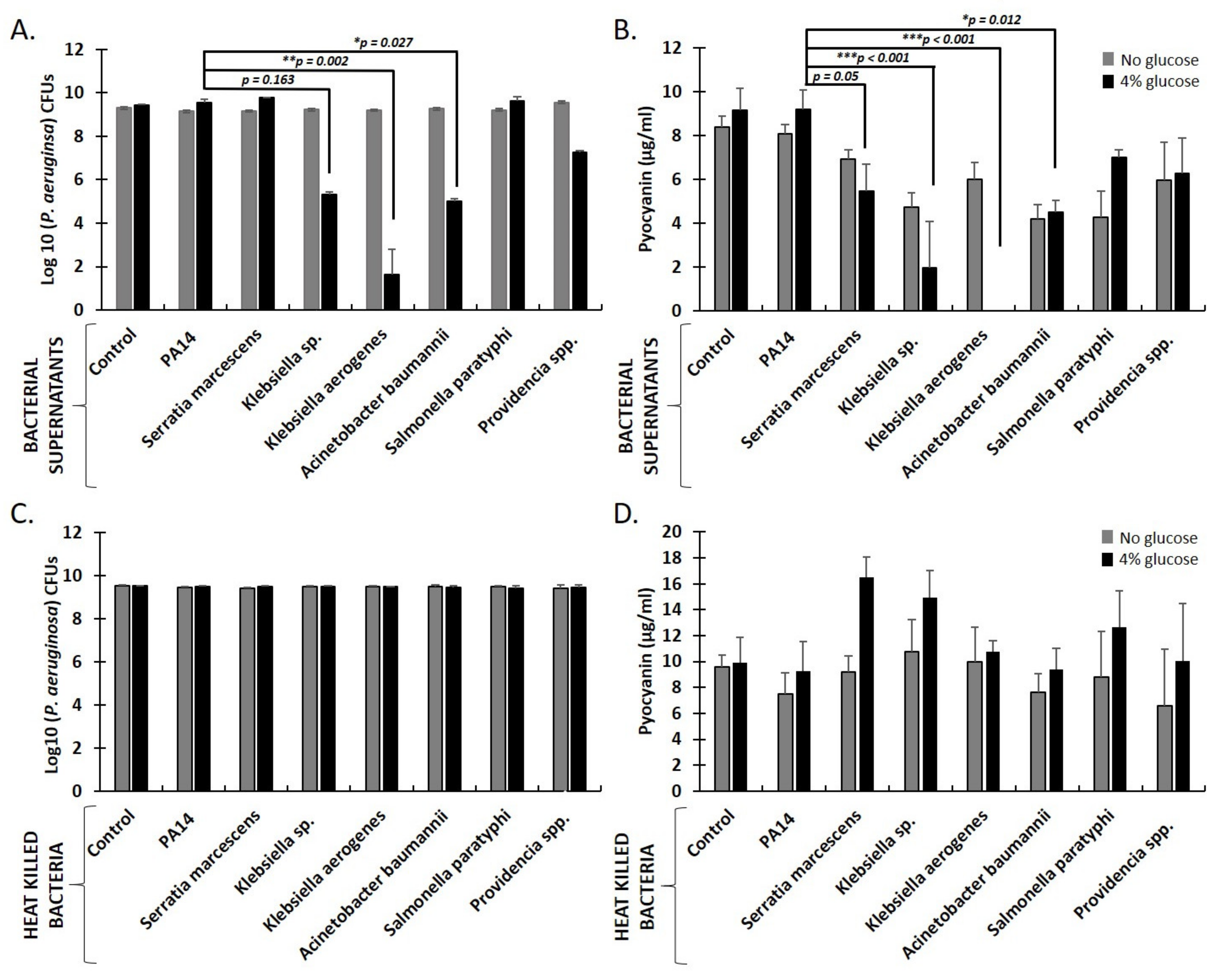
2.4. Secreted Factors from K. aerogenes, Klebsiella sp., and A. baumanni Inhibit P. aeruginosa Growth and Pyocyanin Production in Aerobic Liquid Cultures Supplemented with Sugars
2.5. Antagonistic Interactions between P. aeruginosa and Selected Proteobacteria but Not Lactic Acid Bacteria Also Occur under Mild Hypoxia
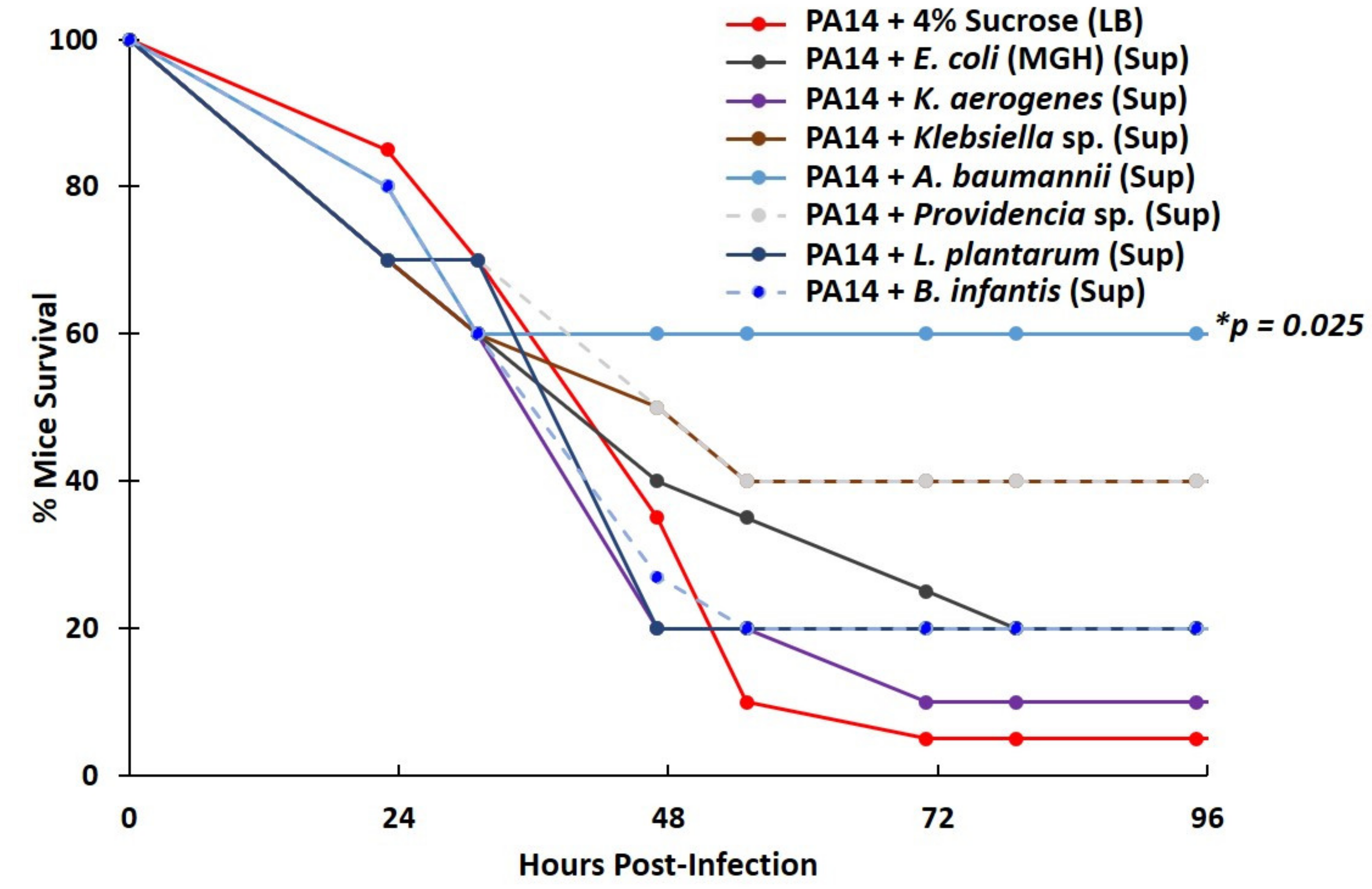
2.6. Lactic Acid Bacteria Supernatants Produced under Severe Hypoxia Fail to Inhibit P. aeruginosa Virulence in a Mouse Lung-Infection Model
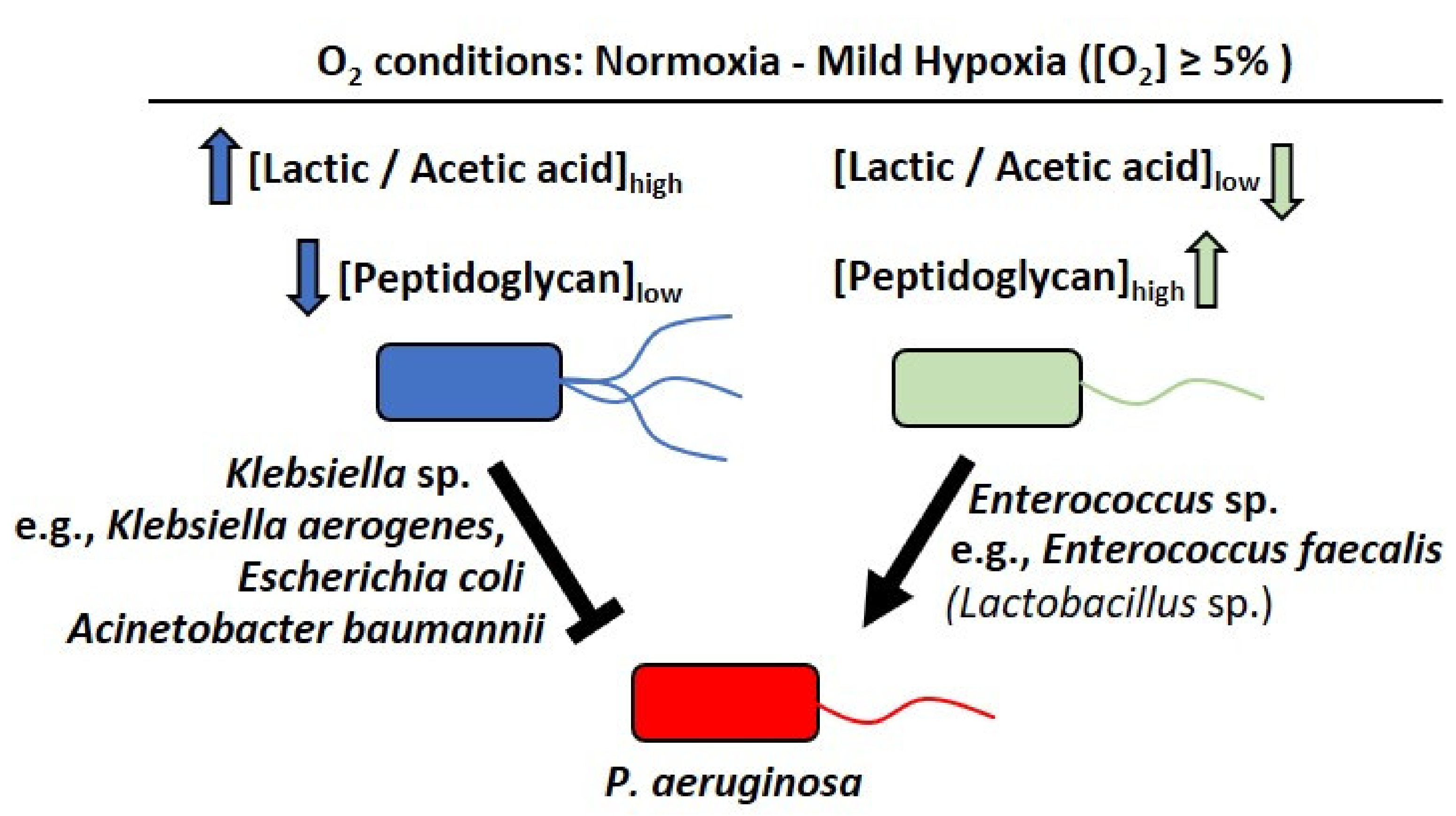
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.1.1. LB/BHI Broth Media Preparation
4.1.2. LB Agar Plates/LB Agar + Rifampicin Plates Preparation
4.2. Bacterial Strains
4.3. Drosophila Diet, Maintenance, and Experiments
4.3.1. Fly infection and Survival
4.3.2. Fly Intestinal Colonization Assay
4.3.3. Mitotic Cells Quantification
4.3.4. Assessing PA14 Growth—CFUs (Colony-Forming Units) Assay
4.3.5. PA14 Pyocyanin Measurement
4.4. Mice Diet, Maintenance and Experiments
Intranasal Mouse Lung Infection Assay
4.5. Computational Analysis
4.6. Ethics Issues
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rice, L.B. Federal Funding for the Study of Antimicrobial Resistance in Nosocomial Pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef]
- Allegranzi, B. Burden of Endemic Health-Care-Associated Infection in Developing Countries: Systematic Review and Meta-Analysis. Lancet 2011, 377, 228–241. [Google Scholar] [CrossRef]
- Ibrahim, M.E.; Bilal, N.E.; Hamid, M.E. Increased Multi-Drug Resistant Escherichia Coli from Hospitals in Khartoum State, Sudan. Afr. Health Sci. 2012, 12, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical Relevance of the ESKAPE Pathogens. Expert Rev. Anti-Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Navidinia, M. The Clinical Importance of Emerging ESKAPE Pathogens in Nosocomial Infections. Arch. Adv. Biosci. 2008, 7, 8–12. [Google Scholar]
- Santajit, S.; Indrawattana, N. Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, K. Persister Cells, Dormancy and Infectious Disease. Nat. Rev. Microbiol. 2007, 5, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Panayidou, S.; Ioannidou, E.; Apidianakis, Y. Human Pathogenic Bacteria, Fungi, and Viruses in Drosophila: Disease Modeling, Lessons, and Shortcomings. Virulence 2014, 5, 253–269. [Google Scholar] [CrossRef] [Green Version]
- Kapsetaki, S.-E.; Tzelepis, I.; Avgousti, K.; Livadaras, I.; Garantonakis, N.; Varikou, K.; Apidianakis, Y. The Bacterial Metabolite 2-Aminoacetophenone Promotes Association of Pathogenic Bacteria with Flies. Nat. Commun. 2014, 5, 4401. [Google Scholar] [CrossRef] [Green Version]
