
Role of Oxylipins in the Inflammatory-Related Diseases NAFLD, Obesity, and Type 2 Diabetes

Abstract
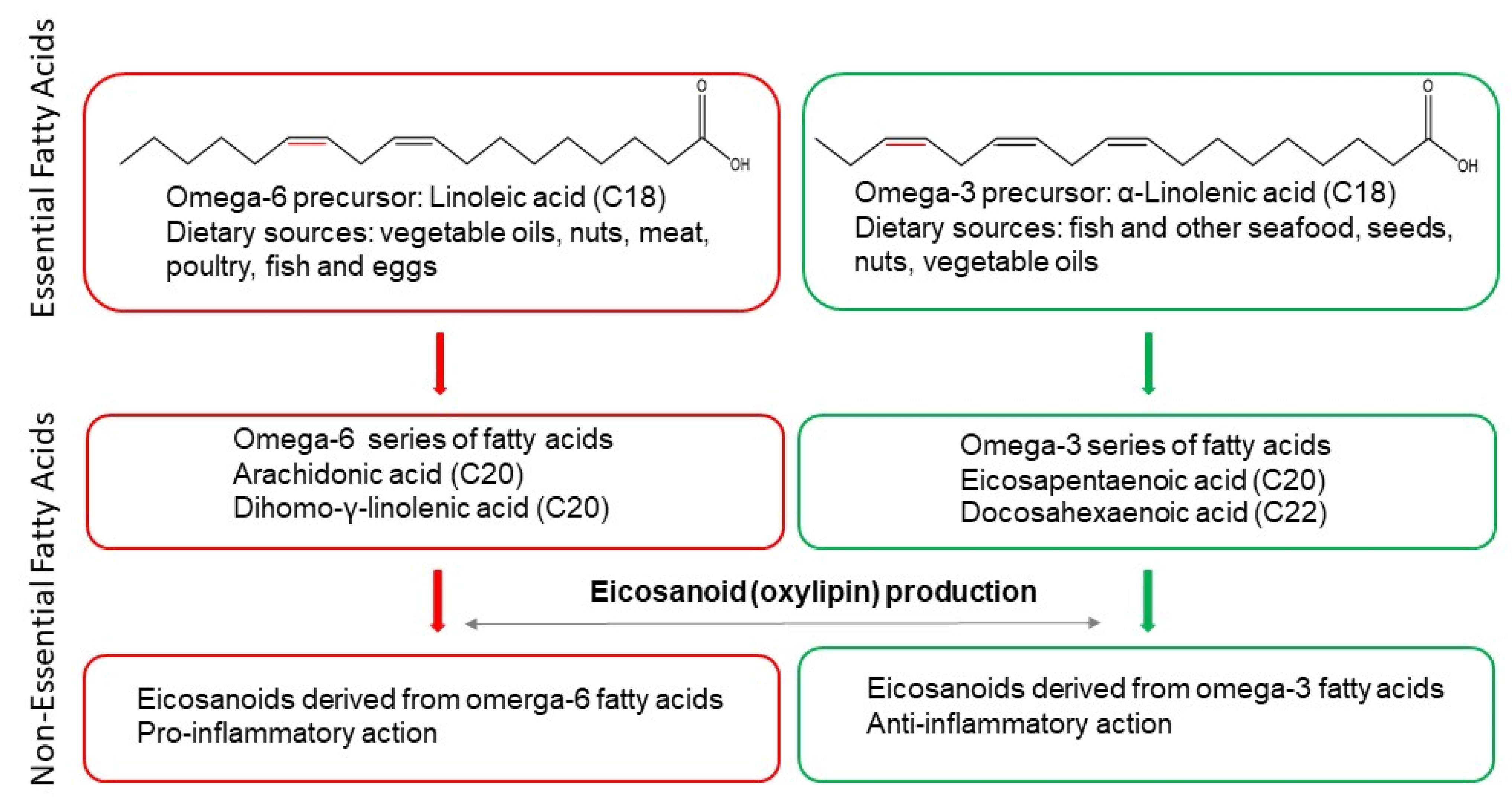
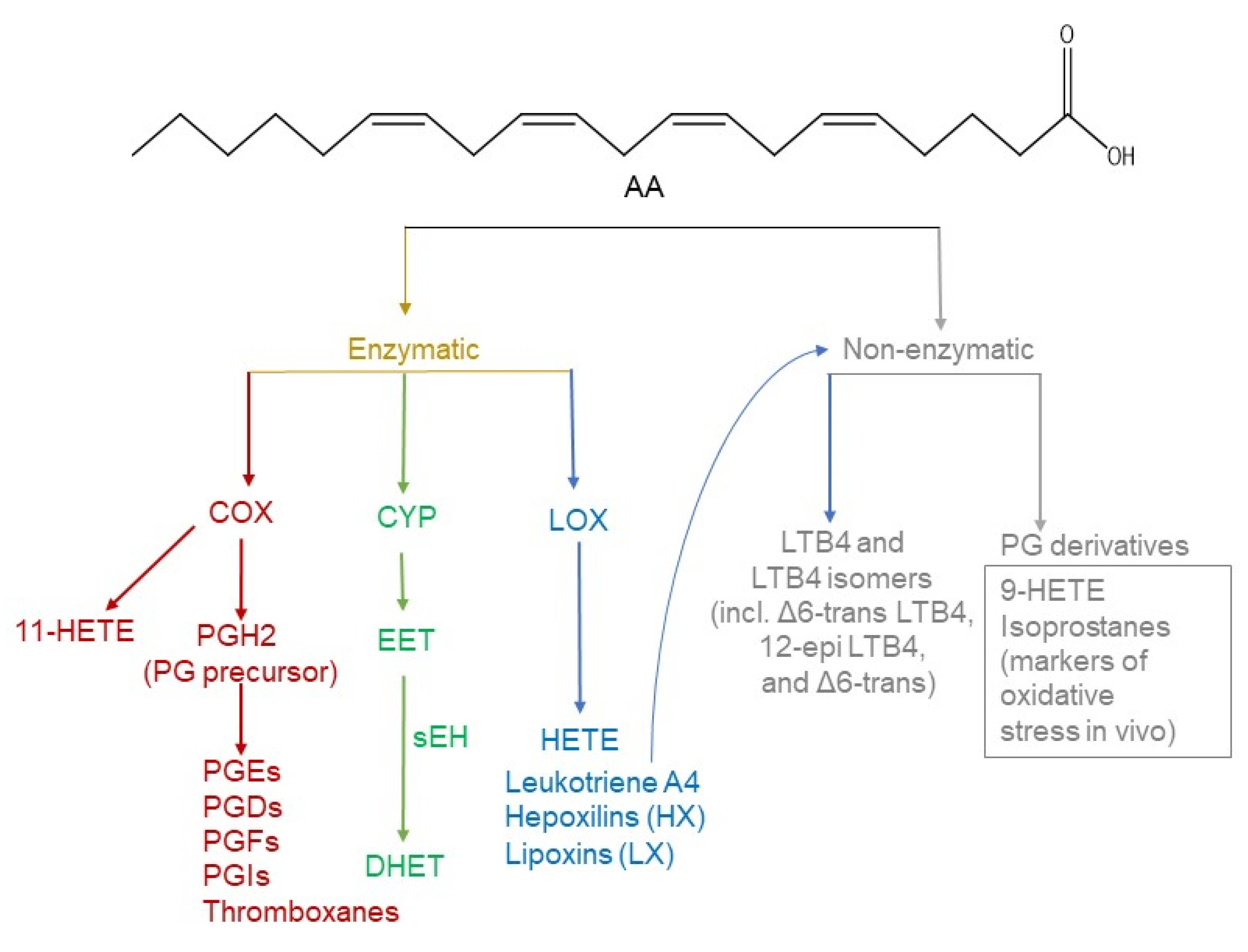
:1. Introduction
2. Non-Alcoholic Fatty Liver Disease (NAFLD)
3. Obesity and Diabetes
4. Technological Advancements and Future Directions
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PUFA | Polyunsaturated fatty acid | COX | Cyclooxygenase |
| LOX | Lipoxygenase | CYP | Cytochrome P450 |
| LA | Linoleic acid | ALA | Alpha-linolenic acid |
| AA | Arachidonic acid | DGLA | Dihomo-gamma-linolenic acid |
| EPA | Eicosapentaenoic acid | DHA | Docosahexaenoic acid |
| PG | Prostaglandin | TX | Thromboxane |
| LT | Leukotriene | NAFLD | Non-alcoholic fatty liver disease |
| NAFL | Non-alcoholic fatty liver | NASH | Non-alcoholic steatohepatosis |
| 5-oxo-ETE | 5-Oxo-eicosatetraenoic acid | HETE | Hydroxyeicosatetraenoic acid |
| HEPE | Hydroxyeicosapentaenoic acid | HDHA | Hydroxydocosahexaenoic acid |
| EET | Epoxyeicosatrienoic acid | DHET | Dihydroxyeicosatrienoic acid |
| LTB4 | Leukotriene B4 | LXA4 | LipoxinA4 |
| HODE | Hydroxyoctadecadienoic acid | PGE2 | Prostaglandin E2 |
| PGE1 | Prostaglandin E1 | PGD1 | Prostaglandin D1 |
| TXB2 | Thromboxane B2 | DiHDPA | Dihydroxydocosapentaenoic acid |
| SSG | Sagittaria sagittifolia | T2D | Type 2 diabetes |
| PGF2α | Prostaglandin F2α | DiHODE | Dihydroxyoctadecadienoic acid |
| DiHOME | Dihydroxyoctadecenoic acid | MS | Mass spectrometry |
| MRM | Multiple reaction monitoring | LC-MS | Liquid chromatography-mass spectrometry |
| IMS-MS | IM-MS | IMS | Ion mobility-mass spectrometry |
| SFC | Supercritical fluid | DESI-MSI | Desorption electrospray ionisation mass spectrometry imaging |
References
- Funk, C.D. Prostaglandins and leukotrienes: Advances in eicosanoid biology. Science 2001, 294, 1871–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, D.L.; Botting, R.M.; Hla, T. Cyclooxygenase isozymes: The biology of prostaglandin synthesis and inhibition. Pharm. Rev 2004, 56, 387–437. [Google Scholar] [CrossRef] [Green Version]
- Smith, W.L.; DeWitt, D.L.; Garavito, R.M. Cyclooxygenases: Structural, cellular, and molecular biology. Annu. Rev. Biochem. 2000, 69, 145–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capdevila, J.H.; Falck, J.R.; Imig, J.D. Roles of the cytochrome P450 arachidonic acid monooxygenases in the control of systemic blood pressure and experimental hypertension. Kidney Int. 2007, 72, 683–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, I. DiscrEET regulators of homeostasis: Epoxyeicosatrienoic acids, cytochrome P450 epoxygenases and vascular inflammation. Trends Pharm. Sci 2007, 28, 448–452. [Google Scholar] [CrossRef]
- Imig, J.D. Targeting epoxides for organ damage in hypertension. J. Cardiovasc. Pharm. 2010, 56, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman, R.J. P-450 metabolites of arachidonic acid in the control of cardiovascular function. Physiol. Rev. 2002, 82, 131–185. [Google Scholar] [CrossRef] [Green Version]
- Spector, A.A.; Norris, A.W. Action of epoxyeicosatrienoic acids on cellular function. Am. J. Physiol. Cell Physiol. 2007, 292, C996–C1012. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.M.; Murphy, S.; Burke, M.; Roman, R.J. 20-hydroxyeicosatetraeonic acid: A new target for the treatment of hypertension. J. Cardiovasc. Pharm. 2010, 56, 336–344. [Google Scholar] [CrossRef]
- Balić, A.; Vlašić, D.; Žužul, K.; Marinović, B.; Bukvić Mokos, Z. Omega-3 Versus Omega-6 Polyunsaturated Fatty Acids in the Prevention and Treatment of Inflammatory Skin Diseases. Int. J. Mol. Sci. 2020, 21, 741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Muhlhausler, B.S.; Cook-Johnson, R.; James, M.; Miljkovic, D.; Duthoit, E.; Gibson, R. Opposing effects of omega-3 and omega-6 long chain polyunsaturated Fatty acids on the expression of lipogenic genes in omental and retroperitoneal adipose depots in the rat. J. Nutr. Metab. 2010, 2010, 927836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, A.; Botta, E.; Holinstat, M. Eicosanoids in inflammation in the blood and the vessel. Front. Pharm. 2022, 13, 997403. [Google Scholar] [CrossRef] [PubMed]
