
Supplementation of Methyl-Donor Nutrients to a High-Fat, High-Sucrose Diet during Pregnancy and Lactation Normalizes Circulating 25-Dihydroxycholecalciferol Levels and Alleviates Inflammation in Offspring

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Design
2.2. Analysis of Serum 25D Concentrations
2.3. Analysis of Proinflammatory Cytokines
2.4. RNA Extraction and Real-Time PCR Analysis
2.5. Microbiome Analysis
2.6. Statistical Analysis
3. Results
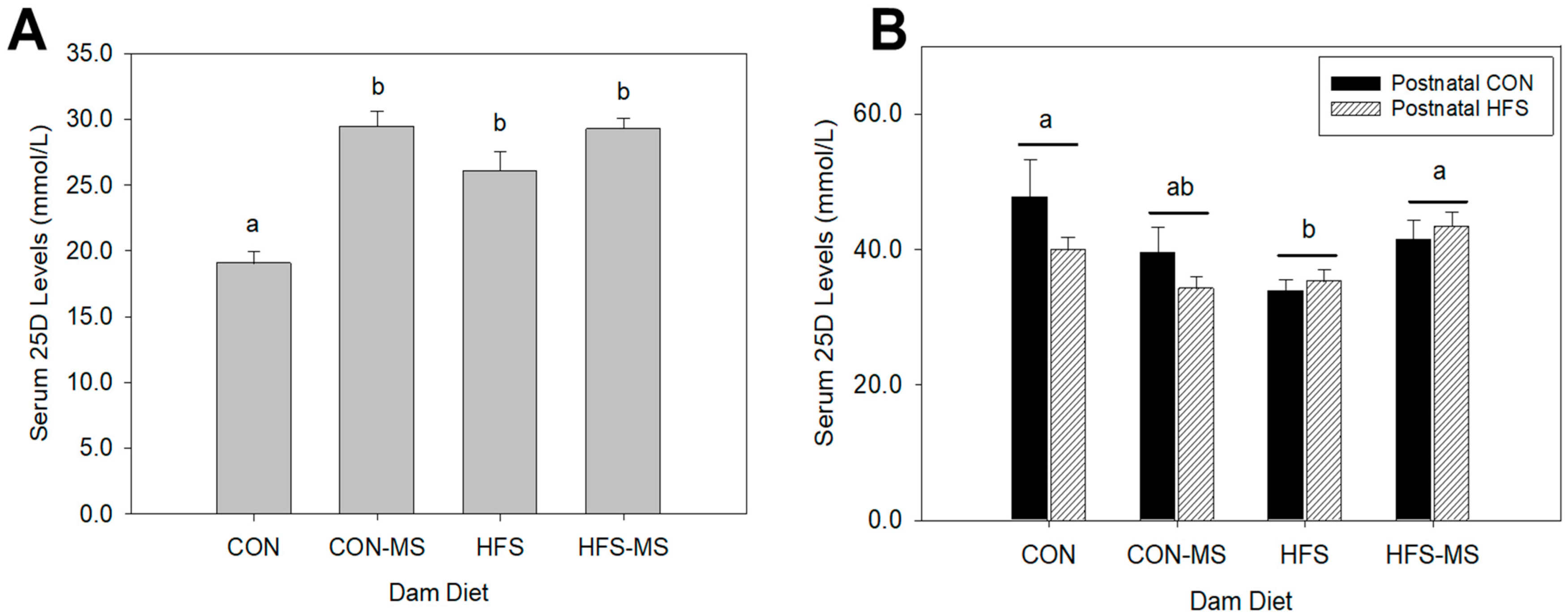
3.1. MS Supplementation during Pregnancy and Lactation Restores Perturbation of Vitamin D Status in Adult Offspring Induced by Maternal HFS Diet
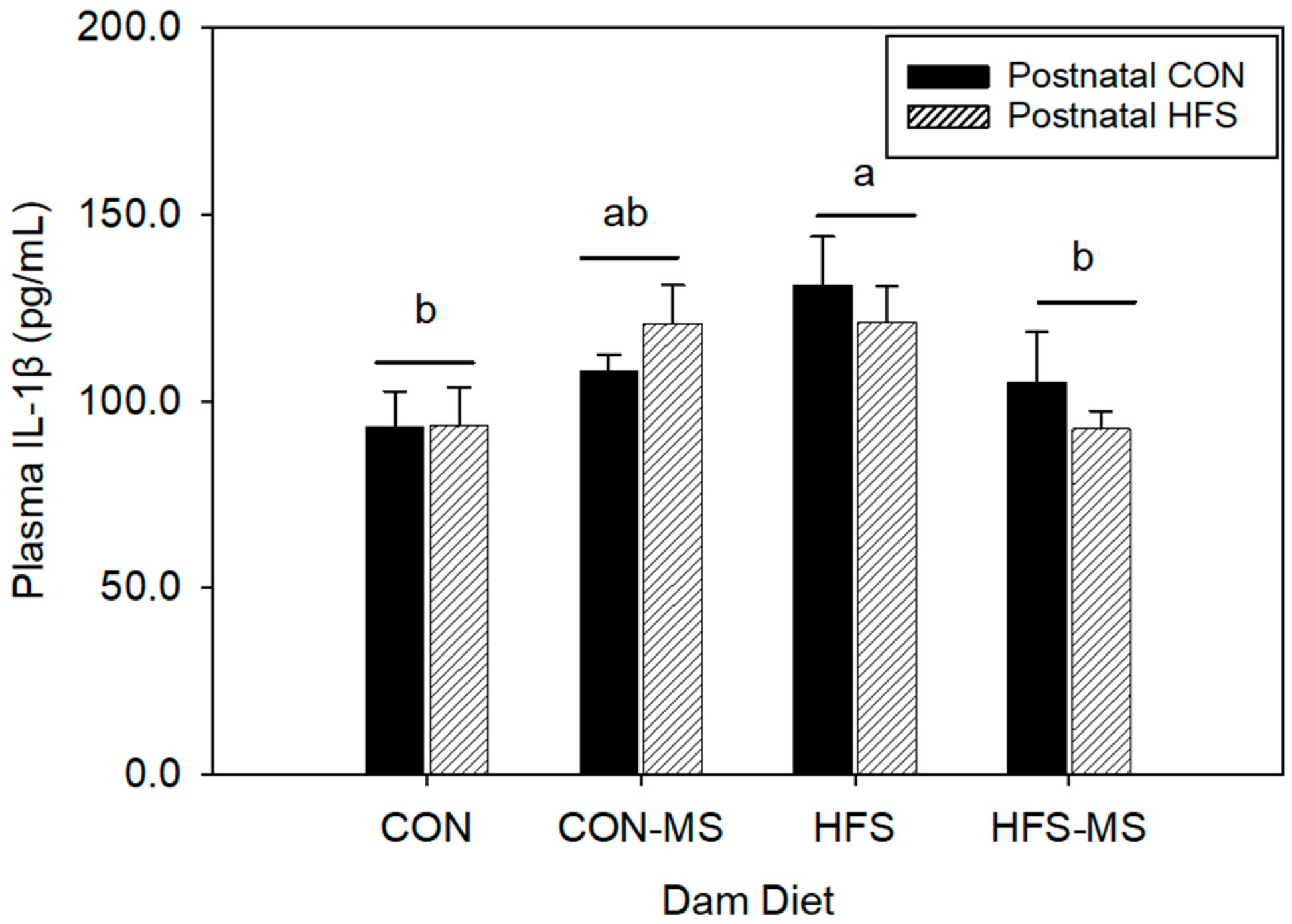
3.2. MS Supplementation during Pregnancy and Lactation Attenuates Maternal HFS-Induced Inflammation in Adult Pups
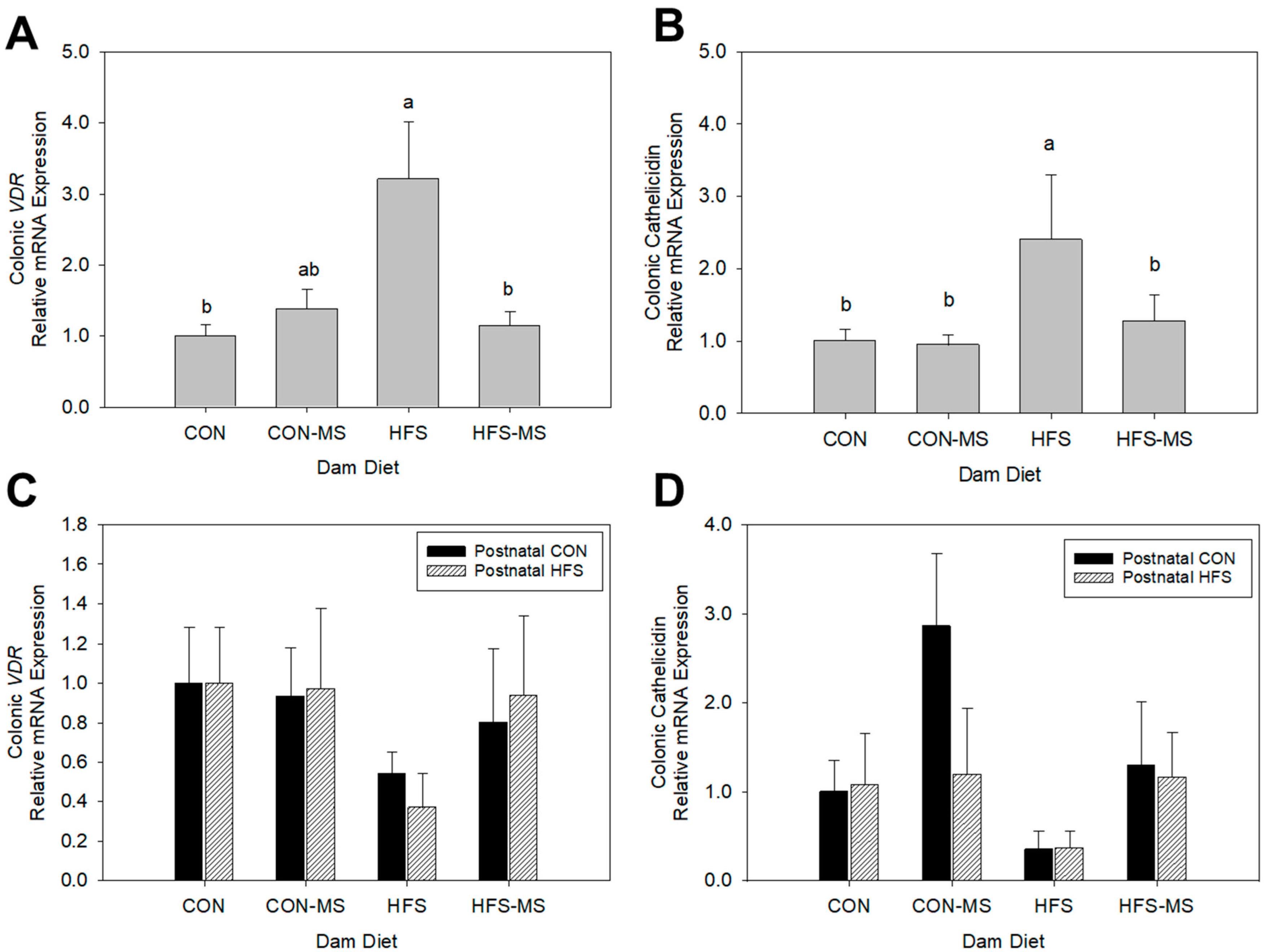
3.3. Maternal Diets Differentially Altered Colonic Vitamin D Signaling in Pups at Weaning and Adulthood
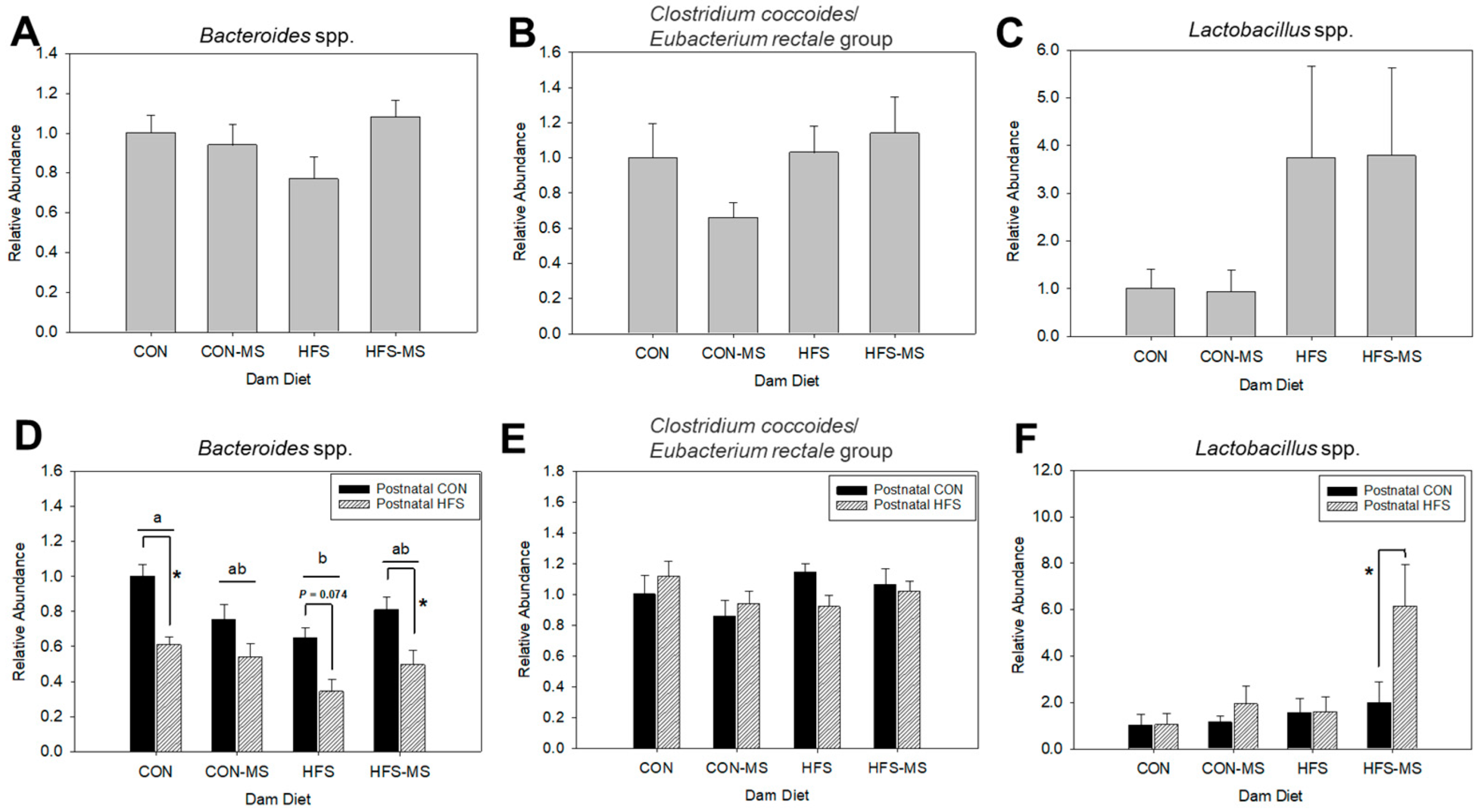
3.4. Prenatal and Postnatal Diets Together Modulate the Abundance of Bacteroides spp. and Lactobacillus spp. in Adult Pups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsoi, M.F.; Li, H.L.; Feng, Q.; Cheung, C.L.; Cheung, T.T.; Cheung, B.M.Y. Prevalence of Childhood Obesity in the United States in 1999–2018: A 20-Year Analysis. Obes. Facts 2022, 15, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Tylavsky, F.A.; Kocak, M.; Fowke, J.H.; Han, J.C.; Davis, R.L.; LeWinn, K.Z.; Bush, N.R.; Sathyanarayana, S.; Karr, C.J.; et al. Effects of Maternal Dietary Patterns during Pregnancy on Early Childhood Growth Trajectories and Obesity Risk: The CANDLE Study. Nutrients 2020, 12, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, J.G.; Bellissimo, C.J.; Yeo, E.; Fei Xia, Y.; Petrik, J.J.; Surette, M.G.; Bowdish, D.M.E.; Sloboda, D.M. Obesity during pregnancy results in maternal intestinal inflammation, placental hypoxia, and alters fetal glucose metabolism at mid-gestation. Sci. Rep. 2019, 9, 17621. [Google Scholar] [CrossRef]
- Gohir, W.; Kennedy, K.M.; Wallace, J.G.; Saoi, M.; Bellissimo, C.J.; Britz-McKibbin, P.; Petrik, J.J.; Surette, M.G.; Sloboda, D.M. High-fat diet intake modulates maternal intestinal adaptations to pregnancy and results in placental hypoxia, as well as altered fetal gut barrier proteins and immune markers. J. Physiol. 2019, 597, 3029–3051. [Google Scholar] [CrossRef] [PubMed]