- Tzelepis, I.; Kapsetaki, S.-E.; Panayidou, S.; Apidianakis, Y. Drosophila melanogaster: A First Step and a Stepping-Stone to Anti-Infectives. Curr. Opin. Pharmacol. 2013, 13, 763–768. [Google Scholar] [CrossRef]
- Fallon, P.G.; Allen, R.L.; Rich, T. Primitive Toll Signalling: Bugs, Flies, Worms and Man. Trends Immunol. 2001, 22, 63–66. [Google Scholar] [CrossRef]
- Medzhitov, R.; Preston-Hurlburt, P.; Janeway, C.A.J. A Human Homologue of the Drosophila Toll Protein Signals Activation of Adaptive Immunity. Nature 1997, 388, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.A.; Reichhart, J.-M. Drosophila Innate Immunity: An Evolutionary Perspective. Nat. Immunol. 2002, 3, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Apidianakis, Y.; Pitsouli, C.; Perrimon, N.; Rahme, L. Synergy between Bacterial Infection and Genetic Predisposition in Intestinal Dysplasia. Proc. Natl. Acad. Sci. USA 2009, 106, 20883–20888. [Google Scholar] [CrossRef] [Green Version]
- Galko, M.J.; Krasnow, M.A. Cellular and Genetic Analysis of Wound Healing in Drosophila Larvae. PLoS Biol. 2004, 2, E239. [Google Scholar] [CrossRef] [Green Version]
- Goto, A.; Kadowaki, T.; Kitagawa, Y. Drosophila Hemolectin Gene Is Expressed in Embryonic and Larval Hemocytes and Its Knock down Causes Bleeding Defects. Dev. Biol. 2003, 264, 582–591. [Google Scholar] [CrossRef] [Green Version]
- Scherfer, C.; Qazi, M.R.; Takahashi, K.; Ueda, R.; Dushay, M.S.; Theopold, U.; Lemaitre, B. The Toll Immune-Regulated Drosophila Protein Fondue Is Involved in Hemolymph Clotting and Puparium Formation. Dev. Biol. 2006, 295, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Scherfer, C.; Karlsson, C.; Loseva, O.; Bidla, G.; Goto, A.; Havemann, J.; Dushay, M.S.; Theopold, U. Isolation and Characterization of Hemolymph Clotting Factors in Drosophila Melanogaster by a Pullout Method. Curr. Biol. 2004, 14, 625–629. [Google Scholar] [CrossRef] [Green Version]
- Dickson, R.P.; Erb-Downward, J.R.; Huffnagle, G.B. The Role of the Bacterial Microbiome in Lung Disease. Expert Rev. Respir. Med. 2013, 7, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The Human Skin Microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Zheng, L.; Kelly, C.J.; Colgan, S.P. Physiologic Hypoxia and Oxygen Homeostasis in the Healthy Intestine. A Review in the Theme: Cellular Responses to Hypoxia. Am. J. Physiol. Cell Physiol. 2015, 309, C350–C360. [Google Scholar] [CrossRef] [Green Version]