- Sheppe, A.E.F.; Edelmann, M.J. Roles of Eicosanoids in Regulating Inflammation and Neutrophil Migration as an Innate Host Response to Bacterial Infections. Infect. Immun. 2021, 89, e0009521. [Google Scholar] [CrossRef]
- Mendoza, S.R.; Zamith-Miranda, D.; Takacs, T.; Gacser, A.; Nosanchuk, J.D.; Guimaraes, A.J. Complex and Controversial Roles of Eicosanoids in Fungal Pathogenesis. J. Fungi 2021, 7, 254. [Google Scholar] [CrossRef]
- Artru, F.; McPhail, M.J.W.; Triantafyllou, E.; Trovato, F.M. Lipids in Liver Failure Syndromes: A Focus on Eicosanoids, Specialized Pro-Resolving Lipid Mediators and Lysophospholipids. Front. Immunol. 2022, 13, 867261. [Google Scholar] [CrossRef]
- Buczynski, M.W.; Dumlao, D.S.; Dennis, E.A. Thematic Review Series: Proteomics. An integrated omics analysis of eicosanoid biology. J. Lipid Res. 2009, 50, 1015–1038. [Google Scholar] [CrossRef] [Green Version]
- Kurzrok, R.; Lieb, C.C. Biochemical Studies of Human Semen. II. The Action of Semen on the Human Uterus. Proc. Soc. Exp. Biol. Med. 1930, 28, 268–272. [Google Scholar] [CrossRef]
- Fitzpatrick, F.A.; Ennis, M.D.; Baze, M.E.; Wynalda, M.A.; McGee, J.E.; Liggett, W.F. Inhibition of cyclooxygenase activity and platelet aggregation by epoxyeicosatrienoic acids. Influence of stereochemistry. J. Biol. Chem. 1986, 261, 15334–15338. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R. Inhibition of Prostaglandin Synthesis as a Mechanism of Action for Aspirin-Like Drugs. Nat.-New Biol. 1971, 231, 232. [Google Scholar] [CrossRef] [PubMed]
- Breyer, R.M.; Bagdassarian, C.K.; Myers, S.A.; Breyer, M.D. Prostanoid receptors: Subtypes and signaling. Annu. Rev. Pharm. Toxicol 2001, 41, 661–690. [Google Scholar] [CrossRef] [PubMed]
- Capra, V.; Thompson, M.D.; Sala, A.; Cole, D.E.; Folco, G.; Rovati, G.E. Cysteinyl-leukotrienes and their receptors in asthma and other inflammatory diseases: Critical update and emerging trends. Med. Res. Rev. 2007, 27, 469–527. [Google Scholar] [CrossRef]
- Cheng, H.F.; Harris, R.C. Cyclooxygenases, the kidney, and hypertension. Hypertension 2004, 43, 525–530. [Google Scholar] [CrossRef]
- Hao, C.M.; Breyer, M.D. Physiological regulation of prostaglandins in the kidney. Annu. Rev. Physiol. 2008, 70, 357–377. [Google Scholar] [CrossRef]
- Ribeiro, J.D.; Toro, A.A.; Baracat, E.C. Antileukotrienes in the treatment of asthma and allergic rhinitis. J. Pediatr. 2006, 82, S213–S221. [Google Scholar] [CrossRef] [Green Version]
- Sharma, J.N.; Mohammed, L.A. The role of leukotrienes in the pathophysiology of inflammatory disorders: Is there a case for revisiting leukotrienes as therapeutic targets? Inflammopharmacology 2006, 14, 10–16. [Google Scholar] [CrossRef]
- Bedossa, P. Pathology of non-alcoholic fatty liver disease. Liver Int. 2017, 37, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Teli, M.R.; James, O.F.; Burt, A.D.; Bennett, M.K.; Day, C.P. The natural history of nonalcoholic fatty liver: A follow-up study. Hepatology 1995, 22, 1714–1719. [Google Scholar] [CrossRef]
- Zoller, H.; Tilg, H. Nonalcoholic fatty liver disease and hepatocellular carcinoma. Metabolism 2016, 65, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- Cotter, T.G.; Rinella, M. Nonalcoholic Fatty Liver Disease 2020: The State of the Disease. Gastroenterology 2020, 158, 1851–1864. [Google Scholar] [CrossRef] [PubMed]
- Golabi, P.; Paik, J.; Hwang, J.P.; Wang, S.; Lee, H.M.; Younossi, Z.M. Prevalence and outcomes of non-alcoholic fatty liver disease (NAFLD) among Asian American adults in the United States. Liver Int. 2019, 39, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef]