- Francis, E.C.; Dabelea, D.; Boyle, K.E.; Jansson, T.; Perng, W. Maternal Diet Quality Is Associated with Placental Proteins in the Placental Insulin/Growth Factor, Environmental Stress, Inflammation, and mTOR Signaling Pathways: The Healthy Start ECHO Cohort. J. Nutr. 2022, 152, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Josefson, J.L.; Feinglass, J.; Rademaker, A.W.; Metzger, B.E.; Zeiss, D.M.; Price, H.E.; Langman, C.B. Maternal obesity and vitamin D sufficiency are associated with cord blood vitamin D insufficiency. J. Clin. Endocrinol. Metab. 2013, 98, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Mata-Greenwood, E.; Huber, H.F.; Li, C.; Nathanielsz, P.W. Role of pregnancy and obesity on vitamin D status, transport, and metabolism in baboons. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E63–E72. [Google Scholar] [CrossRef] [PubMed]
- Meeker, S.M.; Seamons, A.; Treuting, P.M.; Paik, J.; Brabb, T.; Hsu, C.C.; Grady, W.M.; Maggio-Price, L. Effect of Chronic Vitamin D Deficiency on the Development and Severity of DSS-Induced Colon Cancer in Smad3−/− Mice. Comp. Med. 2020, 70, 120–130. [Google Scholar] [CrossRef]
- Larriba, M.J.; Ordonez-Moran, P.; Chicote, I.; Martin-Fernandez, G.; Puig, I.; Munoz, A.; Palmer, H.G. Vitamin D receptor deficiency enhances Wnt/beta-catenin signaling and tumor burden in colon cancer. PLoS ONE 2011, 6, e23524. [Google Scholar] [CrossRef] [Green Version]
- Garg, M.; Hendy, P.; Ding, J.N.; Shaw, S.; Hold, G.; Hart, A. The Effect of Vitamin D on Intestinal Inflammation and Faecal Microbiota in Patients with Ulcerative Colitis. J. Crohns Colitis 2018, 12, 963–972. [Google Scholar] [CrossRef]
- Ooi, J.H.; Li, Y.; Rogers, C.J.; Cantorna, M.T. Vitamin D regulates the gut microbiome and protects mice from dextran sodium sulfate-induced colitis. J. Nutr. 2013, 143, 1679–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Yoon, S.; Zhang, Y.G.; Lu, R.; Xia, Y.; Wan, J.; Petrof, E.O.; Claud, E.C.; Chen, D.; Sun, J. Vitamin D receptor pathway is required for probiotic protection in colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G341–G349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, R.W.; Clemente, J.C.; Peter, I.; Loos, R.J.F. The prenatal gut microbiome: Are we colonized with bacteria in utero? Pediatr. Obes. 2017, 12 (Suppl. S1), 3–17. [Google Scholar] [CrossRef] [Green Version]
