
A Metabolomic Severity Score for Airflow Obstruction and Emphysema

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Demographic Characteristics
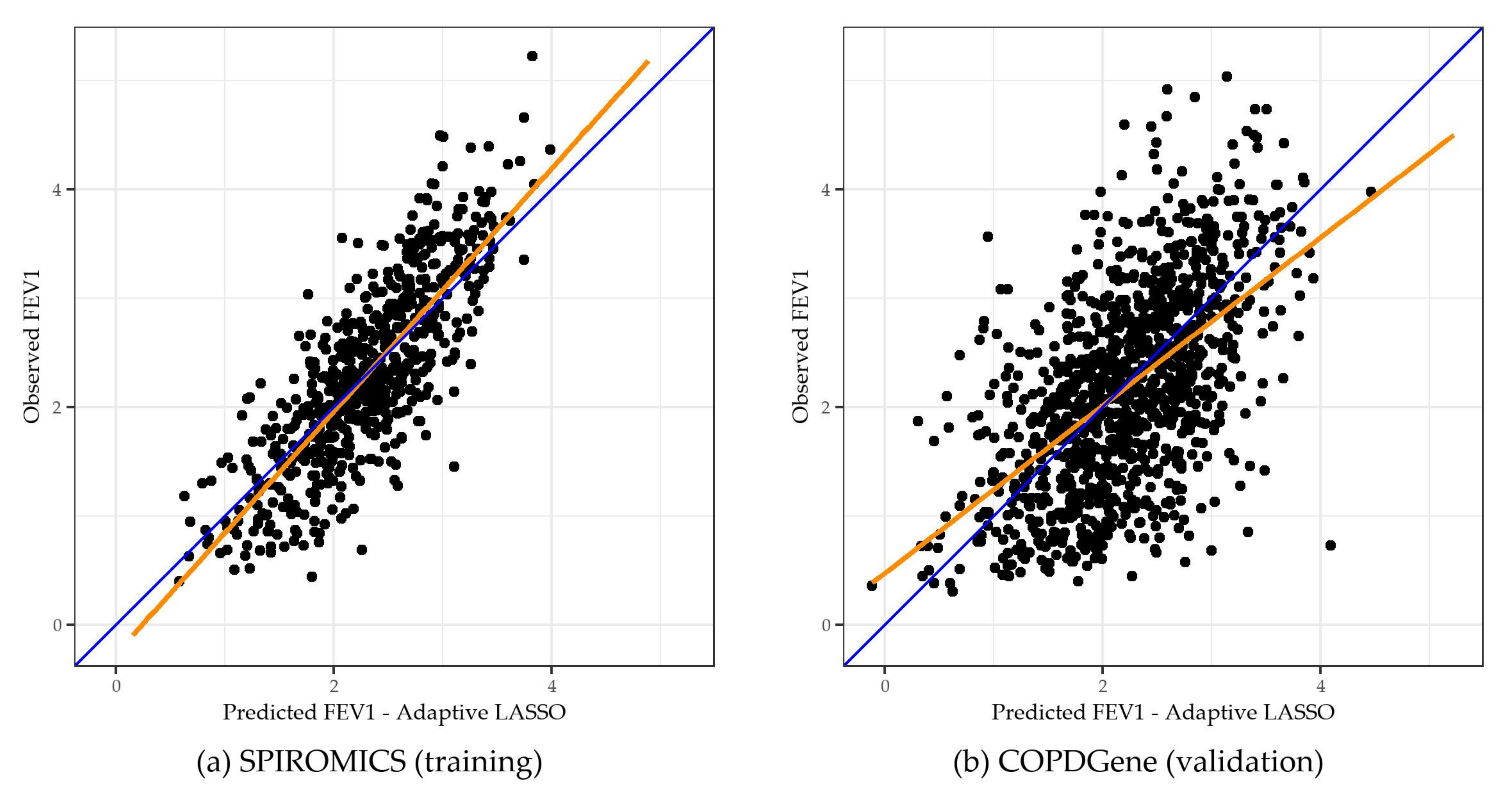
2.2. Adaptive LASSO Results
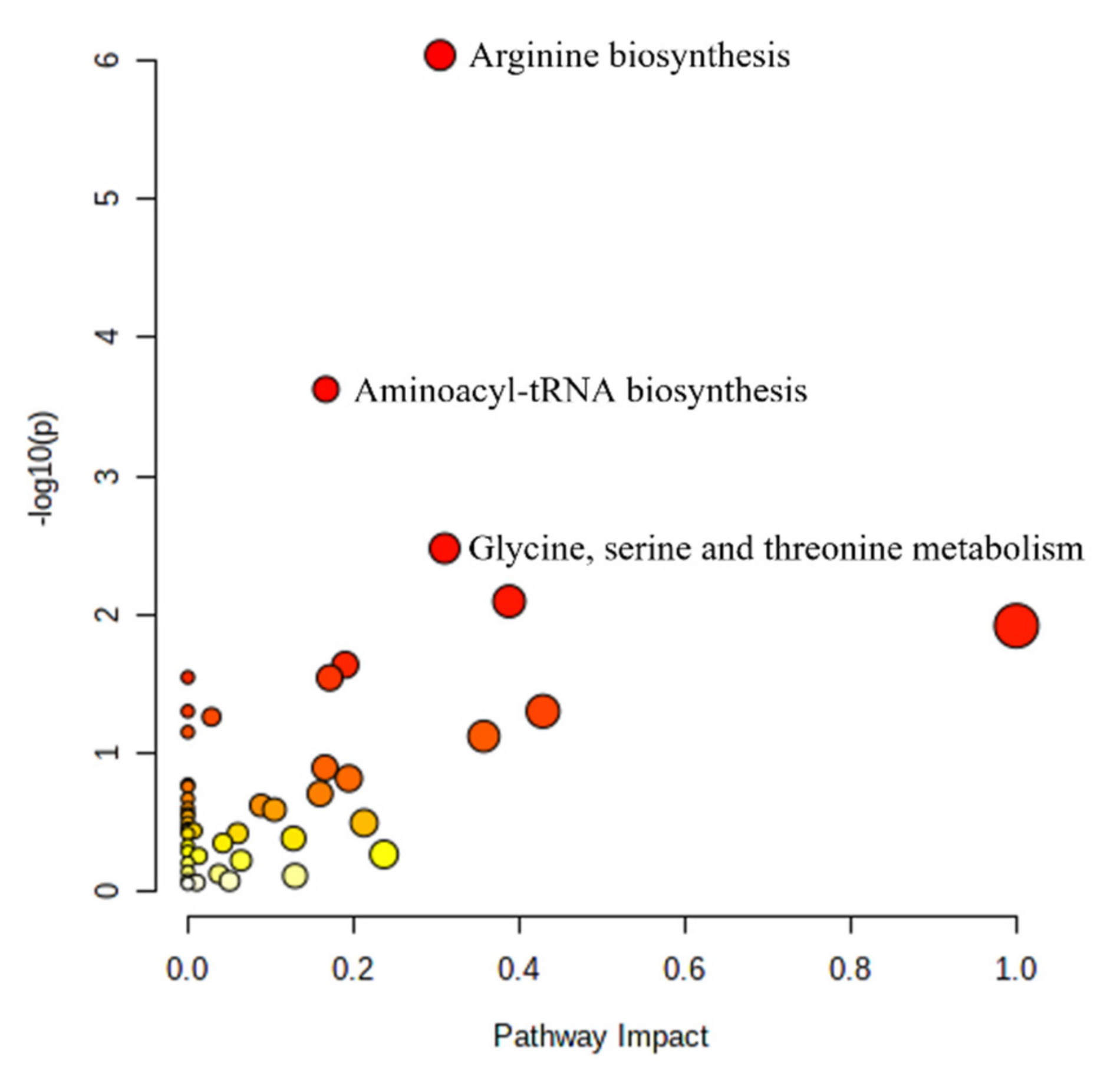
2.3. Pathway Analysis
2.4. Sensitivity Analysis
3. Discussion
4. Materials and Methods
4.1. Study Populations
4.1.1. Training Cohort
4.1.2. Validation Cohort
4.2. Data and Definitions
4.2.1. Clinical Data and Definitions
4.2.2. Metabolomic Profiling and Processing
4.3. Analysis
4.3.1. Statistical and Bioinformatics Analysis
4.3.2. Pathway Analysis
4.3.3. Software
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Initiative for Chronic Obstructive Lung Disease—GOLD. Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Pulmonary Disease (2022 Report); Global Initiative for Chronic Obstructive Lung Disease—GOLD: Fontana, WI, USA, 2021. [Google Scholar]
- Mannino, D.M.; Gagnon, R.C.; Petty, T.L.; Lydick, E. Obstructive lung disease and low lung function in adults in the United States: Data from the National Health and Nutrition Examination Survey, 1988–1994. Arch. Intern. Med. 2000, 160, 1683–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Chronic Obstructive Pulmonary Disease (COPD). 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/chronic-obstructive-pulmonary-disease-(copd) (accessed on 21 June 2021).