- Svegliati-Baroni, G.; Pierantonelli, I.; Torquato, P.; Marinelli, R.; Ferreri, C.; Chatgilialoglu, C.; Bartolini, D.; Galli, F. Lipidomic biomarkers and mechanisms of lipotoxicity in non-alcoholic fatty liver disease. Free Radic. Biol. Med. 2019, 144, 293–309. [Google Scholar] [CrossRef]
- Banaszczak, M.; Maciejewska, D.; Drozd, A.; Ryterska, K.; Milc, D.J.; Raszeja-Wyszomirska, J.; Wunsch, E.; Gonzalez-Muniesa, P.; Stachowska, E. 5-Lipooxygenase Derivatives as Serum Biomarkers of a Successful Dietary Intervention in Patients with NonAlcoholic Fatty Liver Disease. Medicina 2020, 56, 58. [Google Scholar] [CrossRef] [Green Version]
- Kalveram, L.; Schunck, W.H.; Rothe, M.; Rudolph, B.; Loddenkemper, C.; Holzhutter, H.G.; Henning, S.; Bufler, P.; Schulz, M.; Meierhofer, D.; et al. Regulation of the cytochrome P450 epoxyeicosanoid pathway is associated with distinct histologic features in pediatric non-alcoholic fatty liver disease. Prostaglandins Leukot. Essent. Fat. Acids 2021, 164, 102229. [Google Scholar] [CrossRef]
- Arvind, A.; Osganian, S.A.; Sjoquist, J.A.; Corey, K.E.; Simon, T.G. Epoxygenase-Derived Epoxyeicosatrienoic Acid Mediators Are Associated With Nonalcoholic Fatty Liver Disease, Nonalcoholic Steatohepatitis, and Fibrosis. Gastroenterology 2020, 159, 2232–2234.e2234. [Google Scholar] [CrossRef]
- Marchix, J.; Catheline, D.; Duby, C.; Monthean-Boulier, N.; Boissel, F.; Pedrono, F.; Boudry, G.; Legrand, P. Interactive effects of maternal and weaning high linoleic acid intake on hepatic lipid metabolism, oxylipins profile and hepatic steatosis in offspring. J. Nutr. Biochem. 2020, 75, 108241. [Google Scholar] [CrossRef]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The Diagnosis and Management of Non-alcoholic Fatty Liver Disease: Practice Guideline by the American Gastroenterological Association, American Association for the Study of Liver Diseases, and American College of Gastroenterology. Gastroenterology 2012, 142, 1592–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyson, J.K.; Anstee, Q.M.; McPherson, S. Non-alcoholic fatty liver disease: A practical approach to diagnosis and staging. Frontline Gastroenterol. 2014, 5, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Mauro, S.; Scamporrino, A.; Filippello, A.; Di Pino, A.; Scicali, R.; Malaguarnera, R.; Purrello, F.; Piro, S. Clinical and Molecular Biomarkers for Diagnosis and Staging of NAFLD. Int. J. Mol. Sci. 2021, 22, 11905. [Google Scholar] [CrossRef] [PubMed]
- Maciejewska, D.; Drozd, A.; Skonieczna-Zydecka, K.; Skorka-Majewicz, M.; Dec, K.; Jakubczyk, K.; Pilutin, A.; Stachowska, E. Eicosanoids in Nonalcoholic Fatty Liver Disease (NAFLD) Progression. Do Serum Eicosanoids Profile Correspond with Liver Eicosanoids Content during NAFLD Development and Progression? Molecules 2020, 25, 2026. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Munoz, D.; Kiezel-Tsugunova, M.; Kiss, O.; Uddin, M.; Sunden, M.; Ryaboshapkina, M.; Lind, L.; Oscarsson, J.; Nicolaou, A. Omega-3 carboxylic acids and fenofibrate differentially alter plasma lipid mediators in patients with non-alcoholic fatty liver disease. FASEB J. 2021, 35, e21976. [Google Scholar] [CrossRef]
- Deng, X.; Ke, X.; Tang, Y.; Luo, W.; Dong, R.; Ge, D.; Han, L.; Yang, Y.; Liu, H.; Reyila, T.; et al. Sagittaria sagittifolia polysaccharide interferes with arachidonic acid metabolism in non-alcoholic fatty liver disease mice via Nrf2/HO-1 signaling pathway. Biomed. Pharm. 2020, 132, 110806. [Google Scholar] [CrossRef]
- Yoshino, K.; Hosooka, T.; Shinohara, M.; Aoki, C.; Hosokawa, Y.; Imamori, M.; Ogawa, W. Canagliflozin ameliorates hepatic fat deposition in obese diabetic mice: Role of prostaglandin E-2. Biochem. Biophys. Res. Commun. 2021, 557, 62–68. [Google Scholar] [CrossRef]