- Cortese, R.; Lu, L.; Yu, Y.; Ruden, D.; Claud, E.C. Epigenome-Microbiome crosstalk: A potential new paradigm influencing neonatal susceptibility to disease. Epigenetics 2016, 11, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.; Wu, S.; Zhang, Y.G.; Lu, R.; Xia, Y.; Dong, H.; Sun, J. Lack of Vitamin D Receptor Causes Dysbiosis and Changes the Functions of the Murine Intestinal Microbiome. Clin. Ther. 2015, 37, 996–1009.e1007. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Castano, G.P.; Dorris, M.R.; Liu, X.; Bolling, B.W.; Acosta-Gonzalez, A.; Rey, F.E. Bacteroides thetaiotaomicron Starch Utilization Promotes Quercetin Degradation and Butyrate Production by Eubacterium ramulus. Front. Microbiol. 2019, 10, 1145. [Google Scholar] [CrossRef] [PubMed]
- Eloe-Fadrosh, E.A.; Brady, A.; Crabtree, J.; Drabek, E.F.; Ma, B.; Mahurkar, A.; Ravel, J.; Haverkamp, M.; Fiorino, A.M.; Botelho, C.; et al. Functional dynamics of the gut microbiome in elderly people during probiotic consumption. mBio 2015, 6, e00231-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.M.; Jeong, J.J.; Woo, K.H.; Han, M.J.; Kim, D.H. Lactobacillus sakei OK67 ameliorates high-fat diet-induced blood glucose intolerance and obesity in mice by inhibiting gut microbiota lipopolysaccharide production and inducing colon tight junction protein expression. Nutr. Res. 2016, 36, 337–348. [Google Scholar] [CrossRef]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhong, X.; Liu, X.; Wang, X.; Gao, X. Therapeutic and Improving Function of Lactobacilli in the Prevention and Treatment of Cardiovascular-Related Diseases: A Novel Perspective From Gut Microbiota. Front. Nutr. 2021, 8, 693412. [Google Scholar] [CrossRef]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, C.L.; Wu, J.M.; Chen, K.Y.; Wu, M.H.; Yang, P.J.; Lee, P.C.; Chen, P.D.; Yeh, S.L.; Lin, M.T. Effects of Different Routes and Forms of Vitamin D Administration on Mesenteric Lymph Node CD4+ T Cell Polarization and Intestinal Injury in Obese Mice Complicated with Polymicrobial Sepsis. Nutrients 2022, 14, 3557. