
Effects of Enzymatic Cottonseed Protein Concentrate as a Feed Protein Source on the Growth, Plasma Parameters, Liver Antioxidant Capacity and Immune Status of Largemouth Bass (Micropterus salmoides)

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Growth Performance
2.2. Proximate Composition of Whole Fish
2.3. Plasma Parameters
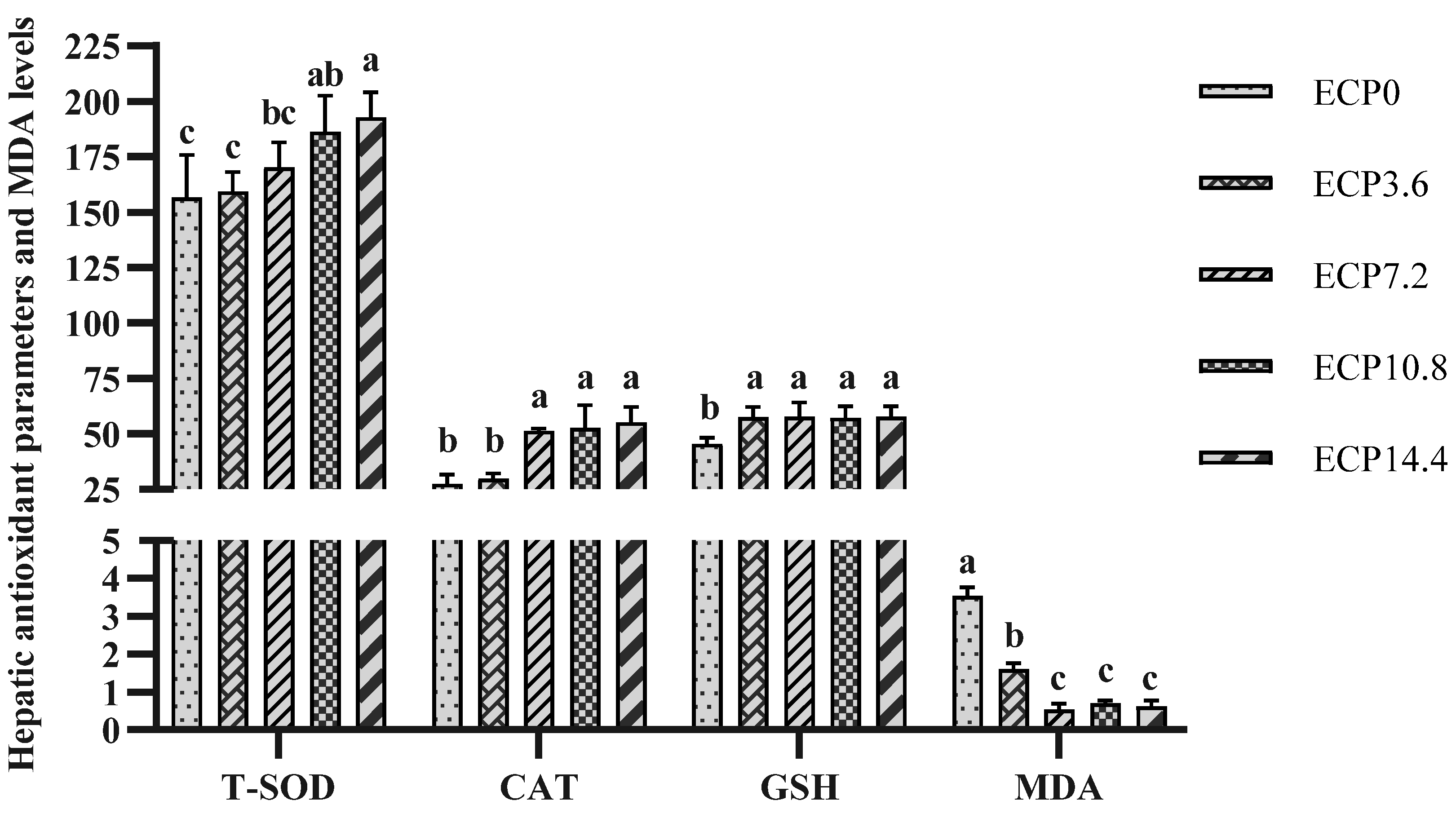
2.4. Hepatic Antioxidant Parameters and MDA Levels
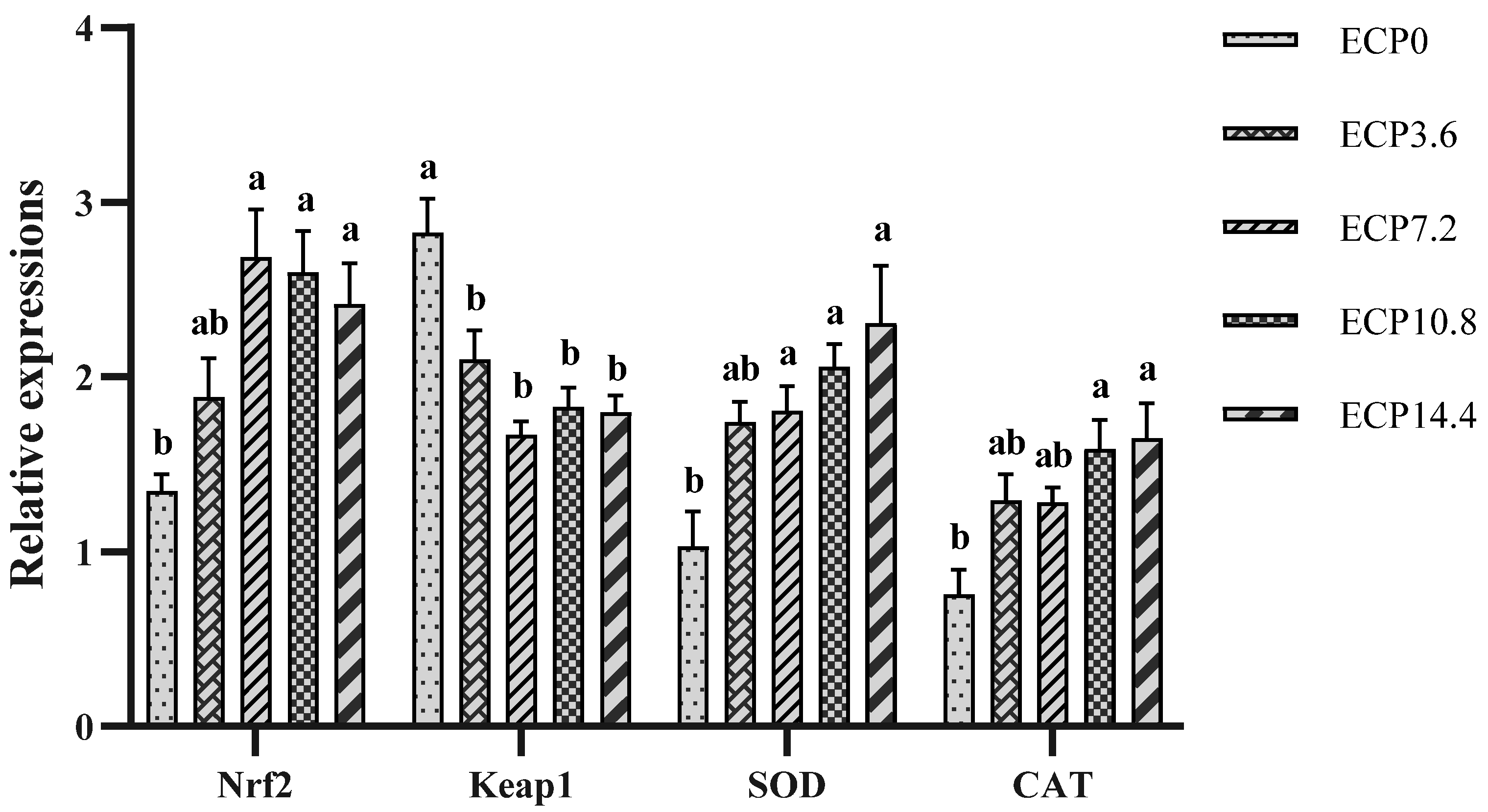
2.5. Expression of Antioxidant Genes mRNA
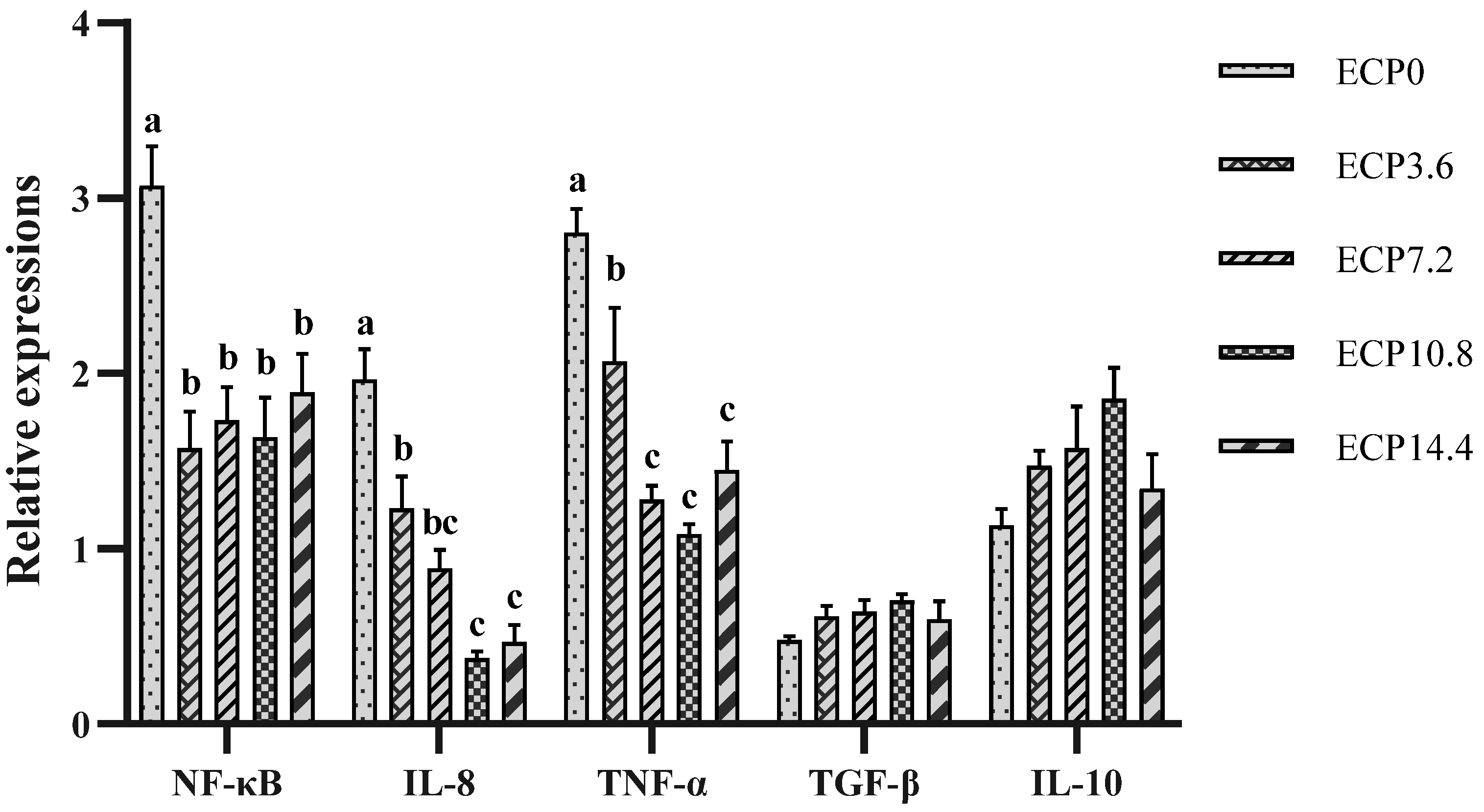
2.6. Expression of Immune-Related Genes mRNA
3. Discussion
4. Materials and Methods
4.1. Experimental Diets
4.2. Feeding Trial
4.3. Sample Collection
4.4. Experimental Parameter Detection
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hussein, G.H.G.; Chen, M.; Qi, P.P.; Cui, Q.K.; Yu, Y.; Hu, W.H.; Tian, Y.; Fan, Q.X.; Gao, Z.X.; Feng, M.W.; et al. Aquaculture industry development, annual price analysis and out-of-season spawning in largemouth bass Micropterus salmoides. Aquaculture 2020, 519, 734901. [Google Scholar] [CrossRef]
- Tidwell, J.H.; Webster, C.D.; Coyle, S.D. Effects of dietary protein level on second year growth and water quality for largemouth bass (Micropterus salmoides) raised in ponds. Aquaculture 1996, 145, 213–223. [Google Scholar] [CrossRef]
- Huang, D.; Wu, Y.B.; Lin, Y.Y. Dietary protein and lipid requirements for juvenile largemouth bass, Micropterus salmoides. J. World Aquac. Soc. 2017, 48, 782–790. [Google Scholar] [CrossRef]
