


Chemical and Biological Properties of Three Poorly Studied Species of Lycium Genus—Short Review

Abstract
:1. Introduction
2. Other Lycium Species
2.1. Lycium europaeum L.
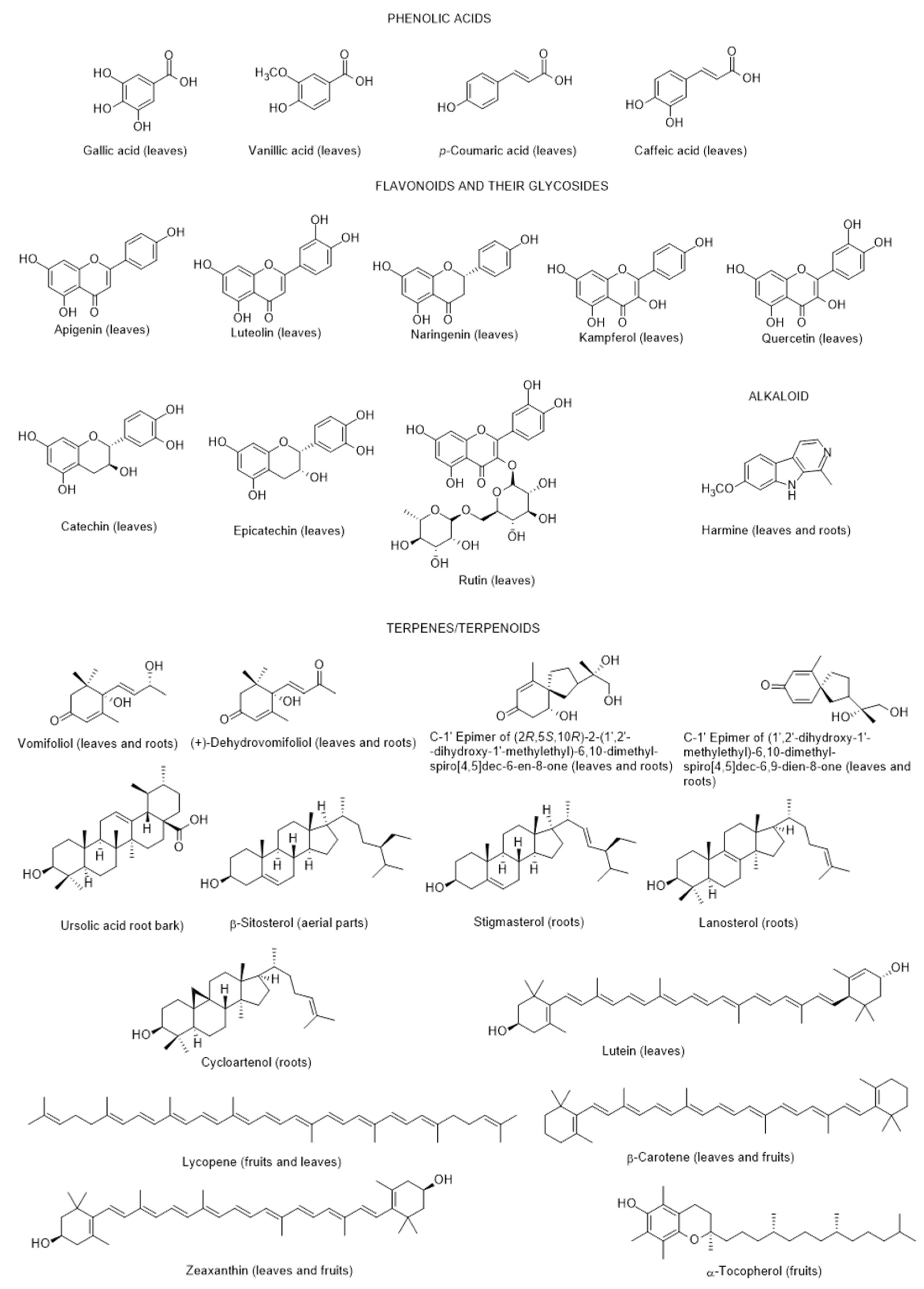
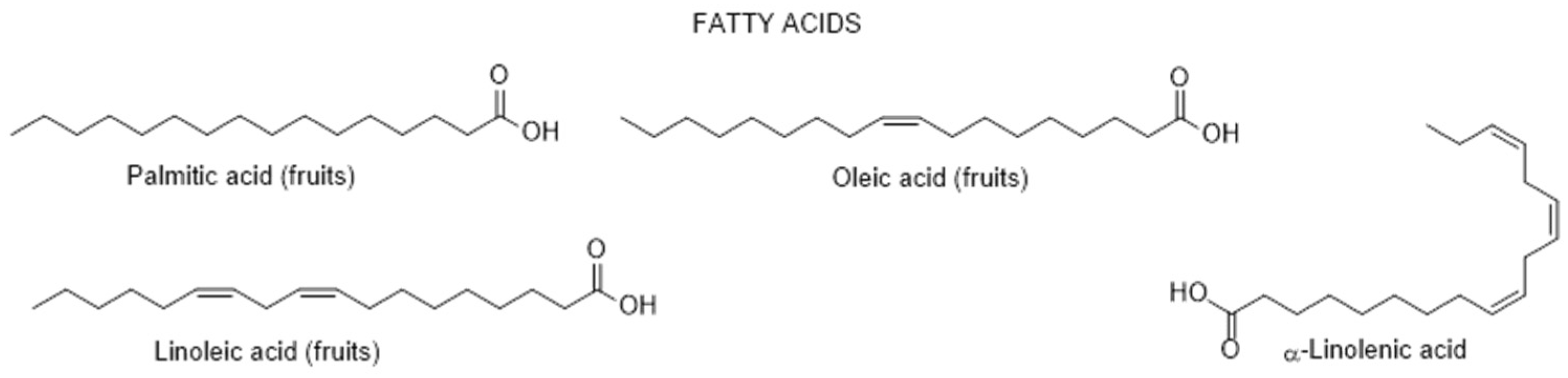
2.1.1. Chemical Composition and Biological Properties
2.1.2. Fungal Infection
2.2. Lycium schweinfurthii Dammer
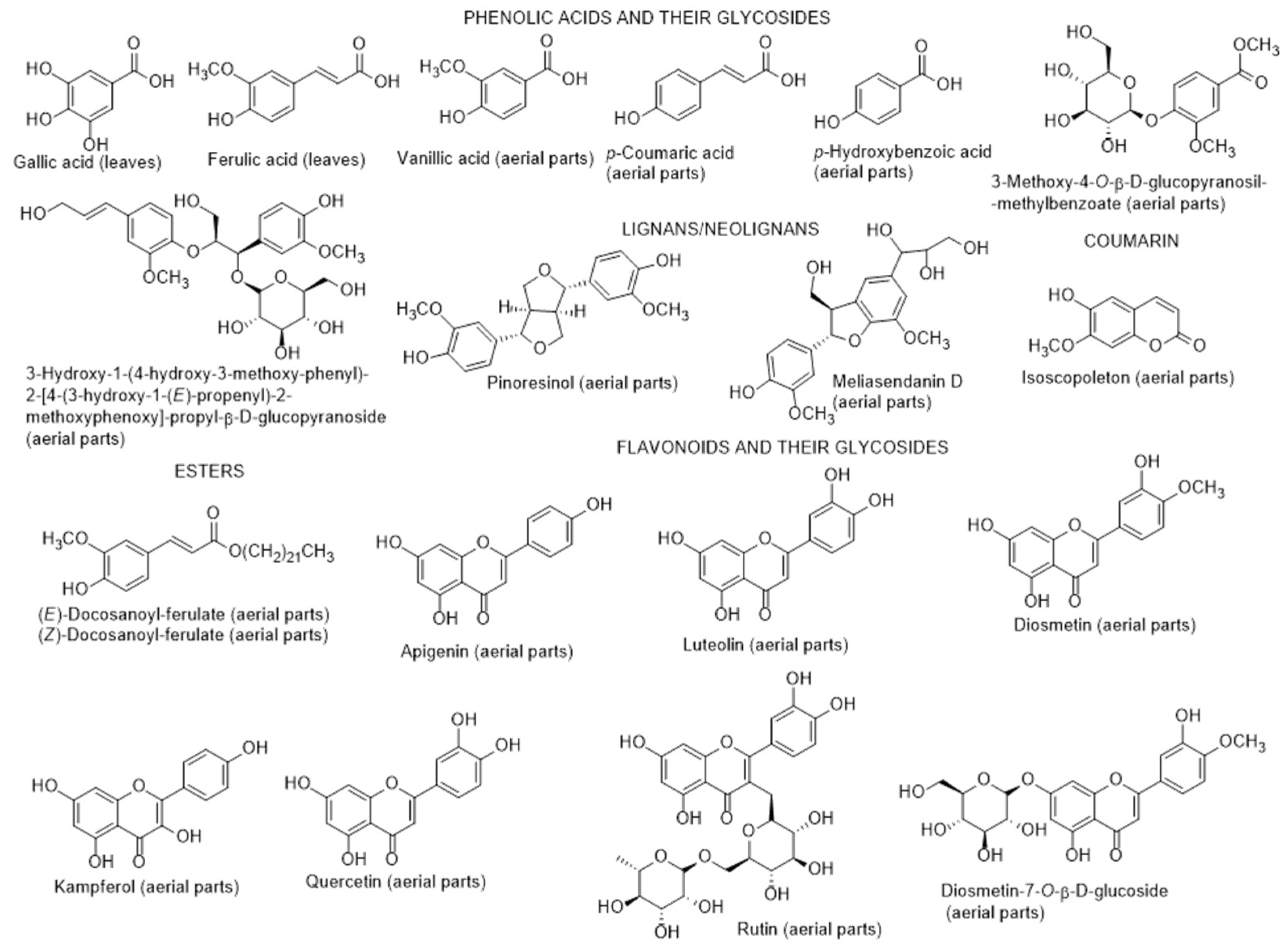
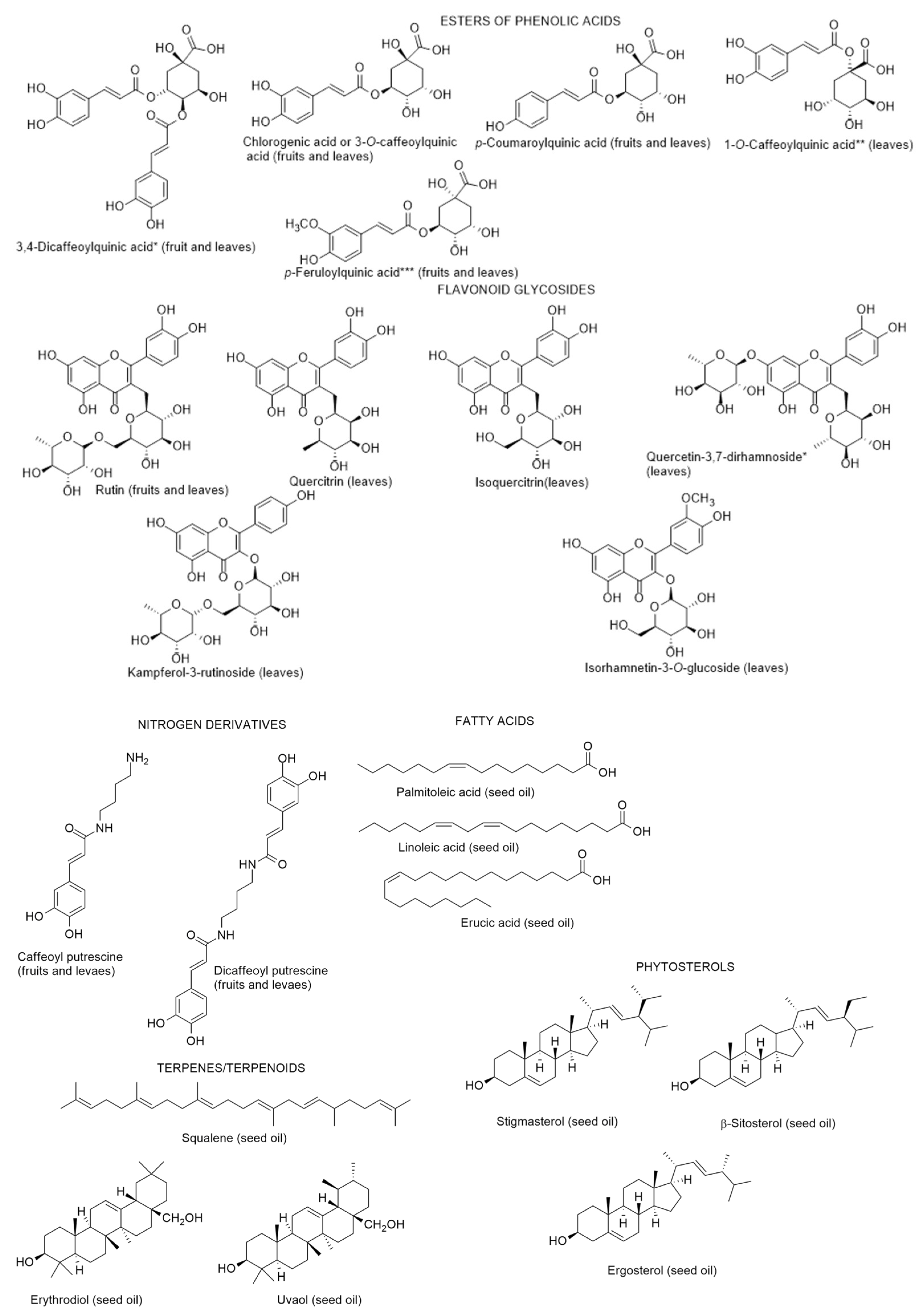
2.2.1. Chemical Composition and Biological Properties
2.2.2. Biotechnological Production of Secondary Metabolites
2.2.3. Secondary Metabolites from Endophytic Fungus
2.3. Lycium intricatum Boiss
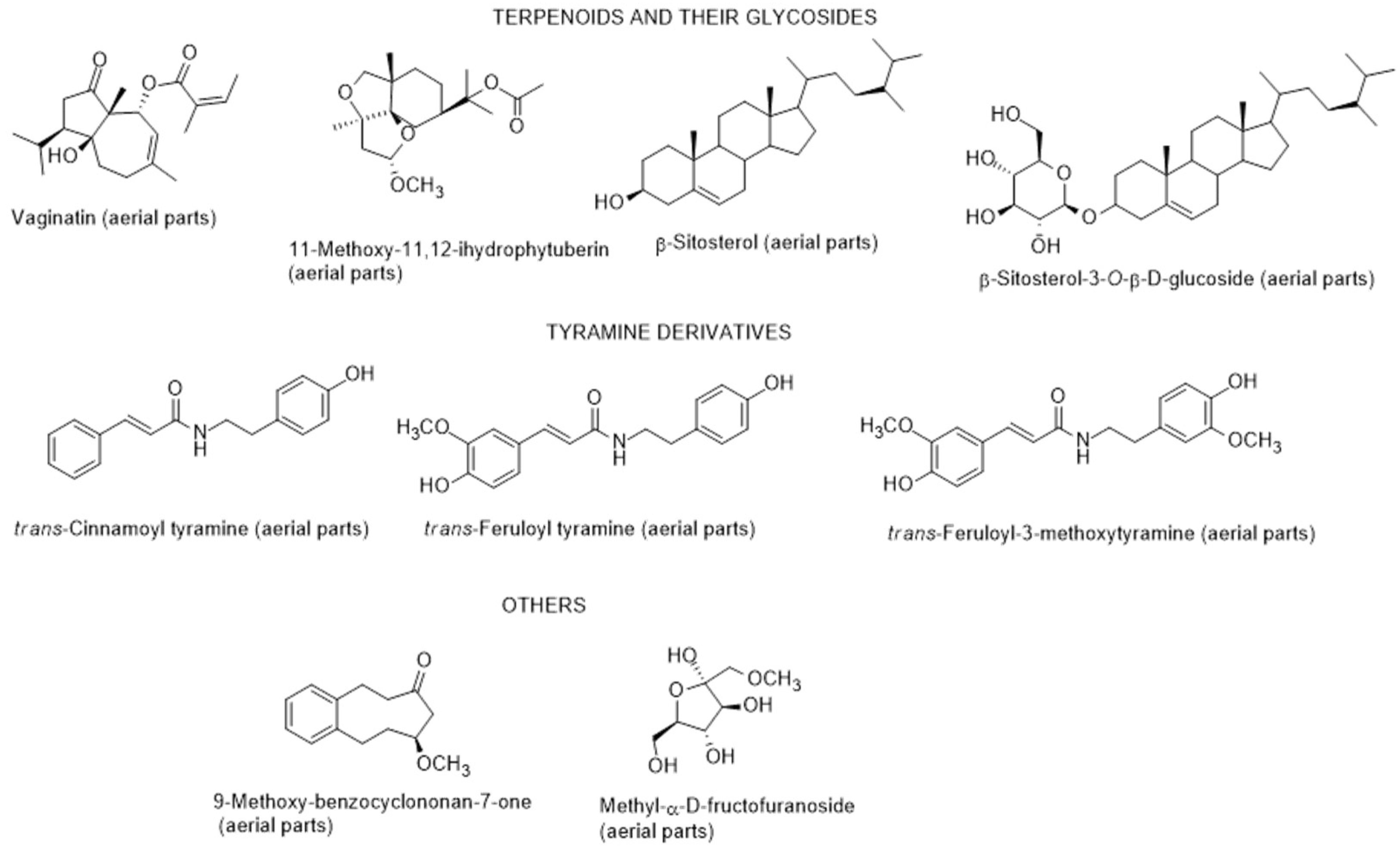
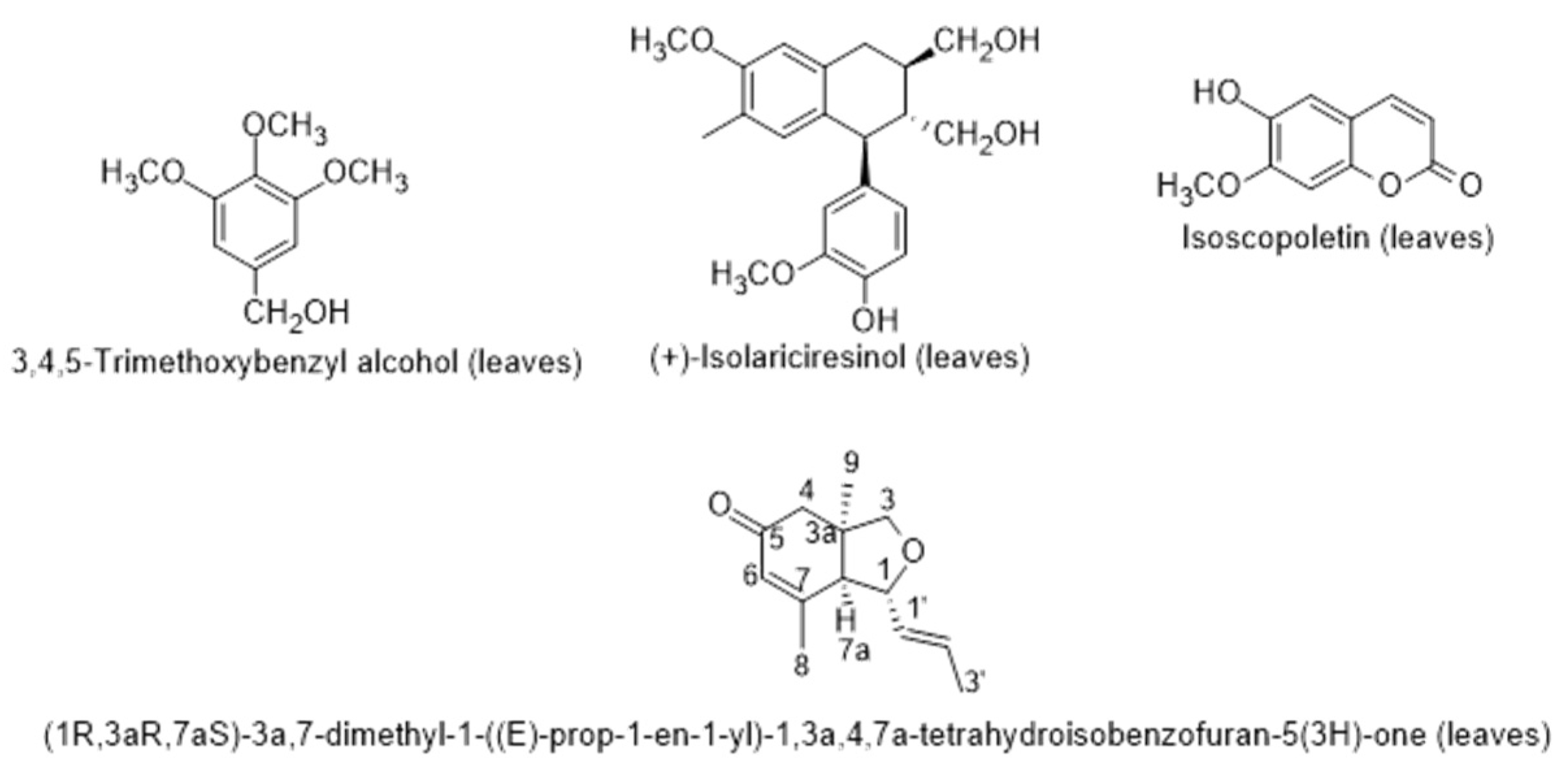
2.3.1. Chemical Composition and Biological Properties
2.3.2. Secondary Metabolites from Endophytic Fungus
3. Conclusions
4. Future Trend
Funding
Conflicts of Interest
References
- Joseph, S.V.; Edirisinghe, I.; Burton-Freeman, B.M. Berries: Anti-inflammatory effects in humans. J. Agric. Food Chem. 2014, 62, 3886–3903. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Heinrich, M.; Weckerle, C.S. The genus Lycium as food and medicine: A botanical, ethnobotanical and historical review. J. Ethnopharmacol. 2018, 212, 50–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Xia, T.; Dang, S.; Fan, G.; Wang, Z. Investigation of Chinese wolberry (Lycium spp.) germplasm by restriction site- associated DNA sequencing (RAD-seq). Biochem. Genet. 2018, 56, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, T.; Yokoyama, J.; Ohashi, H. Phylogeny and biogeography of the genus Lycium (Solanaceae): Inferences from chloroplast DNA sequences. Mol. Phylogenet. Evol. 2001, 19, 246–258. [Google Scholar] [CrossRef]
- Zhou, Z.; Xiao, J.; Fan, H.; Yu, Y.; He, R.; Feng, X.; Gao, H. Polyphenols from wolfberry and their bioactivities. Food Chem. 2017, 214, 644–654. [Google Scholar] [CrossRef]
- Xing, L.; Wang, Y.; Luo, R.; Li, X.; Zou, L. Determination of 31 pesticide residues in wolfberry by LC-MS/MS and dietary risk assessment of wolfberry consumption. Food Sci. Tecnol. 2022, 42, e61921. [Google Scholar] [CrossRef]
- Kulczyński, B.; Gramza-Michałowska, A. Gojy berry (Lycium barbarum): Composition and health effects—A review. Pol. J. Food Nutr. Sci. 2016, 66, 67–76. [Google Scholar] [CrossRef]
- Liang, Z.; Luo, Z.; Li, W.; Yang, M.; Wang, L.; Lin, X.; Li, L. Elevated CO2 enhanced the antioxidant activity and downregulated cell wall metabolism of wolfberry (Lycium barbarum L.). Antioxidants 2022, 11, 16. [Google Scholar] [CrossRef]
- Meng, J.; Liu, Z.; Gou, C.L.; Rogers, K.M.; Yu, W.-J.; Zhang, S.-S.; Yuan, Y.-W.; Zhang, L. Geographical origin of Chinese wolfberry (goji) determined by carbon isotope analysis of specific volatile compounds. J. Chromatogr. B 2019, 1105, 104–112. [Google Scholar] [CrossRef]
- Yin, T.; Guo, T.; Ma, Z.; Wang, Z.; Sun, X.; Li, C. Classification of wolfberry with different geographical origins by using voltametric electronic tongue. IFAC Papers OnLine 2018, 51, 654–659. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, Z.; Yuan, W.; Li, C.; Jing, X.; Han, H. Classification of wolfberry from different geographical origins by using electronic tongue and deep learning algorithm. IFAC Papers OnLine 2019, 52, 397–402. [Google Scholar] [CrossRef]
- Zhang, S.; Wei, Y.; Wei, S.; Liu, H.; Guo, B. Authentication of Zhongning wolfberry with geographical indication by mineral profile. Int. J. Food Sci. Technol. 2017, 52, 457–463. [Google Scholar] [CrossRef]
- Dong, F.; Hao, J.; Luo, R.; Zhang, Z.; Wang, S.; Wu, K. Identification of the proximate geographical origin of wolfberries by two-dimensional correlation spectroscopy combined with deep learning. Comput. Electron. Agric. 2022, 198, 107027. [Google Scholar] [CrossRef]