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.; Mach, N. Role of Vitamin D in the Hygiene Hypothesis: The Interplay between Vitamin D, Vitamin D Receptors, Gut Microbiota, and Immune Response. Front. Immunol. 2016, 7, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheenstra, M.R.; van Harten, R.M.; Veldhuizen, E.J.A.; Haagsman, H.P.; Coorens, M. Cathelicidins Modulate TLR-Activation and Inflammation. Front. Immunol. 2020, 11, 1137. [Google Scholar] [CrossRef] [PubMed]
- Coorens, M.; Schneider, V.A.F.; de Groot, A.M.; van Dijk, A.; Meijerink, M.; Wells, J.M.; Scheenstra, M.R.; Veldhuizen, E.J.A.; Haagsman, H.P. Cathelicidins Inhibit Escherichia coli-Induced TLR2 and TLR4 Activation in a Viability-Dependent Manner. J. Immunol. 2017, 199, 1418–1428. [Google Scholar] [CrossRef] [Green Version]
- Lecorguille, M.; Charles, M.A.; Lepeule, J.; Lioret, S.; de Lauzon-Guillain, B.; Forhan, A.; Tost, J.; Suderman, M.; Heude, B. Association between dietary patterns reflecting one-carbon metabolism nutrients intake before pregnancy and placental DNA methylation. Epigenetics 2022, 17, 715–730. [Google Scholar] [CrossRef]
- Antoun, E.; Issarapu, P.; di Gravio, C.; Shrestha, S.; Betts, M.; Saffari, A.; Sahariah, S.A.; Sankareswaran, A.; Arumalla, M.; Prentice, A.M.; et al. DNA methylation signatures associated with cardiometabolic risk factors in children from India and The Gambia: Results from the EMPHASIS study. Clin. Epigenet. 2022, 14, 6. [Google Scholar] [CrossRef]
- Feigerlova, E.; Demarquet, L.; Melhem, H.; Ghemrawi, R.; Battaglia-Hsu, S.F.; Ewu, E.; Alberto, J.M.; Helle, D.; Weryha, G.; Gueant, J.L. Methyl donor deficiency impairs bone development via peroxisome proliferator-activated receptor-gamma coactivator-1alpha-dependent vitamin D receptor pathway. FASEB J. 2016, 30, 3598–3612. [Google Scholar] [CrossRef] [Green Version]
- Vaishnava, S.; Yamamoto, M.; Severson, K.M.; Ruhn, K.A.; Yu, X.; Koren, O.; Ley, R.; Wakeland, E.K.; Hooper, L.V. The antibacterial lectin RegIIIgamma promotes the spatial segregation of microbiota and host in the intestine. Science 2011, 334, 255–258. [Google Scholar] [CrossRef] [Green Version]