- Dalton, T.; Dowd, S.E.; Wolcott, R.D.; Sun, Y.; Watters, C.; Griswold, J.A.; Rumbaugh, K.P. An in Vivo Polymicrobial Biofilm Wound Infection Model to Study Interspecies Interactions. PLoS ONE 2011, 6, e27317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gjødsbøl, K.; Christensen, J.J.; Karlsmark, T.; Jørgensen, B.; Klein, B.M.; Krogfelt, K.A. Multiple Bacterial Species Reside in Chronic Wounds: A Longitudinal Study. Int. Wound J. 2006, 3, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Halbert, A.R.; Stacey, M.C.; Rohr, J.B.; Jopp-McKay, A. The Effect of Bacterial Colonization on Venous Ulcer Healing. Australas J. Derm. 1992, 33, 75–80. [Google Scholar] [CrossRef]
- Duan, K.; Dammel, C.; Stein, J.; Rabin, H.; Surette, M.G. Modulation of Pseudomonas Aeruginosa Gene Expression by Host Microflora through Interspecies Communication. Mol. Microbiol. 2003, 50, 1477–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibley, C.D.; Duan, K.; Fischer, C.; Parkins, M.D.; Storey, D.G.; Rabin, H.R.; Surette, M.G. Discerning the Complexity of Community Interactions Using a Drosophila Model of Polymicrobial Infections. PLoS Pathog. 2008, 4, e1000184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korgaonkar, A.; Trivedi, U.; Rumbaugh, K.P.; Whiteley, M. Community Surveillance Enhances Pseudomonas Aeruginosa Virulence during Polymicrobial Infection. Proc. Natl. Acad. Sci. USA 2013, 110, 1059–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoetendal, E.G.; Raes, J.; van den Bogert, B.; Arumugam, M.; Booijink, C.C.G.M.; Troost, F.J.; Bork, P.; Wels, M.; de Vos, W.M.; Kleerebezem, M. The Human Small Intestinal Microbiota Is Driven by Rapid Uptake and Conversion of Simple Carbohydrates. ISME J. 2012, 6, 1415–1426. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Macfarlane, S. Fermentation in the Human Large Intestine: Its Physiologic Consequences and the Potential Contribution of Prebiotics. J. Clin. Gastroenterol. 2011, 45, S120–S127. [Google Scholar] [CrossRef] [Green Version]
- Christofi, T.; Panayidou, S.; Dieronitou, I.; Michael, C.; Apidianakis, Y. Metabolic Output Defines Escherichia Coli as a Health-Promoting Microbe against Intestinal Pseudomonas Aeruginosa. Sci. Rep. 2019, 9, 14463. [Google Scholar] [CrossRef] [Green Version]
- Korgaonkar, A.K.; Whiteley, M. Pseudomonas Aeruginosa Enhances Production of an Antimicrobial in Response to N-Acetylglucosamine and Peptidoglycan. J. Bacteriol. 2011, 193, 909–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radtke, F.; Clevers, H. Self-Renewal and Cancer of the Gut: Two Sides of a Coin. Science 2005, 307, 1904–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panayidou, S.; Apidianakis, Y. Regenerative Inflammation: Lessons from Drosophila Intestinal Epithelium in Health and Disease. Pathogens 2013, 2, 209–231. [Google Scholar] [CrossRef] [Green Version]