- Du, M.; Gong, Y.; Lin, Z.Z.; Shi, X.Y.; Hua, G.D.; Qiao, Y.J. Rapid identification of wolfberry fruit of different geographic regions with sample surface near infrared spectra combined with multi-class SVM. Spectrosc. Spect. Anal. 2013, 33, 1211–1214. [Google Scholar]
- Zhong, L.; Liu, M.D.; Ji, S.X. The identification of the origin of Chinese wolfberry based on infrared spectral technology and the artificial neural network. Spectrosc. Spect. Anal. 2016, 36, 720–723. [Google Scholar]
- Gong, H.; Rehman, F.; Li, Z.; Liu, J.; Yang, T.; Liu, J.; Li, H.; Hu, Z.; Ma, Q.; Wu, Z.; et al. Discrimination of geographical origins of wolfberry (Lycium barbarum L.) fruits using stable isotopes, earth elements, free amino acids, and saccharides. J. Agric. Food Chem. 2022, 70, 2984–2997. [Google Scholar] [CrossRef]
- Bondia-Pons, I.; Savolainen, O.; Törrönen, R.; Martinez, J.A.; Poutanen, K.; Hanhineva, K. Metabolic profiling of Gogi berry extracts for discrimination of geographical origin by non-targeted liquid chromatography couples to quadrupole time-of-flight mass spectrometry. Food Res. Int. 2014, 63, 132–138. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, Z.; Wei, S.; Du, G.; Ji, Q.; Zhang, L.; Wei, H. Study on the design and control system for wolfberry harvesting robot. In Proceedings of the 28th Chinese Control and Decision Conference, Yinchuan, China, 28–30 May 2016; pp. 5984–5985. [Google Scholar]
- Wang, J.; Mei, S.; Xiao, G.; Zhou, H. Research on mechanized harvesting methods of Lycium barbarum fruit. IFAC PapersOnLine 2018, 51, 223–226. [Google Scholar]
- Chen, Y.; Zhao, J.; Hu, G.; Chen, J. Design and testing of a pneumatic oscillating Chinese wolfberry harvester. Horticulturae 2021, 7, 214. [Google Scholar] [CrossRef]
- Ban, Z.; Wei, W.; Yang, X.; Feng, J.; Guan, J.; Li, L. Combination of heat treatment and chitosan coating to improve postharvest quality of wolfberry (Lycium barbarum). Int. J. Food Sci. Technol. 2015, 50, 1019–1025. [Google Scholar] [CrossRef]
- Xiang, W.; Wang, H.-W.; Tian, Y.; Sun, D.-W. Effects of salicylic acid combined with gas atmospheric control on postharvest quality and storage stability of wolfberries: Quality attributes and interaction evaluation. J. Food Process Eng. 2021, 44, e13764. [Google Scholar] [CrossRef]
- .Ling, L.; Zhao, Y.; Tu, Y.; Yang, C.; Ma, W.; Feng, S.; Lu, L.; Zhang, J. The inhibitory effect of volatile organic compounds produced by Bacillus subtilis CL2 on pathogenic fungi of wolfberry. J. Basic Microbiol. 2021, 61, 110–121. [Google Scholar] [CrossRef]
- Yang, M.; Ding, C.; Zhu, J. The drying quality and energy consumption of Chinese wolfberry fruits under electrohydrodynamic system. Int. J. Appl. Electromagn. Mech. 2017, 55, 101–112. [Google Scholar] [CrossRef]
- Yang, M.; Ding, C. Electrohydrodynamic (EHD) drying of the Chinese wolfberry fruits. SpringerPlus 2016, 5, 909. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Ding, C.; Zhang, Y.; Song, Z.; Hu, X.; Hao, T. Effect of electrohydrodynamic partially combined with oven drying on Chinese wolfberry. Int. J. Appl. Electromagn. Mech. 2020, 63, 465–482. [Google Scholar] [CrossRef]
- Ni, J.; Ding, C.; Zhang, Y.; Song, Z.; Hu, X.; Hao, T. Electrohydrodynamic drying of Chinese wolfberry in a multiple needle-to-plate electrode system. Foods 2019, 8, 152. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Zengh, Z.-A.; Mujumdar, A.S.; Fang, X.-M.; Wang, J.; Zhang, Q.; Ma, Q.; Xiao, H.-W.; Liu, Y.-H.; Gao, Z.-J. Pulsed vacum drying (PVD) of wolfberry: Drying kinetics and quality attributes. Drying Technol. 2018, 36, 1501–1514. [Google Scholar] [CrossRef]
- Yu, F.; Li, Y.; Wu, Z.; Wang, X.; Wan, N.; Yang, M. Dehydration of wolfberry fruit using pulsed vacuum drying combined with carboxymethyl celulose coating pretreatment. LWT–Food Sci. Technol. 2020, 134, 110159. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, S.; Chu, W.; He, W.; Yu, C.; Yu, H. Numerical analysis and preliminar experiment of a solar assisted heat pump drying system for Chinese wolfberry. Energies 2020, 13, 4306. [Google Scholar] [CrossRef]