- Cruickshank-Quinn, C.; Jacobson, S.; Hughes, G.; Powell, R.L.; Petrache, I.; Kechris, K.; Bowler, R.; Reisdorph, N. Metabolomics and transcriptomics pathway approach reveals outcome-specific perturbations in COPD. Sci. Rep. 2018, 8, 17132. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Flexeder, C.; McGarrah, R.W.; Wyss, A.; Morrison, A.C.; North, K.E.; Boerwinkle, E.; Kastenmüller, G.; Gieger, C.; Suhre, K.; et al. Metabolomics Identifies Novel Blood Biomarkers of Pulmonary Function and COPD in the General Population. Metabolities 2019, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, R.S.; Stewart, I.D.; Bayne, H.; Kachroo, P.; Spiro, A., III; Vokonas, P.; Sparrow, D.; Weiss, S.T.; Knihtilä, H.M.; Litonjua, A.A.; et al. Metabolomic differences in lung function metrics: Evidence from two cohorts. Thorax 2021. [Google Scholar]
- Vaarhorst, A.A.; Verhoeven, A.; Weller, C.M.; Böhringer, S.; Göraler, S.; Meissner, A.; Deelder, A.M.; Henneman, P.; Gorgels, A.P.; van den Brandt, P.; et al. A metabolomic profile is associated with the risk of incident coronary heart disease. Am. Heart J. 2014, 168, 45–52.e7. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, C.; Nambi, V.; Morrison, A.C.; Folsom, A.R.; Ballantyne, C.M.; Boerwinkle, E.; Yu, B. Metabolomic Pattern Predicts Incident Coronary Heart Disease: Findings from the Atherosclerosis Risk in Communities Study. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 7. [Google Scholar] [CrossRef]
- Geidenstam, N.; Hsu, Y.-H.H.; Astley, C.M.; Mercader, J.M.; Ridderstråle, M.; Gonzalez, M.E.; Gonzalez, C.; Hirschorn, J.N.; Salem, R.M. Using metabolite profiling to construct and validate a metabolite risk score for predicting future weight gain. PLoS ONE 2019, 14, e0222445. [Google Scholar] [CrossRef]
- Floegel, A.; Stefan, N.; Yu, Z.; Mühlenbruch, K.; Drogan, D.; Joost, H.-G.; Fritsche, A.; Häring, H.-U.; de Angelis, M.H.; Peters, A.; et al. Identification of Serum Metabolites Associated with Risk of Type 2 Diabetes Using a Targeted Metabolomic Approach. Diabetes 2013, 62, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Pinto-Plata, V.; Casanova, C.; Divo, M.; Tesfaigzi, Y.; Calhoun, V.; Sui, J.; Polverino, F.; Priolo, C.; Petersen, H.; de Torres, J.P.; et al. Plasma metabolomics and clinical predictors of survival differences in COPD patients. Respir. Res. 2019, 20, 219. [Google Scholar] [CrossRef] [Green Version]
- Moll, M.; Sakornsakolpat, P.; Shrine, N.; Hobbs, B.; DeMeo, D.L.; John, C.; Guyatt, A.L.; McGeachie, M.J.; Gharib, S.A.; Obeidat, M.; et al. Chronic obstructive pulmonary disease and related phenotypes: Polygenic risk scores in population-based and case-control cohorts. Lancet Respir. Med. 2020, 8, 696–708. [Google Scholar] [CrossRef]
- Huang, J.; Ma, S.; Zhang, C.-H. Adaptive LASSO for Sparse High-Dimensional Regression Models. Stat. Sin. 2008, 18, 1603–1618. [Google Scholar]