- Karin, M.; Clevers, H. Reparative Inflammation Takes Charge of Tissue Regeneration. Nature 2016, 529, 307–315. [Google Scholar] [CrossRef]
- Tamamouna, V.; Panagi, M.; Theophanous, A.; Demosthenous, M.; Michail, M.; Papadopoulou, M.; Teloni, S.; Pitsouli, C.; Apidianakis, Y. Evidence of Two Types of Balance between Stem Cell Mitosis and Enterocyte Nucleus Growth in the Drosophila Midgut. Development 2020, 147, dev189472. [Google Scholar] [CrossRef] [PubMed]
- Chichlowski, M.; Hale, L.P. Bacterial-Mucosal Interactions in Inflammatory Bowel Disease: An Alliance Gone Bad. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1139–G1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, A.; Hoffmann, J.A.; Meister, M. Analysis of the Drosophila Host Defense in Domino Mutant Larvae, Which Are Devoid of Hemocytes. Proc. Natl. Acad. Sci. USA 1998, 95, 14337–14342. [Google Scholar] [CrossRef] [Green Version]
- Storelli, G.; Defaye, A.; Erkosar, B.; Hols, P.; Royet, J.; Leulier, F. Lactobacillus Plantarum Promotes Drosophila Systemic Growth by Modulating Hormonal Signals through TOR-Dependent Nutrient Sensing. Cell Metab. 2011, 14, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Pais, I.S.; Valente, R.S.; Sporniak, M.; Teixeira, L. Drosophila melanogaster Establishes a Species-Specific Mutualistic Interaction with Stable Gut-Colonizing Bacteria. PLoS Biol. 2018, 16, e2005710. [Google Scholar] [CrossRef] [Green Version]
- Mariam, S.H.; Zegeye, N.; Aseffa, A.; Howe, R. Diffusible Substances from Lactic Acid Bacterial Cultures Exert Strong Inhibitory Effects on Listeria Monocytogenes and Salmonella Enterica Serovar Enteritidis in a Co-Culture Model. BMC Microbiol. 2017, 17, 35. [Google Scholar] [CrossRef] [Green Version]
- Melo, T.A.; dos Santos, T.F.; de Almeida, M.E.; Junior, L.A.G.F.; Andrade, E.F.; Rezende, R.P.; Marques, L.M.; Romano, C.C. Inhibition of Staphylococcus Aureus Biofilm by Lactobacillus Isolated from Fine Cocoa. BMC Microbiol. 2016, 16, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Deng, X.; Zhou, X.; Hao, Y.; Li, Y. Influence of Helicobacter Pylori Culture Supernatant on the Ecological Balance of a Dual-Species Oral Biofilm. J. Appl. Oral Sci. 2018, 26, e20170113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, A.; Zhu, J.; Ye, X.; Ge, Y.; Li, J. Inhibition of Biofilm Development and Spoilage Potential of Shewanella Baltica by Quorum Sensing Signal in Cell-Free Supernatant from Pseudomonas Fluorescens. Int. J. Food Microbiol. 2016, 230, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Amel, A.M.; Farida, B.; Djamila, S. Anti-Adherence Potential of Enterococcus Durans Cells and Its Cell-Free Supernatant on Plastic and Stainless Steel against Foodborne Pathogens. Folia Microbiol. 2015, 60, 357–363. [Google Scholar] [CrossRef]