- Xie, L.; Mujumdar, A.S.; Fang, X.-M.; Wang, J.; Dai, J.-W.; Du, Z.-L.; Xiao, H.-W.; Liu, Y.; Gao, Z.-J. Far-infrared radiation heating assisted pulsed vacuum drying (FIR-PVD) of wolfberry (Lycium barbarum L.): Effects on drying kinetics and quality attributes. Food Bioprod. Process. 2017, 102, 320–331. [Google Scholar] [CrossRef]
- Qi, Y.; Yu, F.; Wang, X.; Wan, N.; Yang, M.; Wu, Z.; Li, Y. Drying of wolfberry fruit juice using low-intensity pulsed ultrasound. LWT–Food Sci. Technol. 2021, 141, 110953. [Google Scholar] [CrossRef]
- Xu, Y.; Zang, Z.; Zhang, Q.; Wang, T.; Shang, J.; Huang, X.; Wan, F. Characteristics and quality analysis of radio frequency-hot air combined segmented drying of wolfberry (Lycium barbarum). Foods 2022, 11, 1645. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Zhao, D.; Huang, J.; Hao, J. Progress on research and development of goji berry drying: A review. Int. J. Food Propert. 2022, 25, 435–449. [Google Scholar] [CrossRef]
- Fan, J.; Lin, L.; Zhao, M. Construction of in vitro fermentation model using gut microbiota relating to glucose and lipid metabolism: A supplementary method for initial screening of polysaccharides with hypoglycemic potentials. J. Sci. Food Agric. 2022, 102, 6328–6339. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.-H.; Zhang, X.-X.; Ni, Z.-J.; Thakur, K.; Wang, W.; Yan, Y.-M.; Cao, Y.-L.; Zhang, J.-G.; Rengasamy, K.R.R.; Wei, Z.-J. Lycium barbarum (goji) as functional food: A review of its nutrition, phytochemical structure, biological features, and food industry prospects. Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jin, H.; Dong, X.; Yang, S.; Ma, S.; Ni, J. Quality evaluation of Lycium barbarum (wolfberry) from different regions in China based on polysaccharide structure, yield and bioactivies. Chin. Med. 2019, 14, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potterat, O. Goji (Lycium barbarum and L. chinense): Phytochemistry, pharmacology and safety in the perspective of traditional uses and recent popularity. Planta Med. 2010, 76, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.-H.; Zhang, X.-X.; Thakur, K.; Zhang, J.-G.; Wei, Z.-J. Research progress of Lycium barbarum L. as functional food: Phytochemical composition and health benefits. Curr. Opin. Food Sci. 2022, 47, 100871. [Google Scholar] [CrossRef]
- Gao, Y.; Wei, Y.; Wang, Y.; Gao, F.; Chen, Z. Lycium barbarum: A traditional Chinese herb and a promising anti-aging agent. Aging Dis. 2017, 8, 778–791. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Guo, S.; Zhou, J.; Zhu, Y.; Zhang, F.; Zeng, F.; Duan, R.; Xu, M.; Duan, J.-A. Chemical constituents from Lycium barbarum (Solanaceae) and their chemophenetic significance. Biochem. Syst. Ecol. 2021, 97, 104292. [Google Scholar] [CrossRef]
- Liu, J.; Gong, Y.; Yang, J.; Di, D. Advance on alkaloids of Lycium genus. Chin. Sci. Bull. 2022, 67, 332–350. [Google Scholar] [CrossRef]
- Vidović, B.B.; Milinčić, D.D.; Marčetić, M.D.; Dijuriš, J.D.; Ilić, T.D.; Kostić, A.Z.; Pešić, M.B. Health benefits and applications of goji berries in functional food products development: A review. Antioxidants 2022, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Wang, Q.Q.; Lei, Y.H.; Wang, S.S.; Chen, K.L.; Li, Y.; Xiong, J.; Liang, X.J.; Zhou, X.; Li, Y.K. Elucidating the interaction of rhizosphere bacteria and environmental factors in influencing active ingredient content of Lycium barbarum fruit in China. J. Appl. Microbiol. 2022, 132, 3783–3796. [Google Scholar] [CrossRef] [PubMed]
- Sim, R.H.; Sirasanagandla, S.R.; Das, S.; Teoh, S.L. Treatment of glaucoma with natural products and their mechanism of action: An update. Nutrients 2022, 14, 534. [Google Scholar] [CrossRef]