- Antoun, E.; Kitaba, N.T.; Titcombe, P.; Dalrymple, K.V.; Garratt, E.S.; Barton, S.J.; Murray, R.; Seed, P.T.; Holbrook, J.D.; Kobor, M.S.; et al. Maternal dysglycaemia, changes in the infant’s epigenome modified with a diet and physical activity intervention in pregnancy: Secondary analysis of a randomised control trial. PLoS Med. 2020, 17, e1003229. [Google Scholar] [CrossRef]
- Zhang, Q.; Xiao, X.; Zheng, J.; Li, M.; Yu, M.; Ping, F.; Wang, T.; Wang, X. A Maternal High-Fat Diet Induces DNA Methylation Changes That Contribute to Glucose Intolerance in Offspring. Front. Endocrinol. 2019, 10, 871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Li, J.; Liu, S.; Zhang, L.; Xiao, H.; Li, J.; Chen, H.; Petersen, R.B.; Huang, K.; Zheng, L. DNA hypomethylation of inflammation-associated genes in adipose tissue of female mice after multigenerational high fat diet feeding. Int. J. Obes. 2014, 38, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Keleher, M.R.; Zaidi, R.; Shah, S.; Oakley, M.E.; Pavlatos, C.; El Idrissi, S.; Xing, X.; Li, D.; Wang, T.; Cheverud, J.M. Maternal high-fat diet associated with altered gene expression, DNA methylation, and obesity risk in mouse offspring. PLoS ONE 2018, 13, e0192606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elizabeth, K.E.; Praveen, S.L.; Preethi, N.R.; Jissa, V.T.; Pillai, M.R. Folate, vitamin B12, homocysteine and polymorphisms in folate metabolizing genes in children with congenital heart disease and their mothers. Eur. J. Clin. Nutr. 2017, 71, 1437–1441. [Google Scholar] [CrossRef] [PubMed]
- Tripodi, F.; Badone, B.; Vescovi, M.; Milanesi, R.; Nonnis, S.; Maffioli, E.; Bonanomi, M.; Gaglio, D.; Tedeschi, G.; Coccetti, P. Methionine Supplementation Affects Metabolism and Reduces Tumor Aggressiveness in Liver Cancer Cells. Cells 2020, 9, 2491. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.; Harvey, N.C.; Bishop, N.J.; Kennedy, S.; Papageorghiou, A.T.; Schoenmakers, I.; Fraser, R.; Gandhi, S.V.; Carr, A.; D’Angelo, S.; et al. Maternal gestational vitamin D supplementation and offspring bone health (MAVIDOS): A multicentre, double-blind, randomised placebo-controlled trial. Lancet Diabetes Endocrinol. 2016, 4, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Wolsk, H.M.; Harshfield, B.J.; Laranjo, N.; Carey, V.J.; O’Connor, G.; Sandel, M.; Strunk, R.C.; Bacharier, L.B.; Zeiger, R.S.; Schatz, M.; et al. Vitamin D supplementation in pregnancy, prenatal 25(OH)D levels, race, and subsequent asthma or recurrent wheeze in offspring: Secondary analyses from the Vitamin D Antenatal Asthma Reduction Trial. J. Allergy Clin. Immunol. 2017, 140, 1423–1429.e5. [Google Scholar] [CrossRef] [Green Version]