- Schoster, A.; Kokotovic, B.; Permin, A.; Pedersen, P.D.; Dal Bello, F.; Guardabassi, L. In Vitro Inhibition of Clostridium Difficile and Clostridium Perfringens by Commercial Probiotic Strains. Anaerobe 2013, 20, 36–41. [Google Scholar] [CrossRef]
- Chorianopoulos, N.G.; Giaouris, E.D.; Kourkoutas, Y.; Nychas, G.-J.E. Inhibition of the Early Stage of Salmonella Enterica Serovar Enteritidis Biofilm Development on Stainless Steel by Cell-Free Supernatant of a Hafnia Alvei Culture. Appl. Environ. Microbiol. 2010, 76, 2018–2022. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-S.; Wu, H.-C.; Kuo, C.-Y.; Chen, Y.-W.; Ho, S.; Yanagida, F. Leucocin C-607, a Novel Bacteriocin from the Multiple-Bacteriocin-Producing Leuconostoc Pseudomesenteroides 607 Isolated from Persimmon. Probiotics Antimicrob. Proteins 2018, 10, 148–156. [Google Scholar] [CrossRef]
- Kim, D.-H.; Jeong, D.; Song, K.-Y.; Kang, I.-B.; Kim, H.; Seo, K.-H. Culture Supernatant Produced by Lactobacillus Kefiri from Kefir Inhibits the Growth of Cronobacter Sakazakii. J. Dairy Res. 2018, 85, 98–103. [Google Scholar] [CrossRef]
- Pridmore, R.D.; Pittet, A.-C.; Praplan, F.; Cavadini, C. Hydrogen Peroxide Production by Lactobacillus Johnsonii NCC 533 and Its Role in Anti-Salmonella Activity. FEMS Microbiol. Lett. 2008, 283, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Sambanthamoorthy, K.; Feng, X.; Patel, R.; Patel, S.; Paranavitana, C. Antimicrobial and Antibiofilm Potential of Biosurfactants Isolated from Lactobacilli against Multi-Drug-Resistant Pathogens. BMC Microbiol. 2014, 14, 197. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Lim, J.J.; Kim, D.G.; Simborio, H.L.; Kim, D.H.; Reyes, A.W.B.; Min, W.; Lee, H.J.; Kim, D.H.; Chang, H.H.; et al. Characterization of Culture Supernatant Proteins from Brucella Abortus and Its Protection Effects against Murine Brucellosis. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Makino, S.; Ikegami, S.; Kano, H.; Sashihara, T.; Sugano, H.; Horiuchi, H.; Saito, T.; Oda, M. Immunomodulatory Effects of Polysaccharides Produced by Lactobacillus Delbrueckii Ssp. Bulgaricus OLL1073R-1. J. Dairy Sci. 2006, 89, 2873–2881. [Google Scholar] [CrossRef] [Green Version]
- Bermudez-Brito, M.; Muñoz-Quezada, S.; Gomez-Llorente, C.; Romero, F.; Gil, A. Lactobacillus Rhamnosus and Its Cell-Free Culture Supernatant Differentially Modulate Inflammatory Biomarkers in Escherichia Coli-Challenged Human Dendritic Cells. Br. J. Nutr. 2014, 111, 1727–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catterson, J.H.; Knowles-Barley, S.; James, K.; Heck, M.M.S.; Harmar, A.J.; Hartley, P.S. Dietary Modulation of Drosophila Sleep-Wake Behaviour. PLoS ONE 2010, 5, e12062. [Google Scholar] [CrossRef] [Green Version]