- Rang, Y.; Liu, H.; Liu, C. Potential for non-starch polysaccharides in the prevention and remediation of cognitive impairment: A comprehensive review. Int. J. Biol. Macromol. 2022, 208, 182–195. [Google Scholar] [CrossRef]
- Qi, Y.; Duan, G.; Fan, G.; Peng, N. Effect of Lycium barbarum polysaccharides on cell signal transduction pathways. Biomed. Pharmacother. 2022, 147, 112620. [Google Scholar] [CrossRef]
- Xiao, Z.; Deng, Q.; Zhou, W.; Zhang, Y. Immune activities of polysaccharides isolated from Lycium barbarum L. What do we know so far? Pharmacol. Therap. 2022, 229, 107921. [Google Scholar] [CrossRef]
- Huang, R.; Wu, E.; Deng, X. Potential of Lycium barbarum polysaccharide for the control of glucose and lipid metabolism disroders: A review. Int. J. Food Prop. 2022, 25, 673–680. [Google Scholar] [CrossRef]
- Pinilla, I.; Maneu, V.; Campello, L.; Fernández-Sánchez, L.; Martínez-Gil, N.; Kutsyr, O.; Sánchez-Sáez, X.; Sánchez-Castillo, C.; Lax, P.; Cuenca, N. Inherited retinal dystrophies: Role of oxidative stress and inflammation in their physiopathology and therapeutic implications. Antioxidants 2022, 11, 1086. [Google Scholar] [CrossRef]
- Yang, C.; Zhao, Q.; Li, S.; Pu, L.; Yu, L.; Liu, Y.; Lai, X. Effects of Lycium barbarum L.: Polysaccharides on vascular retinopathy: An insight review. Molecules 2022, 27, 5628. [Google Scholar] [CrossRef]
- Liu, H.; Cui, B.; Zhang, Z. Mechanism of glycometabolism regulation by bioactive compounds from the fruits of Lycium barbarum: A review. Food Res. Int. 2022, 159, 111408. [Google Scholar] [CrossRef] [PubMed]
- Rohilla, P.; Jain, H.; Chhikara, A.; Singh, L.; Dahiya, P. Anticancer potential of Solanaceae plants: A review. S. Afr. J. Bot. 2022, 149, 269–289. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, S.; Tan, H.; Yang, P.; Li, Y.; Xu, L.; Duan, B.; Liu, Y. Advances in polysaccharides of natural source of the anti-Alzheimer’s disease effect and mechanism. Carbohydr. Polym. 2022, 296, 119961. [Google Scholar] [CrossRef]
- Xu, W.; Jiang, Y.; Wang, N.; Bai, H.; Xu, S. Traditional Chinese medicine as a promising strategy for the treatment od Alzheimer’s disease complicated with osteoporosis. Front. Pharmacol. 2022, 13, 842101. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, C.; Shen, X.; Liu, Y. The use of traditional Chinese medicines in relieving exercise-induced fatigue. Front. Pharmacol. 2022, 13, 969827. [Google Scholar] [CrossRef] [PubMed]
- Che, Z.; Zhou, Z.; Xiao, J. Mechanisms of wolfberry in the treatment of liver diseases. Chin. Sci. Bull. 2022, 67, 351–363. [Google Scholar] [CrossRef]
- Ren, Y.; Li, S.; Song, Z.; Luo, Q.; Zhang, Y.; Wang, H. The regulatory roles of polysaccharides and ferroptosis-related phytochemicals in liver diseases. Nutrients 2022, 14, 2303. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, R.; Yang, Z.; Wen, Q.; Cao, X.; Zhao, N.; Yan, J. Protective effects of polysaccharides in neurodegenerative diseases. Front. Aging Neurosci. 2022, 14, 917629. [Google Scholar] [CrossRef]
- Feng, Y.; Song, Y.; Zhou, J.; Duan, Y.; Kong, T.; Ma, H.; Zhang, H. Recent progress of Lycium barbarum polysaccharides on intestinal microbiota, microbial metabolites and health: A review. Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef]
- Cao, C.; Wang, Z.; Gong, G.; Huang, W.; Huang, L.; Song, S.; Zhu, B. Effects of Lycium barbarum polysaccharides on immunity and metabolic syndrome associated with the modulation of gut microbiota: A review. Foods 2022, 11, 3177. [Google Scholar] [CrossRef]
- Wang, B.; Han, L.; Liu, J.-M.; Zhang, J.; Wang, W.; Li, B.-G.; Dong, C.-X.; Bai, C.-C. Lycium genus polysaccharides: An overview of its extraction, structures, pharmacological activities and biological applications. Separations 2022, 9, 197. [Google Scholar] [CrossRef]