- Purcell, R.H.; Sun, B.; Pass, L.L.; Power, M.L.; Moran, T.H.; Tamashiro, K.L. Maternal stress and high-fat diet effect on maternal behavior, milk composition, and pup ingestive behavior. Physiol. Behav. 2011, 104, 474–479. [Google Scholar] [CrossRef] [Green Version]
- Belenchia, A.M.; Jones, K.L.; Will, M.; Beversdorf, D.Q.; Vieira-Potter, V.; Rosenfeld, C.S.; Peterson, C.A. Maternal vitamin D deficiency during pregnancy affects expression of adipogenic-regulating genes peroxisome proliferator-activated receptor gamma (PPARgamma) and vitamin D receptor (VDR) in lean male mice offspring. Eur. J. Nutr. 2018, 57, 723–730. [Google Scholar] [CrossRef]
- Gazquez, A.; Sanchez-Campillo, M.; Barranco, A.; Rueda, R.; Chan, J.P.; Kuchan, M.J.; Larque, E. Calcifediol During Pregnancy Improves Maternal and Fetal Availability of Vitamin D Compared to Vitamin D3 in Rats and Modifies Fetal Metabolism. Front. Nutr. 2022, 9, 871632. [Google Scholar] [CrossRef]
- Marino, R.; Misra, M. Extra-Skeletal Effects of Vitamin D. Nutrients 2019, 11, 1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabico, S.; Enani, M.A.; Sheshah, E.; Aljohani, N.J.; Aldisi, D.A.; Alotaibi, N.H.; Alshingetti, N.; Alomar, S.Y.; Alnaami, A.M.; Amer, O.E.; et al. Effects of a 2-Week 5000 IU versus 1000 IU Vitamin D3 Supplementation on Recovery of Symptoms in Patients with Mild to Moderate COVID-19: A Randomized Clinical Trial. Nutrients 2021, 13, 2170. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.S.; Fogelman, D.R.; Raghav, K.P.; Heymach, J.V.; Tran, H.T.; Jiang, Z.Q.; Kopetz, S.; Daniel, C.R. Joint prognostic effect of obesity and chronic systemic inflammation in patients with metastatic colorectal cancer. Cancer 2015, 121, 2968–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, T.; Andersson, L.; Hussain, A.; Bosaeus, M.; Jansson, N.; Osmancevic, A.; Hulthen, L.; Holmang, A.; Larsson, I. Lower vitamin D status in obese compared with normal-weight