- Galenza, A.; Hutchinson, J.; Campbell, S.D.; Hazes, B.; Foley, E. Glucose Modulates Drosophila Longevity and Immunity Independent of the Microbiota. Biol. Open 2016, 5, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Na, J.; Musselman, L.P.; Pendse, J.; Baranski, T.J.; Bodmer, R.; Ocorr, K.; Cagan, R. A Drosophila Model of High Sugar Diet-Induced Cardiomyopathy. PLoS Genet. 2013, 9, e1003175. [Google Scholar] [CrossRef] [Green Version]
- Morris, S.N.S.; Coogan, C.; Chamseddin, K.; Fernandez-Kim, S.O.; Kolli, S.; Keller, J.N.; Bauer, J.H. Development of Diet-Induced Insulin Resistance in Adult Drosophila melanogaster. Biochim. Biophys. Acta 2012, 1822, 1230–1237. [Google Scholar] [CrossRef] [Green Version]
- Singhal, R.; Shah, Y.M. Oxygen Battle in the Gut: Hypoxia and Hypoxia-Inducible Factors in Metabolic and Inflammatory Responses in the Intestine. J. Biol. Chem. 2020, 295, 10493–10505. [Google Scholar] [CrossRef]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut Biogeography of the Bacterial Microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Xue, J.; Lai, J.C.K.; Schork, N.J.; White, K.P.; Haddad, G.G. Mechanisms Underlying Hypoxia Tolerance in Drosophila melanogaster: Hairy as a Metabolic Switch. PLoS Genet. 2008, 4, e1000221. [Google Scholar] [CrossRef] [Green Version]
- Maresca, D.; Zotta, T.; Mauriello, G. Adaptation to Aerobic Environment of Lactobacillus Johnsonii/Gasseri Strains. Front. Microbiol. 2018, 9, 157. [Google Scholar] [CrossRef] [PubMed]
- Short, F.L.; Murdoch, S.L.; Ryan, R.P. Polybacterial Human Disease: The Ills of Social Networking. Trends Microbiol. 2014, 22, 508–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schink, B. Synergistic Interactions in the Microbial World. Antonie Van Leeuwenhoek 2002, 81, 257–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elias, S.; Banin, E. Multi-Species Biofilms: Living with Friendly Neighbors. FEMS Microbiol. Rev. 2012, 36, 990–1004. [Google Scholar] [CrossRef]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial Resistance and Virulence: A Successful or Deleterious Association in the Bacterial World? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef] [Green Version]
- Woolard, M.D.; Frelinger, J.A. Outsmarting the Host: Bacteria Modulating the Immune Response. Immunol. Res. 2008, 41, 188–202. [Google Scholar] [CrossRef]
- Phelan, V.V.; Liu, W.-T.; Pogliano, K.; Dorrestein, P.C. Microbial Metabolic Exchange--the Chemotype-to-Phenotype Link. Nat. Chem. Biol. 2011, 8, 26–35. [Google Scholar] [CrossRef]