- Zou, H. The Adaptive Lasso and Its Oracle Properties. J. Am. Stat. Assoc. 2006, 101, 1418–1429. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.A.; Duongh, M.; Young, A.W.; Subbarao, P.; Gauvreau, G.M.; Grasemann, H. Asymmetric Dimethylarginine in Chronic Obstructive Pulmonary Disease (ADMA in COPD). Int. J. Mol. Sci. 2014, 15, 6062–6071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, F.S.; Jiang, X.-C.; Schwartz, J.E.; Hoffman, E.A.; Yeboah, J.; Shea, S.; Burkart, C.M.; Barr, R.G. Plasma sphingomyelin and longitudinal change in percent emphysema on CT. The MESA Lung study. Biomarkers 2014, 19, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Wheelock, C.; Balgoma, D.; Grunewald, J.; Eklund, A.; Skold, M.; Wheelock, A. Lipid mediator levels evidence gender-specific increases in bronchoalveolar lavage fluid of COPD patients relative to healthy smokers. In Proceedings of the European Respiratory Society Annual Congress, Barcelona, Spain, 7–11 September 2013. [Google Scholar]
- Ubhi, B.K.; Cheng, K.K.; Dong, J.; Janowitz, T.; Jodrell, D.; Tal-Singer, R.; MacNee, W.; Lomas, D.A.; Riley, J.H.; Griffin, J.L.; et al. Targeted metabolomics identifies perturbations in amino acid metabolism that sub-classify patients with COPD. Mol. BioSyst. 2012, 8, 3125–3133. [Google Scholar] [CrossRef]
- Ubhi, B.K.; Riley, J.H.; Shaw, P.A.; Lomas, D.A.; Tal-Singer, R.; MacNee, W.; Griffin, J.L.; Connor, S.C. Metabolic profiling detects biomarkers of protein degradation in COPD patients. Eur. Respir. J. 2012, 40, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Aydin, M.; Altintas, N.; Mutlu, L.C.; Bilir, B.; Oran, M.; Tulubas, F.; Topçu, B.; Tayfur, I.; Kucukyalcin, V.; Kaplan, G.; et al. Asymmetric dimethylarginine contributes to airway nitric oxide deficiency in patients with COPD. Clin. Respir. J. 2017, 11, 318–327. [Google Scholar] [CrossRef]
- Ruzsics, I.; Nagy, L.; Keki, S.; Sarosi, V.; Illes, B.; Illes, Z.; Horvath, I.; Bogar, L.; Molnar, T. L-Arginine Pathway in COPD Patients with Acute Exacerbation: A New Potential Biomarker. COPD J. Chronic Obstr. Pulm. Dis. 2016, 13, 139–145. [Google Scholar] [CrossRef]
- Jonker, R.; Deutz, N.E.; Erbland, M.L.; Anderson, P.J.; Engelen, M.P. Alterations in whole-body arginine metabolism in chronic obstructive pulmonary disease. Am. J. Clin. Nutr. 2016, 103, 1458–1464. [Google Scholar] [CrossRef] [Green Version]
- Santos Valença, S.; Rueff-Barroso, C.R.; Alves Pimenta, W.; Correa Melo, A.; Tiscoski Nesi, R.; Santos Silva, M.A.; Porto, L.C. L-NAME and L-arginine differentially ameliorate cigarette smoke-induced emphysema in mice. Pulm. Pharmacol. Ther. 2011, 24, 587–594. [Google Scholar] [CrossRef]