- Gong, H.; Li, W.; Sun, J.; Guan, Q.; Guo, Y.; Wang, Y. A review on plant polysaccharide based on drug delivery system for construction and application, with emphasis on traditional Chinese medicine polysaccharide. Int. J. Biol. Macromol. 2022, 211, 711–728. [Google Scholar] [CrossRef] [PubMed]
- Wannes, W.A.; Tounsi, M.S. Phytochemical composition and health properties of Lycium europaeum L.: A review. Acta Ecol. Sinica 2021, 41, 390–401. [Google Scholar] [CrossRef]
- Yao, X.; Peng, Y.; Xu, L.-J.; Li, L.; Wu, Q.-L.; Xiao, P.-G. Phytochemical and biological studies of Lycium medicinal plants. Chem. Biodivers. 2011, 8, 976–1010. [Google Scholar] [CrossRef] [PubMed]
- Akter, F.; Rahman, M.S.; Al Amin, G.M.; Miah, M.I.; Koh, Y.-S. Beneficial therapy with natural anti-inflammatory agents and supplements. J. Bacteriol. Virol. 2021, 51, 149–162. [Google Scholar] [CrossRef]
- Ikanya, L.W.; Maina, J.G.; Gachuiri, C.K.; Owino, W.O.; Dubeux, J.C.B., Jr. Chemical composition, and digestibility of preferred forage species by lactating Somali camels in Kenya. Rangel. Ecol. Manag. 2022, 80, 61–67. [Google Scholar] [CrossRef]
- Chaachouay, N.; Azeroual, A.; Bencharki, B.; Douira, A.; Zidane, L. Ethnoveterinary medicinal plants for animal therapy in the Rif, North of Morocco. S. Afr. J. Bot. 2022, 147, 176–191. [Google Scholar] [CrossRef]
- Mollel, N.P.; Otieno, J.N.; Sitoni, D.K. Medicinal plants traded in Arusha city, Tanzania. J. Med. Plants Stud. 2022, 10, 175–182. [Google Scholar]
- El-Mokasabi, F. Survey of wild trees and shrubs in Eastern region of Libya and their economical value. Alq. J. Med. App. Sci. 2022, 5, 48–55. [Google Scholar]
- Almushghub, F.; Ahmed, D.; El-Din, A.S.; Shaltout, K. Vegetation analysis of Wadi Kaam at northwest Libya. J. Basic Environ. Sci. 2022, 9, 20–37. [Google Scholar]
- Bendjedou, H.; Bennaceura, M.; Benamar, H.; Rodrigues, M.J.; Pereira, C.; Bensouicie, C.; Custódio, L. In vitro antioxidant and enzyme inhibitory properties and phenolic contents of crude extracts and fractions from different organs of the halophyte Lycium europaeum L. Curr. Bioact. Compd. 2022, 18, 2. [Google Scholar] [CrossRef]
- Mejri, H.; Ouerghemi, I.; Wannes, W.A.; Haddada, F.M.; Tlili, N.; Hammami, M.; Dussault, C.; Lancette, K.G.-L.; Pichette, A.; Legault, J.; et al. Phytochemical analysis, antioxidant, anticancer and anti-inflammatory activities of Lycium europaeum fruits. Int. J. Environ. Health Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Khodaparast, S.A.; Darsaraei, H.; Abbasi, M. The genus Arthrocladiella: A new report of powdery mildew fungi from Iran. Mycol. Iran. 2021, 8, 135–140. [Google Scholar]
- Ajjoun, M.; Kharchoufa, L.; Merrouni, I.A.; Elachouri, M. Moroccan medicinal plants traditionally used for the treatment of skin diseases: From ethnobotany to clinical trials. J. Ethnopharmacol. 2022, 297, 115532. [Google Scholar] [CrossRef]
- Jaadan, H.; Akodad, M.; Moumen, A.; Baghour, M.; Skalli, A.; Ezrari, S.; Belmalha, S. Ethnobotanical survey of medicinal plants growing in the region of “Oulad Daoud Zkhanine” (Nador Province), in Northeastern Morocco. Ethnobot. Res. Appl. 2020, 19, 39. [Google Scholar] [CrossRef]
- Montoleone, E.; Longo, D.; Cassanego, E.; Lazzeri, V. Lycium schweinfurthii Dammer (Solanaceae, Lycieae), species mal compressa confermata per meridione d’Italia. Acta Plant. Notes 2022, 8, 11–16. [Google Scholar]
- Jamous, R.M.; Zaitoun, S.Y.A.; Husein, A.I.; Qasem, I.B.Y.; Ali-Shtayeh, M.S. Screening for biological activities of medicinal plants used in traditional Arabic Palestinian herbal medicine. Eur. J. Med. Plants 2015, 9, 1–13. [Google Scholar] [CrossRef]
- Auda, M.A. An ethnobotanical uses of plants in the Middle Area, Gaza Strip, Palestine. Adv. Environ. Biol. 2011, 5, 3681–3687. [Google Scholar]