women despite higher vitamin D intake in early pregnancy. Clin. Nutr. 2015, 34, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Rock, C.L.; Emond, J.A.; Flatt, S.W.; Heath, D.D.; Karanja, N.; Pakiz, B.; Sherwood, N.E.; Thomson, C.A. Weight loss is associated with increased serum 25-hydroxyvitamin D in overweight or obese women. Obesity 2012, 20, 2296–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perna, S. Is Vitamin D Supplementation Useful for Weight Loss Programs? A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Medicina 2019, 55, 368. [Google Scholar] [CrossRef] [Green Version]
- Lowry, M.B.; Guo, C.; Zhang, Y.; Fantacone, M.L.; Logan, I.E.; Campbell, Y.; Zhang, W.; Le, M.; Indra, A.K.; Ganguli-Indra, G.; et al. A mouse model for vitamin D-induced human cathelicidin antimicrobial peptide gene expression. J. Steroid Biochem. Mol. Biol. 2020, 198, 105552. [Google Scholar] [CrossRef]
- Villa, C.R.; Taibi, A.; Chen, J.; Ward, W.E.; Comelli, E.M. Colonic Bacteroides are positively associated with trabecular bone structure and programmed by maternal vitamin D in male but not female offspring in an obesogenic environment. Int. J. Obes. 2018, 42, 696–703. [Google Scholar] [CrossRef]
- Schaffler, H.; Herlemann, D.P.; Klinitzke, P.; Berlin, P.; Kreikemeyer, B.; Jaster, R.; Lamprecht, G. Vitamin D administration leads to a shift of the intestinal bacterial composition in Crohn’s disease patients, but not in healthy controls. J. Dig. Dis. 2018, 19, 225–234. [Google Scholar] [CrossRef]
- Naderpoor, N.; Mousa, A.; Fernanda Gomez Arango, L.; Barrett, H.L.; Dekker Nitert, M.; de Courten, B. Effect of Vitamin D Supplementation on Faecal Microbiota: A Randomised Clinical Trial. Nutrients 2019, 11, 2888. [Google Scholar] [CrossRef]
- Chu, D.M.; Antony, K.M.; Ma, J.; Prince, A.L.; Showalter, L.; Moller, M.; Aagaard, K.M. The early infant gut microbiome varies in association with a maternal high-fat diet. Genome Med. 2016, 8, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salyers, A.A.; West, S.E.; Vercellotti, J.R.; Wilkins, T.D. Fermentation of mucins and plant polysaccharides by anaerobic bacteria from the human colon. Appl. Environ. Microbiol. 