- Kilk, K.; Aug, A.; Ottas, A.; Soomets, U.; Altraja, S.; Altraja, A. Phenotyping of Chronic Obstructive Pulmonary Disease Based on the Integration of Metabolomes and Clinical Characteristics. Int. J. Mol. Sci. 2018, 19, 666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naz, S.; Kolmert, J.; Yang, M.; Reinke, S.N.; Kamleh, M.A.; Snowden, S.; Heyder, T.; Levänen, B.; Erle, D.J.; Sköld, C.M.; et al. Metabolomics analysis identifies sex-associated metabotypes of oxidative stress and the autotaxin–lysoPA axis in COPD. Eur. Respir. J. 2017, 49, 1602322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Chen, Z.; Fang, Z.; Zhao, J.; Zhou, Y.; Tang, C. Multi-omics study on biomarker and pathway discovery of chronic ob-structive pulmonary disease. J. Breath Res. 2021, 15, 044001. [Google Scholar] [CrossRef] [PubMed]
- Ran, N.; Pang, Z.; Gu, Y.; Pan, H.; Zuo, X.; Guan, X.; Yuan, Y.; Wang, Z.; Guo, Y.; Cui, Z.; et al. An updated overview of metabolomic profile changes in chronic obstructive pulmonary disease. Metabolites 2019, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Engelen, M.P.K.J.; Wouters, E.F.M.; Deutz, N.E.P.; Does, J.D.; Schols, A.M.W.J. Effects of Exercise on Amino Acid Metabolism in Patients with Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2001, 163, 859–864. [Google Scholar] [CrossRef]
- Gillenwater, L.; Kechris, K.; Pratte, K.; DeMeo, D.; Bowler, R. Metabolomic Profiling Reveals Sex Specific Associations with COPD and Emphysema. Metabolites 2020, 11, 17. [Google Scholar]
- Couper, D.; LaVange, L.M.; Han, M.; Barr, R.G.; Bleecker, E.; Hoffman, E.; Kanner, R.; Kleerup, E.; Martinez, F.J.; Woodruff, P.G.; et al. Design of the Subpopulations and Intermediate Outcomes in COPD Study (SPIROMICS). Thorax 2013, 69, 492–495. [Google Scholar] [CrossRef] [Green Version]
- Regan, E.A.; Hokanson, J.E.; Murphy, J.R.; Make, B.; Lynch, D.A.; Beaty, T.H.; Curran-Everett, D.; Silverman, E.K.; Crapo, J.D. Genetic Epidemiology of COPD (COPDGene) Study Design. COPD J. Chronic Obstr. Pulm. Dis. 2010, 7, 32–43. [Google Scholar] [CrossRef]
- Gillenwater, L.A.; Pratte, K.A.; Hobbs, B.D.; Cho, M.H.; Zhuang, Y.; Halper-Stromberg, E.; Cruickshank-Quinn, C.; Reisdorph, N.; Petrache, I.; Labaki, W.W.; et al. Plasma Metabolomic Signatures of Chronic Obstructive Pulmonary Disease and the Impact of Genetic Variants on Phenotype-Driven Modules. Netw. Syst. Med. 2020, 3, 159–181. [Google Scholar] [CrossRef]
- Hoffman, E.; Ahmed, F.S.; Baumhauer, H.; Budoff, M.; Carr, J.; Kronmal, R.; Reddy, S.; Barr, R.G. Variation in the Percent of Emphysema-like Lung in a Healthy, Nonsmoking Multiethnic Sample. The MESA Lung Study. Ann. Am. Thorac. Soc. 2014, 11, 898–907. [Google Scholar] [CrossRef] [Green Version]
- DeHaven, C.D.; Evans, A.M.; Dai, H.; Lawton, K.A. Organization of GC/MS and LC/MS metabolomics data into chemical libraries. J. Cheminform. 2010, 2, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, A.M.; DeHaven, C.D.; Barrett, T.; Mitchell, M.; Milgram, E. Integrated, nontargeted ultrahigh performance liquid chromatography/electrospray ionization tandem mass spectrometry platform for the identification and relative quantification of the small-molecule complement of biological systems. Anal. Chem. 2009, 81, 6656–6667. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.J.; Kennedy, A.D.; Eckhart, A.D.; Burrage, L.C.; Wulff, J.E.; Miller, L.A.; Milburn, M.V.; Ryals, J.A.; Beaudet, A.L.; Sun, Q.; et al. Untargeted metabolomic analysis for the clinical screening of inborn errors of metabolism. J. Inherit. Metab. Dis. 2015, 38, 1029–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Wishart, D.S. MetPA: A web-based metabolomics tool for pathway analysis and visualization. Bioinformatics 2010, 26, 2342–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Characteristic | SPIROMICS (Training) (n = 648) | COPDGene (Validation) (n = 1120) | p |
|---|---|---|---|
| Age (yrs): mean (sd) | 63.2 (8.89) | 67.3 (8.82) | <0.001 |
| Sex: n (%) | |||
| Male | 349 (53.9) | 562 (50.2) | 0.149 |
| Female | 299 (46.1) | 558 (49.8) | |
| Race/Ethnicity: n (%) | |||
| Non-Hispanic, White | 469 (72.4) | 1025 (91.5) | <0.001 |
| Black/African American | 122 (18.8) | 95 (8.5) | |
| Other | 57 (8.8) | 0 (0) | |
| BMI (kg/m2) | 28.4 (5.22) | 28.8 (6.14) | 0.206 |
| Spirometry category †: n (%) | |||
| PRISm | 11 (1.7) | 101 (9.1) | <0.001 |
| GOLD 0 | 261 (40.3) | 505 (45.7) | |
| GOLD 1 | 113 (17.5) | 114 (10.3) | |
| GOLD 2 | 181 (28) | 208 (18.8) | |
| GOLD 3 | 66 (10.2) | 115 (10.4) | |
| GOLD 4 | 15 (2.3) | 62 (5.6) | |
| Smoking Status: n (%) | |||
| Never Smoker | 57 (8.9) | 65 (5.8) | <0.001 |
| Former Smoker | 369 (57.6) | 787 (70.3) | |
| Current Smoker | 215 (33.5) | 268 (23.9) | |
| Smoking pack-yrs: mean (sd) | 45.1 (30.94) | 42.4 (26.18) | 0.048 |
| Postbronchodilator FEV1 (L): mean (sd) | 2.32 (0.84) | 2.19 (0.91) | 0.003 |
| Postbronchodilator FEV1/FVC: mean (sd) | 0.65 (0.15) | 0.67 (0.15) | 0.008 |
| Postbronchodilator FEV1 percent predicted: mean (sd) | 80.7 (23.5) | 79.3 (26.5) | 0.297 |
| Percent Emphysema ‡: mean (sd) | 5.79 (8.08) | 6.66 (9.8) | 0.058 |
| MESA-adjusted lung density (g/L): mean (sd) | 86.1 (24.4) | 81.7 (22.6) | <0.001 |
| Metabolite | adaLASSO β | Super Pathway (Metabolon) | Sub Pathway (Metabolon) |
|---|---|---|---|
| vanillylmandelate (VMA) | −0.5654 | Amino Acid | Tyrosine Metabolism |
| N1-methyladenosine | −0.3571 | Nucleotide | Purine Metabolism, Adenine containing |
| Glutamine | −0.3414 | Amino Acid | Glutamate Metabolism |
| 2-hydroxypalmitate | −0.3264 | Lipid | Fatty Acid, Monohydroxy |
| choline phosphate | 0.2924 | Lipid | Phospholipid Metabolism |
| 1-palmitoyl-2-stearoyl-GPC (16:0/18:0) | 0.2912 | Lipid | Phosphatidylcholine (PC) |
| cerotoylcarnitine (C26) * | 0.2859 | Lipid | Fatty Acid Metabolism (Acyl Carnitine, Long Chain Saturated) |
| phenylalanine | −0.2675 | Amino Acid | Phenylalanine Metabolism |
| dimethylarginine (SDMA + ADMA) | 0.2646 | Amino Acid | Urea cycle; Arginine and Proline Metabolism |
| myo-inositol | 0.2568 | Lipid | Inositol Metabolism |