- Ighbareyeh, J.M.H.; Cano-Ortiz, A.; Cano, E. Phytosociology and vegetation of plants of Beit Jibrin in Palestine. Land 2022, 11, 264. [Google Scholar] [CrossRef]
- Baghdadi, H.; El-Sayed, S.H.; Salem, G.; Metwally, A.; Salem, A.M. A comparative chemical study of Lycium species growing in Egypt. Alex. J. Pharm. Sci. 1988, 2, 73–76. [Google Scholar]
- Ewais, E.A.; Abd El-Maboud, M.M.; Elhaw, M.H.; Haggag, M.I. Phytochemical studies on Lycium schweinfurthii var. schweinfurthii (Solanaceae) and isolation of five flavonoids from leaves. J. Med. Plants Stud. 2016, 4, 288–300. [Google Scholar]
- Elbermawi, A.; Halim, A.F.; Mansour, E.-S.S.; Ahmad, K.F.; Ashour, A.; Amen, Y.; Shimizu, K. A new glucoside with a potent α-glucosidase inhibitory activity from Lycium schweinfurthii. Nat. Prod. Res. 2021, 35, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Elbermawi, A.; Halim, A.F.; Mansour, E.-S.S.; Ahmad, K.F.; Elsbaey, M.; Ashour, A.; Amen, Y.; El-Gamil, M.M.; Tomofumi, M.; Shimizu, K. Lycium schweinfurthii: New secondary metabolites and their cytotoxic activities. Nat. Prod. Res. 2022, 36, 5134–5141. [Google Scholar] [CrossRef] [PubMed]
- Bedair, H.; Shaltout, K.; Ahmed, D.; El-Din, A.S.; El-Fahhar, R. Characterization of the wild trees and shrubs in the Egyptian flora. Egypt. J. Bot. 2020, 60, 147–168. [Google Scholar] [CrossRef]
- Mamdouh, D.; Mahgoub, H.A.M.; Gabr, A.M.M.; Ewais, E.A.; Smetanska, I. Genetic stability, phenolic, flavonoid, ferulic acid contents, and antioxidant activity of micropropagated Lycium schweinfurthii plants. Plants 2021, 10, 2089. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Kerbler, S.M.; Fernie, A.R.; Zhang, Y. Plant cell cultures as heterologous bio-factories for secondary metabolite production. Plant Commun. 2021, 2, 100235. [Google Scholar] [CrossRef]
- Mamdouh, D.; Smetanska, I. Optimization of callus and cell suspension cultures of Lycium schweinfurthii for improved production of phenolics, flavonoids, and antioxidant activity. Horticulturae 2022, 8, 394. [Google Scholar] [CrossRef]
- Elbermawi, A.; Ali, A.R.; Amen, Y.; Ashour, A.; Ahmad, K.F.; Mansour, E.-S.S.; Halim, A.F. Anti-diabetic activities of phenolic compounds of Alternaria sp., an endophyte isolated from the leaves of desert plants growing in Egypt. RSC Adv. 2022, 12, 24935. [Google Scholar] [CrossRef]
- Chen, A.; Mao, X.; Sun, Q.; Wei, Z.; Li, J.; You, Y.; Zhao, J.; Jiang, G.; Wu, Y.; Wang, L.; et al. Alternaria mycotoxins: An overview of toxicity, metabolism, and analysis in food. J. Agric. Food Chem. 2021, 69, 7817–7830. [Google Scholar] [CrossRef]
- Zhang, W.; Krohn, K.; Egold, H.; Draeger, S.; Schulz, B. Diversity of antimicrobial pyrenophorol derivatives from an endophytic fungus, Phoma sp. Eur. J. Org. Chem. 2008, 2008, 4320–4328. [Google Scholar] [CrossRef]
- Krohn, K.; Kouam, S.F.; Kuigoua, G.M.; Hussain, H.; Cludius-Brandt, S.; Flörke, U.; Kurtán, T.; Pescitelli, G.; di Bari, L.; Draeger, S.; et al. Xanthones and oxepino[2,3-b]chromones from three endophytic fungi. Chem. Eur. J. 2009, 15, 12121–12132. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, I.N.; Zahoor, A.; Hussain, H.; Ahmed, I.; Ahmad, V.U.; Padula, D.; Draeger, S.; Schulz, B.; Meier, K.; Steiner, M.; et al. Diversonol and blennolide derivatives from the endophytic fungus Microdiplodia sp.: Absolute configuration of diversonol. J. Nat. Prod. 2011, 74, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, E.; Bancheva, S.; Domina, G.; Venturella, G. Distribution, ecological role and symbioses of selected shrubby species in the Mediterranean basin: A review. Plant Biosyst. 2020, 154, 438–454. [Google Scholar] [CrossRef]
- Obón, C.; Rivera, D.; Verde, A.; Alcaraz, F. Ethnopharmacology and medicinal uses of extreme halophytes. In Handbook of Halophytes; Grigore, M.N., Ed.; Springer: Cham, Switzerland, 2020; pp. 1–27. [Google Scholar] [CrossRef]