- Tuomisto, K.; Palmu, J.; Long, T.; Watrous, J.D.; Mercader, K.; Lagerborg, K.A.; Andres, A.; Salmi, M.; Jalkanen, S.; Vasan, R.S.; et al. A plasma metabolite score of three eicosanoids predicts incident type 2 diabetes: A prospective study in three independent cohorts. BMJ Open Diabetes Res. Care 2022, 10, e002519. [Google Scholar] [CrossRef]
- Miao, Z.J.; Tang, X.; Schultzberg, M.; Zhao, Y.W.; Wang, X.Z. Plasma Resolvin D2 to Leukotriene B-4 Ratio Is Reduced in Diabetic Patients with Ischemic Stroke and Related to Prognosis. Biomed. Res. Int. 2021, 2021, 6657646. [Google Scholar] [CrossRef]
- Tans, R.; Bande, R.; van Rooij, A.; Molloy, B.J.; Stienstra, R.; Tack, C.J.; Wevers, R.A.; Wessels, H.; Gloerich, J.; van Gool, A.J. Evaluation of cyclooxygenase oxylipins as potential biomarker for obesity-associated adipose tissue inflammation and type 2 diabetes using targeted multiple reaction monitoring mass spectrometry. Prostaglandins Leukot. Essent. Fat. Acids 2020, 160, 102157. [Google Scholar] [CrossRef]
- Pawelzik, S.C.; Avignon, A.; Idborg, H.; Boegner, C.; Stanke-Labesque, F.; Jakobsson, P.J.; Sultan, A.; Back, M. Urinary prostaglandin D2 and E2 metabolites associate with abdominal obesity, glucose metabolism, and triglycerides in obese subjects. Prostaglandins Other Lipid Mediat. 2019, 145, 106361. [Google Scholar] [CrossRef] [PubMed]
- Truchan, N.A.; Fenske, R.J.; Sandhu, H.K.; Weeks, A.M.; Patibandla, C.; Wancewicz, B.; Pabich, S.; Reuter, A.; Harrington, J.M.; Brill, A.L.; et al. Human Islet Expression Levels of Prostaglandin E2 Synthetic Enzymes, But Not Prostaglandin EP3 Receptor, Are Positively Correlated with Markers of beta-Cell Function and Mass in Nondiabetic Obesity. ACS Pharm. Transl. Sci. 2021, 4, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Grapov, D.; Fiehn, O.; Campbell, C.; Chandler, C.J.; Burnett, D.J.; Souza, E.C.; Casazza, G.A.; Keim, N.L.; Hunter, G.R.; Fernandez, J.R.; et al. Impact of a weight loss and fitness intervention on exercise-associated plasma oxylipin patterns in obese, insulin-resistant, sedentary women. Physiol. Rep. 2020, 8, e14547. [Google Scholar] [CrossRef] [PubMed]
- Heemskerk, M.M.; Giera, M.; el Bouazzaoui, F.; Lips, M.A.; Pijl, H.; van Dijk, K.W.; van Harmelen, V. Increased PUFA Content and 5-Lipoxygenase Pathway Expression Are Associated with Subcutaneous Adipose Tissue Inflammation in Obese Women with Type 2 Diabetes. Nutrients 2015, 7, 7676–7690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisk, H.L.; Childs, C.E.; Miles, E.A.; Ayres, R.; Noakes, P.S.; Paras-Chavez, C.; Kuda, O.; Kopecky, J.; Antoun, E.; Lillycrop, K.A.; et al. Modification of subcutaneous white adipose tissue inflammation by omega-3 fatty acids is limited in human obesity-a double blind, randomised clinical trial. EBioMedicine 2022, 77, 103909. [Google Scholar] [CrossRef]
- Sanchez-Fernandez, A.; Zandee, S.; Mastrogiovanni, M.; Charabati, M.; Rubbo, H.; Prat, A.; Lopez-Vales, R. Administration of Maresin-1 ameliorates the physiopathology of experimental autoimmune encephalomyelitis. J. Neuroinflammation 2022, 19, 27. [Google Scholar] [CrossRef]
- Dieckmann, S.; Maurer, S.; Fromme, T.; Colson, C.; Virtanen, K.A.; Amri, E.Z.; Klingenspor, M. Fatty Acid Metabolite Profiling Reveals Oxylipins as Markers of Brown but Not Brite Adipose Tissue. Front. Endocrinol. 2020, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Y.; Huang, S.S.; Wu, S.F.; Qi, J.J.; Li, W.C.; Liu, S.S.; Cong, Y.; Chen, H.W.; Lu, L.W.; Shi, S.T.; et al. Clearance of apoptotic cells by mesenchymal stem cells contributes to immunosuppression via PGE2. Ebiomedicine 2019, 45, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.L. Eicosanoids in the gastrointestinal tract. Br. J. Pharm. 2019, 176, 1000–1008. [Google Scholar] [CrossRef] [Green Version]
- Tsiantas, K.; Konteles, S.J.; Kritsi, E.; Sinanoglou, V.J.; Tsiaka, T.; Zoumpoulakis, P. Effects of Non-Polar Dietary and Endogenous Lipids on Gut Microbiota Alterations: The Role of Lipidomics. Int. J. Mol. Sci. 2022, 23, 4070. [Google Scholar] [CrossRef]