- Van Delden, C.; Iglewski, B.H. Cell-to-Cell Signaling and Pseudomonas Aeruginosa Infections. Emerg. Infect Dis. 1998, 4, 551–560. [Google Scholar] [CrossRef]
- Rettger, L.F. The Antagonism of Bacteria and Their Products to Other Bacteria. J. Infect. Dis. 1905, 2, 562–568. [Google Scholar] [CrossRef]
- Byrd, A.L.; Segre, J.A. Infectious Disease. Adapting Koch’s Postulates. Science 2016, 351, 224–226. [Google Scholar] [CrossRef]
- Lee, K.; Lee, K.-M.; Kim, D.; Yoon, S.S. Molecular Determinants of the Thickened Matrix in a Dual-Species Pseudomonas Aeruginosa and Enterococcus Faecalis Biofilm. Appl. Environ. Microbiol. 2017, 83, e01182-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coenye, T.; Nelis, H.J. In Vitro and in Vivo Model Systems to Study Microbial Biofilm Formation. J. Microbiol. Methods 2010, 83, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Pereira, U.A.; Barbosa, L.C.A.; Maltha, C.R.A.; Demuner, A.J.; Masood, M.A.; Pimenta, A.L. Inhibition of Enterococcus Faecalis Biofilm Formation by Highly Active Lactones and Lactams Analogues of Rubrolides. Eur. J. Med. Chem. 2014, 82, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.M.; Luo, L.; Ardita, C.S.; Richardson, A.N.; Kwon, Y.M.; Mercante, J.W.; Alam, A.; Gates, C.L.; Wu, H.; Swanson, P.A.; et al. Symbiotic Lactobacilli Stimulate Gut Epithelial Proliferation via Nox-Mediated Generation of Reactive Oxygen Species. EMBO J. 2013, 32, 3017–3028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rada, B.; Leto, T.L. Pyocyanin Effects on Respiratory Epithelium: Relevance in Pseudomonas Aeruginosa Airway Infections. Trends Microbiol. 2013, 21, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Jayaseelan, S.; Ramaswamy, D.; Dharmaraj, S. Pyocyanin: Production, Applications, Challenges and New Insights. World J. Microbiol. Biotechnol. 2014, 30, 1159–1168. [Google Scholar] [CrossRef]
- Zhu, K.; Chen, S.; Sysoeva, T.A.; You, L. Universal Antibiotic Tolerance Arising from Antibiotic-Triggered Accumulation of Pyocyanin in Pseudomonas Aeruginosa. PLoS Biol. 2019, 17, e3000573. [Google Scholar] [CrossRef] [PubMed]
- Rada, B.; Lekstrom, K.; Damian, S.; Dupuy, C.; Leto, T.L. The Pseudomonas Toxin Pyocyanin Inhibits the Dual Oxidase-Based Antimicrobial System as It Imposes Oxidative Stress on Airway Epithelial Cells. J. Immunol. 2008, 181, 4883–4893. [Google Scholar] [CrossRef] [Green Version]
- Jones-Nelson, O.; Hilliard, J.J.; DiGiandomenico, A.; Warrener, P.; Alfaro, A.; Cheng, L.; Stover, C.K.; Cohen, T.S.; Sellman, B.R. The Neutrophilic Response to Pseudomonas Damages the Airway Barrier, Promoting Infection by Klebsiella Pneumoniae. Am. J. Respir. Cell Mol. Biol. 2018, 59, 745–756. [Google Scholar] [CrossRef]