- Marrero-Rodríguez, N.; García-Romero, L.; Hrnández-Cordero, A.I.; Peña-Alonso, C.; Pérez-Chacón Espino, E. Deforestation by historical lime industry in an arid aeolian sedimentary system: An applied and methodological research. Sci. Total Environ. 2022, 819, 152009. [Google Scholar] [CrossRef] [PubMed]
- Midhat, L.; Ouazzani, N.; Hejjaj, A.; Ouhammou, A.; Mandi, L. Accumulation of heavy metals in metallophytes from three mining sites (Southern Centre Morocco) and evaluation of their phytoremediation potential. Eotoxicol. Environ. Saf. 2019, 169, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Idm’hand, E.; Msanda, F.; Cherifi, K. Ethnobotanical study and biodiversity of medicinal plants used in the Tarfaya Province, Morocco. Acta Ecol. Sin. 2020, 40, 134–144. [Google Scholar] [CrossRef]
- Fatiha, B.A.; Souad, S.; Ouafe, B.; Jamila, D.; Allal, D.; Lahcen, Z. Ehnobotanical study of medicinal plants used in the region of Middle Oumrbia (Morocco). Plant Arch. 2019, 19, 2005–2017. [Google Scholar]
- Abdennacer, B.; Karim, M.; Yassine, M.; Nesrine, R.; Mouna, D.; Mohamed, B. Determination of phytochemicals and antioxidante acivity of mthanol extracts obtained from the fruit and leaves of Tunisian Lycium intricatum Boiss. Food Chem. 2015, 174, 577–584. [Google Scholar] [CrossRef]
- Jiang, Y.; Fang, Z.; Leonard, W.; Zhang, P. Phenolic compounds in Lycium berry: Composition, health benefits and industrial applications. J. Funct. Foods 2021, 77, 104340. [Google Scholar] [CrossRef]
- Boulila, A.; Bejaoui, A. Lycium intricatum Boiss.: An unexploited and rich source of unsaturated fatty acids, 4-desmethylsterols and other valuable phytochemicals. Lipids Heath Dis. 2015, 14, 59. [Google Scholar] [CrossRef] [Green Version]
- Bendjedou, H.; Maggi, F.; Bennaceur, M.; Mancinelli, M.; Benamar, H.; Barboni, L. A new ionone derivative from Lycium intricatum Boiss. (Solanaceae). Nat. Prod. Res. 2022, 36, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, R.; Paul, S.; Tayung, K. Antifungal drugs from endophytic microbes: Present and future prospects. In Bioresource Utilization and Management. Applications in Therapeutics, Biofuels, Agriculture, and Environmental Science; Thatoi, H., Das, S.K., Mohapatra, S., Eds.; Apple Acacemic Press: Palm Bay, FL, USA, 2022; pp. 3–37. [Google Scholar]
- Khattab, A.R.; Farag, M.A. Marine and terrestrial endophytic fungi: A mine of bioactive xanthone compounds, recent progress, limitations, and novel applications. Crit. Rev. Biotechnol. 2022, 42, 403–430. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Singh, S.P. Role of the endogenous fungal metabolites in the plant growth improvement and stress. Fungi Bio-Prospects Sustain. Agric. Environ. Nano-Technol. 2021, 3, 381–401. [Google Scholar] [CrossRef]
- Qian, D.; Zhao, Y.; Yang, G.; Huang, L. Systematic review of chemical constituents in the genus Lycium (Solanaceae). Molecules 2017, 22, 911. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Cell Line | Assay | IC50 (μM) | Reference |
|---|---|---|---|---|
| Diosmetin | NT | Inhibition of α-glucosidase | 23.55 | [83] |
| Quercetin | NT | Inhibition of α-glucosidase | 21.38 | [83] |
| Diosmetin | NT | Inhibition of α-glucosidase | 7.12 | [83] |
| Diosmetin-7-O-β-D-glucoside | NT | Inhibition of α-glucosidase | 168.85 | [83] |
| 3-Methoxy-4-O-β-D-glucopyranosyl-methylbenzoate | NT | Inhibition of α-glucosidase | 22.76 | [83] |
| 3-Hydroxy-1-(4-hydroxy-3-methoxyphenyl)-2[4-(3-hydroxy-1-(E)-propenyl)-2-methoxyphenoxy]-propyl-β-D-glucopyranoside | NT | 181.39 | [83] | |
| Vaginatin | 1/2/3 | Cytotoxicity | 25.65/82.91/69.18 | [84] |
| 11S-Methoxy-11,12-dihydrophytuberin | 1/2/3 | Cytotoxicity | 92.09/>100/>100 | [84] |
| (E)-Docosanoyl ferulate | 1/2/3 | Cytotoxicity | 73.49/>100/>100 | [84] |
| (Z)-Docosanoyl ferulate | 1/2/3 | Cytotoxicity | >100/>100>100 | [84] |
| β-Sitosterol | 1/2/3 | Cytotoxicity | NT/>100/>100 | [84] |