- Avila-Roman, J.; Arreaza-Gil, V.; Cortes-Espinar, A.J.; Soliz-Rueda, J.R.; Mulero, M.; Muguerza, B.; Arola-Arnal, A.; Arola, L.; Torres-Fuentes, C. Impact of gut microbiota on plasma oxylipins profile under healthy and obesogenic conditions. Clin. Nutr. 2021, 40, 1475–1486. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Oh, Y.T.; Wan, D.; Watanabe, R.M.; Hammock, B.D.; Youn, J.H. Postprandial effect to decrease soluble epoxide hydrolase activity: Roles of insulin and gut microbiota. J. Nutr. Biochem. 2017, 49, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Crittenden, S.; Goepp, M.; Pollock, J.; Robb, C.T.; Smyth, D.J.; Zhou, Y.; Andrews, R.; Tyrrell, V.; Gkikas, K.; Adima, A.; et al. Prostaglandin E2 promotes intestinal inflammation via inhibiting microbiota-dependent regulatory T cells. Sci. Adv. 2021, 7, eabd7954. [Google Scholar] [CrossRef] [PubMed]
- Chistyakov, D.V.; Gavrish, G.E.; Goriainov, S.V.; Chistyakov, V.V.; Astakhova, A.A.; Azbukina, N.V.; Sergeeva, M.G. Oxylipin Profiles as Functional Characteristics of Acute Inflammatory Responses in Astrocytes Pre-Treated with IL-4, IL-10, or LPS. Int. J. Mol. Sci. 2020, 21, 780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chistyakov, D.V.; Astakhova, A.A.; Goriainov, S.V.; Sergeeva, M.G. Comparison of PPAR Ligands as Modulators of Resolution of Inflammation, via Their Influence on Cytokines and Oxylipins Release in Astrocytes. Int. J. Mol. Sci. 2020, 21, 9577. [Google Scholar] [CrossRef]
- Na, Y.R.; Jung, D.; Stakenborg, M.; Jang, H.; Gu, G.J.; Jeong, M.R.; Suh, S.Y.; Kim, H.J.; Kwon, Y.H.; Sung, T.S.; et al. Prostaglandin E2 receptor PTGER4-expressing macrophages promote intestinal epithelial barrier regeneration upon inflammation. Gut 2021, 70, 2249–2260. [Google Scholar] [CrossRef]
- Hinz, C.; Liggi, S.; Mocciaro, G.; Jung, S.; Induruwa, I.; Pereira, M.; Bryant, C.E.; Meckelmann, S.W.; O′Donnell, V.B.; Farndale, R.W.; et al. A Comprehensive UHPLC Ion Mobility Quadrupole Time-of-Flight Method for Profiling and Quantification of Eicosanoids, Other Oxylipins, and Fatty Acids. Anal. Chem. 2019, 91, 8025–8035. [Google Scholar] [CrossRef]
- Mabud, M.D.A.; Dekrey, M.J.; Cooks, R.G. Surface-Induced Dissociation of Molecular-Ions. Int. J. Mass Spectrom. 1985, 67, 285–294. [Google Scholar] [CrossRef]
- Syka, J.E.; Coon, J.J.; Schroeder, M.J.; Shabanowitz, J.; Hunt, D.F. Peptide and protein sequence analysis by electron transfer dissociation mass spectrometry. Proc. Natl. Acad. Sci. USA 2004, 101, 9528–9533. [Google Scholar] [CrossRef] [Green Version]
- Zubarev, R.A.; Horn, D.M.; Fridriksson, E.K.; Kelleher, N.L.; Kruger, N.A.; Lewis, M.A.; Carpenter, B.K.; McLafferty, F.W. Electron capture dissociation for structural characterization of multiply charged protein cations. Anal. Chem. 2000, 72, 563–573. [Google Scholar] [CrossRef]
- Chait, B.T. Mass spectrometry in the postgenomic era. Annu. Rev. Biochem. 2011, 80, 239–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kind, T.; Fiehn, O. Advances in structure elucidation of small molecules using mass spectrometry. Bioanal. Rev. 2010, 2, 23–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCullagh JSO, O.N. Mass Spectrometry; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Mann, M.; Hendrickson, R.C.; Pandey, A. Analysis of proteins and proteomes by mass spectrometry. Annu. Rev. Biochem. 2001, 70, 437–473. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Y.; Zhang, Q.B. Comprehensive analysis of oxylipins in human plasma using reversed-phase liquid chromatography-triple quadrupole mass spectrometry with heatmap-assisted selection of transitions. Anal. Bioanal. Chem. 2019, 411, 367–385. [Google Scholar] [CrossRef] [PubMed]