- Cai, Z.N.; Qian, X.Q.; Xie, S.Q. Optimal dietary protein concentrations for largemouth bass (Micropterus salmoides) of different sizes (10–500 g). Aquac. Int. 2020, 28, 831–840. [Google Scholar] [CrossRef]
- Li, X.Y.; Zheng, S.X.; Ma, X.K.; Cheng, K.M.; Wu, G.Y. Effects of dietary protein and lipid levels on the growth performance, feed utilization, and liver histology of largemouth bass (Micropterus salmoides). Amino Acids 2020, 52, 1043–1061. [Google Scholar] [CrossRef]
- Hardy, R.W. Utilization of plant proteins in fish diets: Effects of global demand and supplies of fishmeal. Aquac. Res. 2010, 41, 770–776. [Google Scholar] [CrossRef]
- Olsen, R.L.; Hasan, M.R. A limited supply of fishmeal: Impact on future increases in global aquaculture production. Trends Food Sci. Technol. 2012, 27, 12–128. [Google Scholar] [CrossRef]
- Jannathulla, R.; Rajaram, V.; Kalanjiam, R.; Ambasankar, K.; Muralidhar, M.; Dayal, J.S. Fishmeal availability in the scenarios of climate change: Inevitability of fishmeal replacement in aquafeeds and approaches for the utilization of plant protein sources. Aquac. Res. 2019, 50, 3493–3506. [Google Scholar] [CrossRef]
- Kumar, M.; Potkule, J.; Patil, S.; Saxena, S.; Patil, P.G.; Mageshwaran, V.; Puniad, S.; Varghesee, E.; Mahapatra, A.; Ashtaputrea, N.; et al. Extraction of ultra-low gossypol protein from cottonseed: Characterization based on antioxidant activity, structural morphology and functional group analysis. LWT 2021, 140, 110692. [Google Scholar] [CrossRef]