- Panayidou, S.; Georgiades, K.; Christofi, T.; Tamana, S.; Promponas, V.J.; Apidianakis, Y. Pseudomonas Aeruginosa Core Metabolism Exerts a Widespread Growth-Independent Control on Virulence. Sci. Rep. 2020, 10, 9505. [Google Scholar] [CrossRef]
- Chandler, J.A.; Morgan Lang, J.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial Communities of Diverse Drosophila Species: Ecological Context of a Host–Microbe Model System. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.; Fan, L.; Yan, J.; Jiang, Y.; Doucette, C.; Fillmore, S.; Walker, B. Influence of Culture Media, PH and Temperature on Growth and Bacteriocin Production of Bacteriocinogenic Lactic Acid Bacteria. AMB Express 2018, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, C.L.; Hogue, L.A.; Vajravelu, R.K.; Capps, G.H.; Ibricevic, A.; Hindi, K.M.; Kascatan-Nebioglu, A.; Walter, M.J.; Brody, S.L.; Youngs, W.J. In Vitro and Murine Efficacy and Toxicity Studies of Nebulized SCC1, a Methylated Caffeine-Silver(I) Complex, for Treatment of Pulmonary Infections. Antimicrob. Agents Chemother. 2009, 53, 3285–3293. [Google Scholar] [CrossRef] [Green Version]
- Munder, A.; Wölbeling, F.; Kerber-Momot, T.; Wedekind, D.; Baumann, U.; Gulbins, E.; Tümmler, B. Acute Intratracheal Pseudomonas Aeruginosa Infection in Cystic Fibrosis Mice Is Age-Independent. Respir. Res. 2011, 12, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teichgräber, V.; Ulrich, M.; Endlich, N.; Riethmüller, J.; Wilker, B.; de Oliveira-Munding, C.C.; van Heeckeren, A.M.; Barr, M.L.; von Kürthy, G.; Schmid, K.W.; et al. Ceramide Accumulation Mediates Inflammation, Cell Death and Infection Susceptibility in Cystic Fibrosis. Nat. Med. 2008, 14, 382–391. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charalambous, A.; Grivogiannis, E.; Dieronitou, I.; Michael, C.; Rahme, L.; Apidianakis, Y. Proteobacteria and Firmicutes Secreted Factors Exert Distinct Effects on Pseudomonas aeruginosa Infection under Normoxia or Mild Hypoxia. Metabolites 2022, 12, 449. https://doi.org/10.3390/metabo12050449
Charalambous A, Grivogiannis E, Dieronitou I, Michael C, Rahme L, Apidianakis Y. Proteobacteria and Firmicutes Secreted Factors Exert Distinct Effects on Pseudomonas aeruginosa Infection under Normoxia or Mild Hypoxia. Metabolites. 2022; 12(5):449. https://doi.org/10.3390/metabo12050449
Chicago/Turabian StyleCharalambous, Anna, Evangelos Grivogiannis, Irene Dieronitou, Christina Michael, Laurence Rahme, and Yiorgos Apidianakis. 2022. "Proteobacteria and Firmicutes Secreted Factors Exert Distinct Effects on Pseudomonas aeruginosa Infection under Normoxia or Mild Hypoxia" Metabolites 12, no. 5: 449. https://doi.org/10.3390/metabo12050449