- Theodoridis, G.A.; Gika, H.G.; Want, E.J.; Wilson, I.D. Liquid chromatography-mass spectrometry based global metabolite profiling: A review. Anal. Chim. Acta 2012, 711, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Molnar, I.; Horvath, C. Reverse-phase chromatography of polar biological substances: Separation of catechol compounds by high-performance liquid chromatography. Clin. Chem. 1976, 22, 1497–1502. [Google Scholar] [CrossRef]
- Kanu, A.B.; Dwivedi, P.; Tam, M.; Matz, L.; Hill, H.H., Jr. Ion mobility-mass spectrometry. J. Mass Spectrom. 2008, 43, 1–22. [Google Scholar] [CrossRef]
- Lanucara, F.; Holman, S.W.; Gray, C.J.; Eyers, C.E. The power of ion mobility-mass spectrometry for structural characterization and the study of conformational dynamics. Nat. Chem. 2014, 6, 281–294. [Google Scholar] [CrossRef]
- Hill, H.H., Jr.; Siems, W.F.; St Louis, R.H.; McMinn, D.G. Ion mobility spectrometry. Anal. Chem. 1990, 62, 1201A–1209A. [Google Scholar] [CrossRef]
- Dodds, J.N.; Baker, E.S. Ion Mobility Spectrometry: Fundamental Concepts, Instrumentation, Applications, and the Road Ahead. J. Am. Soc. Mass Spectrom. 2019, 30, 2185–2195. [Google Scholar] [CrossRef]
- Paglia, G.; Smith, A.J.; Astarita, G. Ion mobility mass spectrometry in the omics era: Challenges and opportunities for metabolomics and lipidomics. Mass Spectrom. Rev. 2022, 41, 722–765. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.A.; Leaptrot, K.L.; May, J.C.; McLean, J.A. New Frontiers in Lipidomics Analyses using Structurally Selective Ion Mobility-Mass Spectrometry. Trends Anal. Chem. 2019, 116, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Domalain, V.; Tognetti, V.; Hubert-Roux, M.; Lange, C.M.; Joubert, L.; Baudoux, J.; Rouden, J.; Afonso, C. Role of cationization and multimers formation for diastereomers differentiation by ion mobility-mass spectrometry. J. Am. Soc. Mass Spectrom. 2013, 24, 1437–1445. [Google Scholar] [CrossRef] [PubMed]
- Stauber, J.; MacAleese, L.; Franck, J.; Claude, E.; Snel, M.; Kaletas, B.K.; Wiel, I.M.; Wisztorski, M.; Fournier, I.; Heeren, R.M. On-tissue protein identification and imaging by MALDI-ion mobility mass spectrometry. J. Am. Soc. Mass Spectrom. 2010, 21, 338–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurneczko, E.; Kalapothakis, J.; Campuzano, I.D.; Morris, M.; Barran, P.E. Effects of drift gas on collision cross sections of a protein standard in linear drift tube and traveling wave ion mobility mass spectrometry. Anal. Chem. 2012, 84, 8524–8531. [Google Scholar] [CrossRef] [PubMed]
- Bouza, M.; Li, Y.; Wang, A.C.; Wang, Z.L.; Fernandez, F.M. Triboelectric Nanogenerator Ion Mobility-Mass Spectrometry for In-Depth Lipid Annotation. Anal. Chem. 2021, 93, 5468–5475. [Google Scholar] [CrossRef]
- Fenn, L.S.; Kliman, M.; Mahsut, A.; Zhao, S.R.; McLean, J.A. Characterizing ion mobility-mass spectrometry conformation space for the analysis of complex biological samples. Anal. Bioanal. Chem. 2009, 394, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Fenn, L.S.; McLean, J.A. Biomolecular structural separations by ion mobility-mass spectrometry. Anal. Bioanal. Chem. 2008, 391, 905–909. [Google Scholar] [CrossRef]
- Kliman, M.; May, J.C.; McLean, J.A. Lipid analysis and lipidomics by structurally selective ion mobility-mass spectrometry. Biochim. Biophys. Acta 2011, 1811, 935–945. [Google Scholar] [CrossRef] [Green Version]
- Hellhake, S.; Meckelmann, S.W.; Empl, M.T.; Rentmeister, K.; Wissdorf, W.; Steinberg, P.; Schmitz, O.J.; Benter, T.; Schebb, N.H. Non-targeted and targeted analysis of oxylipins in combination with charge-switch derivatization by ion mobility high-resolution mass spectrometry. Anal. Bioanal. Chem. 2020, 412, 5743–5757. [Google Scholar] [CrossRef]
- Chollet, C.; Boutet-Mercey, S.; Laboureur, L.; Rincon, C.; Mejean, M.; Jouhet, J.; Fenaille, F.; Colsch, B.; Touboul, D. Supercritical fluid chromatography coupled to mass spectrometry for lipidomics. J. Mass Spectrom. 2019, 54, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liang, Y.; Yang, J.; Ye, F.; Zhou, T.; Gongke, L. Advances of supercritical fluid chromatography in lipid profiling. J. Pharm. Anal. 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.T. Supercritical fluid chromatography. Anal. Chem. 2010, 82, 4925–4935. [Google Scholar] [CrossRef] [PubMed]