- Anderson, A.D.; Alam, M.S.; Watanabe, W.O.; Carroll, P.M.; Wedegaertner, T.C.; Dowd, M.K. Full replacement of menhaden fish meal protein by low-gossypol cottonseed flour protein in the diet of juvenile black sea bass Cent. Striata. Aquac. 2016, 464, 618–628. [Google Scholar] [CrossRef]
- Yin, B.; Liu, H.G.; Tan, B.P.; Dong, X.H.; Chi, S.Y.; Yang, Q.H.; Zhang, S.; Chen, L.Q. Cottonseed protein concentrate (CPC) suppresses immune function in different intestinal segments of hybrid grouper ♀Epinephelus fuscoguttatus × ♂Epinephelus lanceolatu via TLR-2/MyD88 signaling pathways. Fish Shellfish Immunol. 2018, 81, 318–328. [Google Scholar] [CrossRef]
- Wan, M.G.; Yin, P.; Fang, W.P.; Xie, S.W.; Chen, S.J.; Tian, L.X.; Niu, J. The effect of replacement of fishmeal by concentrated dephenolization cottonseed protein on the growth, body composition, haemolymph indexes and haematological enzyme activities of the Pacific white shrimp (Litopenaeus vannamei). Aquac. Nutr. 2018, 24, 1845–1854. [Google Scholar] [CrossRef]
- Shen, J.F.; Liu, H.Y.; Tan, B.P.; Dong, X.H.; Yang, Q.H.; Chi, S.Y.; Zhang, S. Effects of replacement of fishmeal with cottonseed protein concentrate on the growth, intestinal microflora, haematological and antioxidant indices of juvenile golden pompano (Trachinotus ovatus). Aquac. Nutr. 2020, 26, 1119–1130. [Google Scholar] [CrossRef]
- Ye, G.L.; Dong, X.H.; Yang, Q.H.; Chi, S.Y.; Liu, H.Y.; Zhang, H.T.; Tan, B.P.; Zhang, S. Low-gossypol cottonseed protein concentrate used as a replacement of fish meal for juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂): Effects on growth performance, immune responses and intestinal microbiota. Aquaculture 2020, 524, 735309. [Google Scholar] [CrossRef]