| β-Sitosterol-3-O-β-D-glucoside | 1/2/3 | Cytotoxicity | 53.19/>100>96.19 | [84] |
| 9S-Methoxy-benzocyclononan-7-one | 1/2/3 | Cytotoxicity | 47.34/>100/>100 | [84] |
| Isoscopoletin | 1/2/3 | Cytotoxicity | 31.90/>100/>100 | [84] |
| Pinoresinol | 1/2/3 | Cytotoxicity | 34.57/>100/75.76 | [84] |
| trans-Cinnamoyl tyramine | 1/2/3 | Cytotoxicity | 45.58/>100/>100 | [84] |
| trans-Ferulyl tyramine | 1/2/3 | Cytotoxicity | 58.76/>100/90.26 | [84] |
| trans-Ferulyl-3-methoxytyramine | 1/2/3 | Cytotoxicity | 7.14/63.39/51.74 | [84] |
| Diosmetin | 1/2/3 | Cytotoxicity | 42.17/40.24/40.57 | [84] |
| Apigenin | 1/2/3 | Cytotoxicity | 54.49/>100/84.32 | [84] |
| Luteolin | 1/2/3 | Cytotoxicity | NT/>100/>100 | [84] |
| p-Hydroxybenzoic acid | 1/2/3 | Cytotoxicity | NT/>100/>100 | [84] |
| Vanillic acid | 1/2/3 | Cytotoxicity | 9.55/62.35/61.68 | [84] |
| Kampferol | 1/2/3 | Cytotoxicity | 49.10/>100/>100 | [84] |
| Quercetin | 1/2/3 | Cytotoxicity | NT/NT/NT | [84] |
| Methyl-α-D-fructofuranoside | 1/2/3 | Cytotoxicity | NT/NT/NT | [84] |
| Meliasendanin D | 1/2/3 | Cytotoxicity | >100/>100/>100 | [84] |
| trans-Ferulic acid | 1/2/3 | Cytotoxicity | NT/>100/>100 | [84] |
| p-Coumaric acid | 1/2/3 | Cytotoxicity | NT/>100/>100 | [84] |
| Diosmetin-7-O-β-D-glucoside | 1/2/3 | Cytotoxicity | NT/>100/>100 | [84] |
| Rutin | 1/2/3 | Cytotoxicity | 89.47//>100/>100 | [84] |
| 3-Methoxy-4-O-β-D-glucopyranosyl-methylbenzoate | 1/2/3 | Cytotoxicity | NT/>100/86.76 | [84] |
| Gallic acid | 1/2/3 | Cytotoxicity | 11.09/>100/75.92 | [84] |
| 3-Hydroxy-1-(4-hydroxy-3-methoxyphenyl)-2[4-(3-hydroxy-1-(E)-propenyl)-2-methoxyphenoxy]-propyl-β-D-glucopyranoside | 1/2/3 | Cytotoxicity | NT/>100/>100 | [84] |
| Inhibition of Enzymatic Activity | |||
|---|---|---|---|
| Compound/Endophytic Fungus | α-Glucosidase IC50 (μM) | Pancreatic Lipase IC50 (μM) | References |
| Alternaria sp. | Lycium schweinfurthii | ||
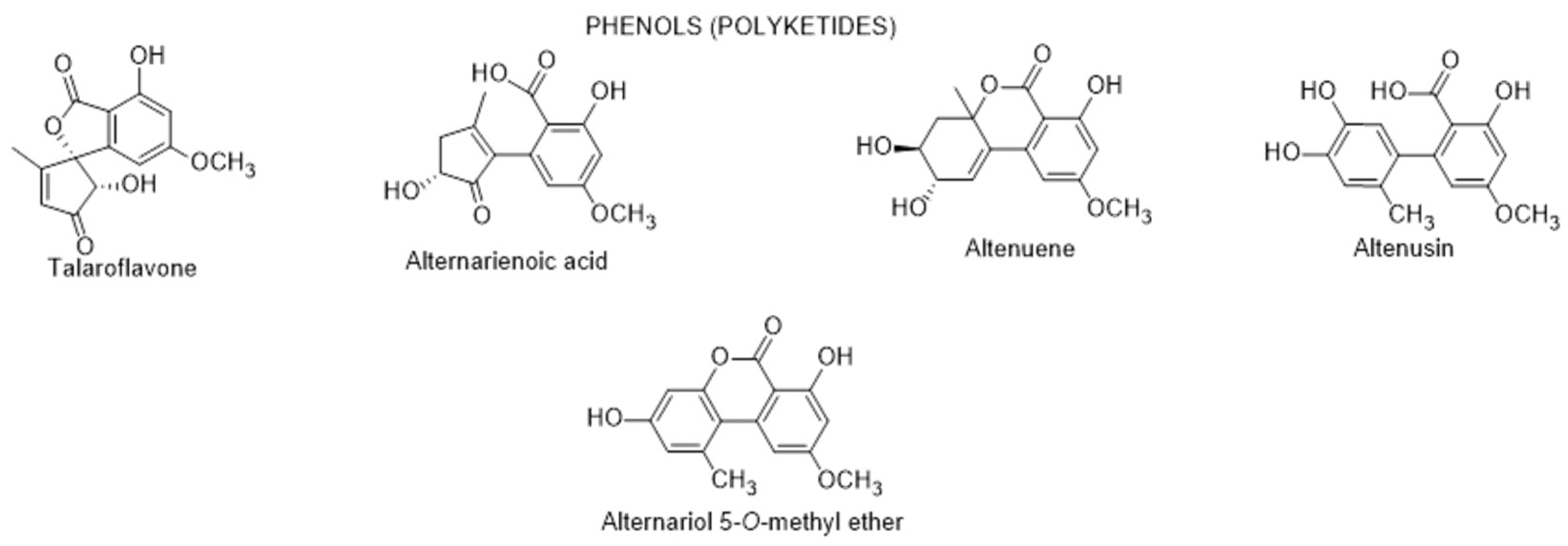
| Talaroflavone | >300 | 73.82 | [89] |
| Alternarienoic acid | 7.95 | 20.82 | [89] |
| Altenuene | 40.38 | 3.18 | [89] |
| Altenusin, | 46.14 | 21.46 | [89] |
| Alternariol | 179.88 | 56.85 | [89] |
| Alternariol-5-O-methyl ether | 236.25 | 7.60 | [89] |
| Compound/Endophytic Fungus | Antimicrobial Activity | ||
| Phoma sp. | Lycium intricatum | ||
| Microbotryum violaceum * | |||
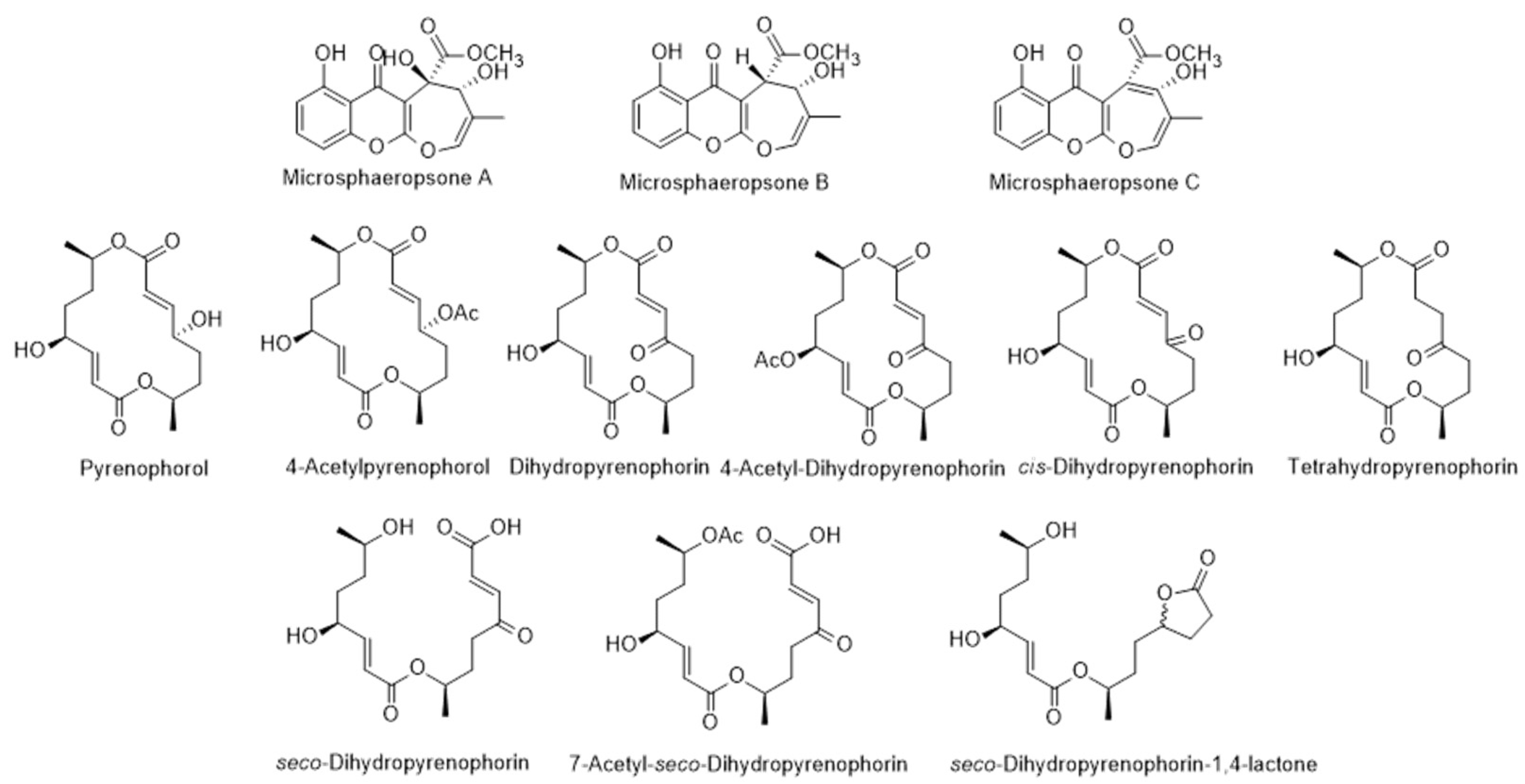
| Pyrenophorol | 7 | [91] | |
| Dihydropyrenophorin | 7 | [91] | |
| 4-Acetylpyrenophorol 4-Acetyldihydropyrenophorin | 10 7 | [91] [91] | |
| cis-Dihydro-pyrenophorin | 7 | [91] | |
| Tetrahydropyrenophorin | 10 | [91] | |
| seco-Dihydropyrenophorin | 10 | [91] | |
| 7-acetyl seco-dihydropyrenophorin | 9 | [91] | |
| seco-Dihydropyrenophorin-1,4-lactone | 7 | [91] | |
| Microsphaeropsis sp. | Lycium intricatum | ||
| Escherichia coli * | Bacillus megaterium * | ||
| Microsphaeropsone A | 8 | 10 | [92] |
| Microsphaeropsone C | 6 | 10 | [92] |
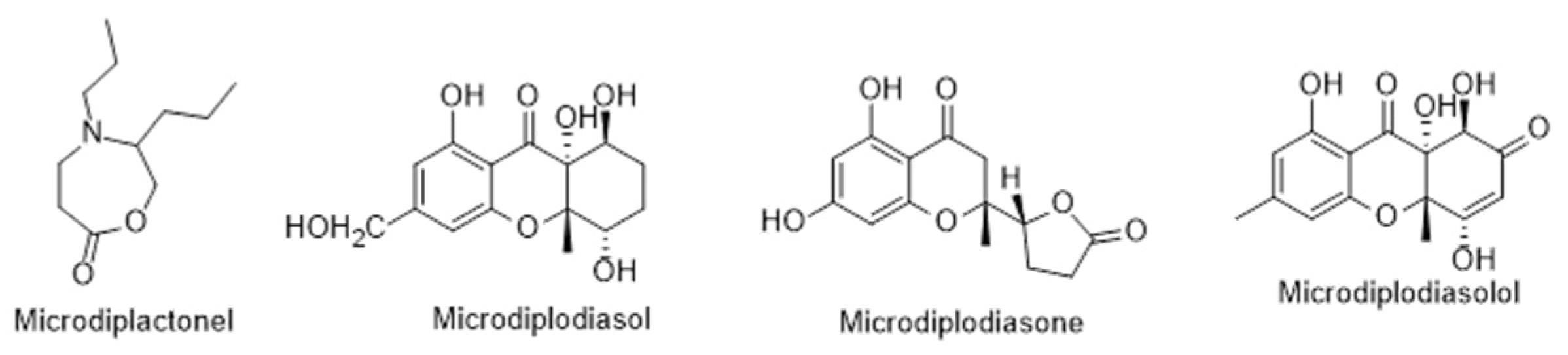
| Microdiplodia sp. | Lycium intricatum | ||
| Microbotryum violaceum * | Legionella pneumophila ** | ||
| Microdiplodiasol | 7 | 0.125–0.25 | [93] |
| Microdiplodiasone | 0 | 1.0 | [93] |
| Microdiplodiasolol | 6 | 1.0 | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miguel, M.d.G. Chemical and Biological Properties of Three Poorly Studied Species of Lycium Genus—Short Review. Metabolites 2022, 12, 1265. https://doi.org/10.3390/metabo12121265
Miguel MdG. Chemical and Biological Properties of Three Poorly Studied Species of Lycium Genus—Short Review. Metabolites. 2022; 12(12):1265. https://doi.org/10.3390/metabo12121265
Chicago/Turabian StyleMiguel, Maria da Graça. 2022. "Chemical and Biological Properties of Three Poorly Studied Species of Lycium Genus—Short Review" Metabolites 12, no. 12: 1265. https://doi.org/10.3390/metabo12121265