- Saito, M. History of supercritical fluid chromatography: Instrumental development. J. Biosci. Bioeng. 2013, 115, 590–599. [Google Scholar] [CrossRef]
- Kalikova, K.; Slechtova, T.; Vozka, J.; Tesarova, E. Supercritical fluid chromatography as a tool for enantioselective separation; A review. Anal. Chim. Acta 2014, 821, 1–33. [Google Scholar] [CrossRef]
- Jin, W.; Yang, J.; Liu, D.; Zhong, Q.; Zhou, T. Determination of inflammation-related lipids in depressive rats by on-line supercritical fluid extraction-supercritical fluid chromatography-tandem mass spectrometry. J. Pharm. Biomed. Anal. 2021, 203, 114210. [Google Scholar] [CrossRef]
- Kumari, A.U.S.J.; Acharya, S.R.; Bergquist, J. A novel, fast and sensitive supercritical fluid chromatography-tandem mass spectrometry (SFC-MS/MS) method for analysis of arachidonic acid metabolites. Analyst 2018, 143, 3661–3669. [Google Scholar] [CrossRef] [Green Version]
- Qi, K.; Wu, L.; Liu, C.; Pan, Y. Recent Advances of Ambient Mass Spectrometry Imaging and Its Applications in Lipid and Metabolite Analysis. Metabolites 2021, 11, 780. [Google Scholar] [CrossRef]
- Su, H.; Jiang, Z.H.; Chiou, S.F.; Shiea, J.; Wu, D.C.; Tseng, S.P.; Jain, S.H.; Chang, C.Y.; Lu, P.L. Rapid Characterization of Bacterial Lipids with Ambient Ionization Mass Spectrometry for Species Differentiation. Molecules 2022, 27, 2772. [Google Scholar] [CrossRef]
- Bowman, A.P.; Heeren, R.M.A.; Ellis, S.R. Advances in mass spectrometry imaging enabling observation of localised lipid biochemistry within tissues. TrAC Trends Anal. Chem. 2019, 120, 115197. [Google Scholar] [CrossRef]
- Passarelli, M.K.; Winograd, N. Lipid imaging with time-of-flight secondary ion mass spectrometry (ToF-SIMS). Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 976–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takats, Z.; Wiseman, J.M.; Gologan, B.; Cooks, R.G. Mass spectrometry sampling under ambient conditions with desorption electrospray ionization. Science 2004, 306, 471–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Qi, F.; Pan, Y. Imaging of Polar and Nonpolar Lipids Using Desorption Electrospray Ionization/Post-photoionization Mass Spectrometry. Methods Mol. Biol. 2021, 2306, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Bowman, A.P.; Bogie, J.F.J.; Hendriks, J.J.A.; Haidar, M.; Belov, M.; Heeren, R.M.A.; Ellis, S.R. Evaluation of lipid coverage and high spatial resolution MALDI-imaging capabilities of oversampling combined with laser post-ionisation. Anal. Bioanal. Chem. 2020, 412, 2277–2289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coras, R.; Kavanaugh, A.; Kluzniak, A.; Holt, D.; Weilgosz, A.; Aaron, A.; Quehenberger, O.; Ritchlin, C.; Guma, M. Differences in oxylipin profile in psoriasis versus psoriatic arthritis. Arthritis Res. 2021, 23, 200. [Google Scholar] [CrossRef]
- Bansal, K.; Narayana, Y.; Patil, S.A.; Balaji, K.N.M. bovis BCG induced expression of COX-2 involves nitric oxide-dependent and -independent signaling pathways. J. Leukoc. Biol. 2009, 85, 804–816. [Google Scholar] [CrossRef]
- Eckmann, L.; Stenson, W.F.; Savidge, T.C.; Lowe, D.C.; Barrett, K.E.; Fierer, J.; Smith, J.R.; Kagnoff, M.F. Role of intestinal epithelial cells in the host secretory response to infection by invasive bacteria. Bacterial entry induces epithelial prostaglandin h synthase-2 expression and prostaglandin E2 and F2alpha production. J. Clin. Investig. 1997, 100, 296–309. [Google Scholar] [CrossRef] [Green Version]
- Teng, O.; Ang, C.K.E.; Guan, X.L. Macrophage–Bacteria Interactions—A Lipid-Centric Relationship. Front. Immunol. 2017, 8, 1836. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Oxylipin Abbreviation | Full Name | Fatty Acid Source | Enzyme | Method of Detection | REF |