- Jiang, S.; Chen, Z.B.; Zhou, F.L.; Yang, Q.B.; Huang, J.H.; Yang, L.S.; Li, Y.D.; Jiang, S.G. Study on partial replacement of fish meal with concentrated dephenolized cottonseed protein in feed of Penaeus monodon. Aquac. Res. 2021, 52, 3871–3881. [Google Scholar] [CrossRef]
- He, Y.F.; Guo, X.W.; Tan, B.P.; Dong, X.H.; Yang, Q.H.; Liu, H.; Zhang, S.; Chi, S.Y. Replacing fishmeal with cottonseed protein concentrate in feed for pearl gentian groupers (Epinephelus fuscoguttatus♀ × E. lanceolatus♂): Effects on growth and expressions of key genes involved in appetite and hepatic glucose and lipid metabolism. Aquac. Rep. 2021, 20, 100710. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, Z.L.; Niu, J. Growth performance, intestinal histomorphology, body composition, hematological and antioxidant parameters of Oncorhynchus mykiss were not detrimentally affected by replacement of fish meal with concentrated dephenolization cottonseed protein. Aquac. Rep. 2021, 19, 100557. [Google Scholar] [CrossRef]
- Sun, X.D. Enzymatic hydrolysis of soy proteins and the hydrolysates utilisation. Int. J. Food Sci. Technol. 2011, 46, 2447–2459. [Google Scholar] [CrossRef]
- Ma, X.K.; Shang, Q.H.; Hu, J.X.; Liu, H.S.; Brøkner, C.; Piao, X.S. Effects of replacing soybean meal, soy protein concentrate, fermented soybean meal or fish meal with enzyme-treated soybean meal on growth performance, nutrient digestibility, antioxidant capacity, immunity and intestinal morphology in weaned pigs. Livest. Sci. 2019, 225, 39–46. [Google Scholar] [CrossRef]