| imidazole lactate | 0.2561 | Amino Acid | Histidine Metabolism |
| 1-stearoyl-2-arachidonoyl-GPC (18:0/20:4) | −0.2456 | Lipid | Phosphatidylcholine (PC) |
| N-acetylvaline | 0.2374 | Amino Acid | Leucine, Isoleucine and Valine Metabolism |
| taurine | −0.2325 | Amino Acid | Methionine, Cysteine, SAM, and Taurine Metabolism |
| sulfate * | 0.2325 | Xenobiotics | Chemical |
| 3-methyl-2-oxovalerate | −0.2206 | Amino Acid | Leucine, Isoleucine, and Valine Metabolism |
| gamma-glutamylthreonine | 0.2162 | Peptide | Gamma-glutamyl Amino Acid |
| proline | 0.2127 | Amino Acid | Urea cycle; Arginine and Proline Metabolism |
| mannonate * | −0.2107 | Xenobiotics | Food Component/Plant |
| retinol (vitamin A) | 0.2103 | Cofactors and Vitamins | Vitamin A Metabolism |
| sphingomyelin (d18:2/21:0, d16:2/23:0) * | −0.1919 | Lipid | Sphingomyelins |
| N-acetylcarnosine | 0.1825 | Amino Acid | Histidine Metabolism |
| 3beta-hydroxy-5-cholestenoate | 0.1798 | Lipid | Sterol |
| pimelate (C7-DC) | −0.1790 | Lipid | Fatty Acid, Dicarboxylate |
| FEV1 (132 Metabolites) | ||||||
|---|---|---|---|---|---|---|
| Adjusted R2 | MSE | |||||
| Clinical Covariates 1 Only | metSS Only | metSS + Covariates 1 | Clinical Covariates 1 Only | metSS Only | metSS + Covariates 1 | |
| SPIROMICS (training) | 42.1 | 64.8 | 68.9 | 0.397 | 0.246 | 0.213 |
| COPDGene (validation) | 46.4 | 31.7 | 53.4 | 0.435 | 0.559 | 0.378 |
| KEGG Pathway | adaLASSO (β) |
|---|---|
| Arginine Biosynthesis | |
| glutamine | −0.34 |
| arginine | −0.11 |
| N-acetylglutamate | −0.10 |
| citrulline | −0.10 |
| alpha-ketoglutarate | −0.07 |
| aspartate | 0.07 |
| fumarate | −0.06 |
| Aminoacyl-tRNA Biosynthesis | |
| glutamine | −0.34 |
| phenylalanine | −0.27 |
| proline | 0.21 |
| threonine | −0.16 |
| tyrosine | −0.12 |
| arginine | −0.11 |
| aspartate | 0.07 |
| serine | 0.05 |
| cysteine | 0.05 |
| Glycine, serine, and threonine metabolism | |
| threonine | −0.16 |
| creatine | −0.12 |
| serine | 0.05 |
| cysteine | 0.05 |
| sarcosine | −0.03 |
| choline | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godbole, S.; Labaki, W.W.; Pratte, K.A.; Hill, A.; Moll, M.; Hastie, A.T.; Peters, S.P.; Gregory, A.; Ortega, V.E.; DeMeo, D.; et al. A Metabolomic Severity Score for Airflow Obstruction and Emphysema. Metabolites 2022, 12, 368. https://doi.org/10.3390/metabo12050368
Godbole S, Labaki WW, Pratte KA, Hill A, Moll M, Hastie AT, Peters SP, Gregory A, Ortega VE, DeMeo D, et al. A Metabolomic Severity Score for Airflow Obstruction and Emphysema. Metabolites. 2022; 12(5):368. https://doi.org/10.3390/metabo12050368
Chicago/Turabian StyleGodbole, Suneeta, Wassim W. Labaki, Katherine A. Pratte, Andrew Hill, Matthew Moll, Annette T. Hastie, Stephen P. Peters, Andrew Gregory, Victor E. Ortega, Dawn DeMeo, and et al. 2022. "A Metabolomic Severity Score for Airflow Obstruction and Emphysema" Metabolites 12, no. 5: 368. https://doi.org/10.3390/metabo12050368