|---|---|---|---|---|---|
| 5-HETE | 5-Hydroxyeicosatetraenoic acid | AA | LOX | Liquid chromatography-mass spectrometry (LC-MS/MS) Unspecified lipidomics approach | [40] [57] |
| 12-HETE | 12-Hydroxyeicosatetraenoic acid | AA | LOX | LC-MS/MS | [40] |
| 18-HEPE | 18-Hydroxyeicosapentaenoic acid | EPA | unknown, non-enzymatic | UPLC-MS/MS | [45] |
| 17-HDOHE (17-HDHA) | 17-Hydroxydocosahexaenoic acid | DHA | LOX | UPLC-MS/MS | [45] |
| 9-HODE | 9-Hydroxyoctadecadienoic acid | LA | LOX | HPLC | [44] |
| 13-HODE | 13-Hydroxyoctadecadienoic acid | LA | LOX | HPLC | [44] |
| 5-OxoETE | 5-Oxo-eicosatetraenoic acid | AA | LOX | HPLC | [37] |
| 9,10-DiHOME | 9,10-Dihydroxyoctadecenoic acid | LA | CYP | LC-MS/MS | [53] |
| 11,12-DiHETrE (DHET) | 11,12-Dihydroxyeicosatrienoic acid | AA | CYP | LC-MS | [39] |
| 14,15-DiHETrE (DHET) | 14,15-Dihydroxyeicosatrienoic acid | AA | CYP | LC-MS | [39] |
| RvD2 | 7S,16R,17S-Trihydroxydocosahexaenoic acid | DHA | LOX | Enzyme immunoassay (EIA) | [49] |
| LTB4 (Leukotriene B4) | 5S,12R-Dihydroxyeicosatetraenoic acid | AA | LOX | LC-MS/MS Enzyme immunoassay (EIA) LC-MS/MS | [40] [49] [61] |
| LXA4 (Lipoxin A4) | 5S,6R,15S-Trihydroxyeicosatetraenoic acid | AA | LOX | HPLC LC-MS/MS | [37] [40] |
| Maresin1 | 7R,14S-Dihydroxydocosahexaenoic acid | DHA | LOX | LC-MS/MS | [56] |
| 5,6-EET (EpETrE) | 5(6)-Epoxyeicosatrienoic acid | AA | CYP | Unspecified lipidomics approach | [57] |
| 8(9)-EET (EpETrE) | 8(9)-Epoxyeicosatrienoic acid | AA | CYP | LC-MS/MS | [40] |
| 11(12)-EET (EpETrE) | 11(12)-Epoxyeicosatrienoic acid | AA | CYP | LC-MS | [39] |
| 14(15)-EET (EpETrE) | 5(6)-Epoxyeicosatrienoic acid | AA | CYP | LC-MS | [39] |
| PGD1 (prostaglandin D1) | 9α,15S-dihydroxy-11-oxo-prost-13E-en-1-oic acid | DGLA | COX | UPLC-MS/MS | [45] |
| PGE1 (prostaglandin E1) | 9-oxo-11α,15S-dihydroxy-prost-13E-en-1-oic acid | DGLA | COX | UPLC-MS/MS | [45] |
| PGE2 (Prostaglandin E2) | 9-oxo-11α,15S-dihydroxy-prosta-5Z,13E-dien-1-oic acid | AA | COX | UPLC-MS/MS LC-MS/MS UPLC-MS/MS Direct injection MS ELISA LC-MS/MS | [45] [47] [50] [52] [58] [63] |
| 13,14-dihydro-15-keto PGE2 | 9,15-dioxo-11α-hydroxy-prost-5Z-en-1-oic acid | AA | COX | UPLC-MS/MS | [50] |
| PGF2α | 9α,11α,15S-trihydroxy-prosta-5Z,13E-dien-1-oic acid | AA | COX | UPLC-MS/MS | [50] |
| TXB2 (Thromboxane B2) | 9α,11,15S-trihydroxythromba-5Z,13E-dien-1-oic acid | AA | COX | UPLC-MS/MS | [45] |
| 19,20-DiHDPA | 19,20-dihydroxy-4Z,7Z,10Z,13Z,16Z-docosapentaenoic acid | DHA | CYP | UPLC-MS/MS | [45] |
| 8-iso-prostaglandin A1 | 9-oxo-15S-hydroxy-(8β)-prosta-10,13E-dien-1-oic acid | DGLA | non-cyclooxygenase origin (minor impurity during commercial preparations of PGE1) | LC-MS | [48] |
| 12-HHTrE | 12S-hydroxy-5Z,8E,10E-heptadecatrienoic acid | AA | COX | LC-MS | [48] |
| 15-keto-PGE2 | 9,15-dioxo-11α-hydroxy-prosta-5Z,13E-dien-1-oic acid | AA | COX | UPLC-MS/MS | [50] |
| 9,10-DiHODE | 9,10-dihydroxy-12Z,15Z-octadecadienoic acid | LA | CYP | LC-MS/MS | [53] |
| 12,13-DiHODE | 12,13-dihydroxy-9Z,15Z-octadecadienoic acid | LA | CYP | LC-MS/MS | [53] |
| RvE3 | 17R,18S-dihydroxy-5Z,8Z,11Z,13E,15E-eicosapentaenoic acid | EPA | LOX | LC-MS/MS | [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Misheva, M.; Johnson, J.; McCullagh, J. Role of Oxylipins in the Inflammatory-Related Diseases NAFLD, Obesity, and Type 2 Diabetes. Metabolites 2022, 12, 1238. https://doi.org/10.3390/metabo12121238
Misheva M, Johnson J, McCullagh J. Role of Oxylipins in the Inflammatory-Related Diseases NAFLD, Obesity, and Type 2 Diabetes. Metabolites. 2022; 12(12):1238. https://doi.org/10.3390/metabo12121238
Chicago/Turabian StyleMisheva, Mariya, Jethro Johnson, and James McCullagh. 2022. "Role of Oxylipins in the Inflammatory-Related Diseases NAFLD, Obesity, and Type 2 Diabetes" Metabolites 12, no. 12: 1238. https://doi.org/10.3390/metabo12121238