- Li, S.L.; Ding, G.T.; Song, F.; Sang, C.Y.; Wang, A.; Chen, N.S. Comparison of dehulled, fermented and enzyme-treated soybean meal in diets for largemouth bass, Micropterus salmoides: Effects on growth performance, feed utilization, immune response and intestinal morphology. Anim. Feed Sci. Technol. 2020, 267, 114548. [Google Scholar] [CrossRef]
- Gui, D.; Liu, W.B.; Shao, X.P.; Xu, W.N. Effects of different dietary levels of cottonseed meal protein hydrolysate on growth, digestibility, body composition and serum biochemical indices in crucian carp (Carassius auratus gibelio). Anim. Feed Sci. Technol. 2010, 156, 112–120. [Google Scholar] [CrossRef]
- Tang, X.P.; Xiang, R.; Chen, S.J.; Yang, S.F.; Liu, H.; Fang, R.J.; Li, A.K. Effects of Fermented Cottonseed Meal and Enzymatic Hydrolyzed Cottonseed Meal on Amino Acid Digestibility and Metabolic Energy in White Leghorn Rooster. Pak. J. Zool. 2018, 50, 957–962. [Google Scholar] [CrossRef]
- Song, W.G.; Kong, X.Z.; Hua, Y.F.; Chen, Y.M.; Zhang, C.M.; Chen, Y.X. Identification of antibacterial peptides generated from enzymatic hydrolysis of cottonseed proteins. LWT 2020, 125, 109199. [Google Scholar] [CrossRef]
- Wang, L.Y.; Ma, M.G.; Yu, Z.P.; Du, S.K. Preparation and identification of antioxidant peptides from cottonseed proteins. Food Chem. 2021, 352, 129399. [Google Scholar] [CrossRef] [PubMed]