1977, 34, 529–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaible, T.D.; Harris, R.A.; Dowd, S.E.; Smith, C.W.; Kellermayer, R. Maternal methyl-donor supplementation induces prolonged murine offspring colitis susceptibility in association with mucosal epigenetic and microbiomic changes. Hum. Mol. Genet. 2011, 20, 1687–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manson, J.E.; Cook, N.R.; Lee, I.M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Gordon, D.; Copeland, T.; D’Agostino, D.; et al. Vitamin D Supplements and Prevention of Cancer and Cardiovascular Disease. N. Engl. J. Med. 2019, 380, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Wacker, M.; Holick, M.F. Vitamin D—Effects on skeletal and extraskeletal health and the need for supplementation. Nutrients 2013, 5, 111–148. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weanling Pups (3-Weeks-Old) | ||
|---|---|---|
| mRNA species | Pearson’s correlation coefficient (r) with colonic VDR | p-value |
| Il1β | 0.265 | 0.07 |
| Il6 | 0.405 | <0.01 |
| Tlr4 | 0.273 | 0.06 |
| Adult Pups (15-Weeks-Old) | ||
| mRNA species | Pearson’s correlation coefficient (r) with colonic VDR | p-value |
| Il1β | 0.323 | <0.01 |
| Il6 | 0.294 | <0.01 |
| Tlr4 | 0.419 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teoh, C.M.; Cooper, A.; Renteria, K.M.; Lane, M.; Zhu, J.; Koh, G.Y. Supplementation of Methyl-Donor Nutrients to a High-Fat, High-Sucrose Diet during Pregnancy and Lactation Normalizes Circulating 25-Dihydroxycholecalciferol Levels and Alleviates Inflammation in Offspring. Metabolites 2022, 12, 1252. https://doi.org/10.3390/metabo12121252
Teoh CM, Cooper A, Renteria KM, Lane M, Zhu J, Koh GY. Supplementation of Methyl-Donor Nutrients to a High-Fat, High-Sucrose Diet during Pregnancy and Lactation Normalizes Circulating 25-Dihydroxycholecalciferol Levels and Alleviates Inflammation in Offspring. Metabolites. 2022; 12(12):1252. https://doi.org/10.3390/metabo12121252
Chicago/Turabian StyleTeoh, Chin May, Analynn Cooper, Karisa M. Renteria, Michelle Lane, Jie Zhu, and Gar Yee Koh. 2022. "Supplementation of Methyl-Donor Nutrients to a High-Fat, High-Sucrose Diet during Pregnancy and Lactation Normalizes Circulating 25-Dihydroxycholecalciferol Levels and Alleviates Inflammation in Offspring" Metabolites 12, no. 12: 1252. https://doi.org/10.3390/metabo12121252