- Filho, J.G.D.O.; Rodrigues, J.M.; Valadares, A.C.F.; Almeida, A.B.D.; Valencia-Mejia, E.; Fernandes, K.F.; Lemes, A.C.; Egea, M.B.; Dyszy, F.H. Bioactive properties of protein hydrolysate of cottonseed byproduct: Antioxidant, antimicrobial, and angiotensin-converting enzyme (ACE) inhibitory activities. Waste Biomass Valorization 2021, 12, 1395–1404. [Google Scholar] [CrossRef]
- Wang, K.Z.; Xu, W.N.; Zhou, M.; Zhang, D.D.; Sun, C.X.; Qian, Y.; Liu, W.B. Effects of fishmeal replacement with cottonseed meal protein hydrolysate on growth, digestion and intestinal histology of juvenile Chinese soft-shelled turtle, Pelodiscus sinensis. Aquac. Nutr. 2018, 24, 1406–1415. [Google Scholar] [CrossRef]
- Yuan, X.Y.; Jiang, G.Z.; Cheng, H.H.; Cao, X.F.; Shi, H.J.; Liu, W.B. An evaluation of replacing fish meal with cottonseed meal protein hydrolysate in diet for juvenile blunt snout bream (Megalobrama amblycephala): Growth, antioxidant, innate immunity and disease resistance. Aquac. Nutr. 2019, 25, 1334–1344. [Google Scholar] [CrossRef]
- Song, Z.D.; Li, P.Y.; Wang, J.Y.; Sun, Y.Z.; Wang, C.Q. Dietary inclusion of hydrolyzed soybean and cottonseed meals influence digestion, metabolic enzymes, and growth-related hormones and growth of juvenile turbot (Scophthalmus maximus). Aquac. Int. 2018, 26, 1017–1033. [Google Scholar] [CrossRef]
- Haghbayan, S.; Shamsaie Mehrgan, M. The effect of replacing fish meal in the diet with enzyme-treated soybean meal (HP310) on growth and body composition of rainbow trout fry. Molecules 2015, 20, 21058–21066. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Chi, S.Y.; Li, S.; Cheng, X.L.; Gao, W.H.; Xu, Q.Q.; Zhang, W.B.; Zhou, X.Q. Substitution of fish meal with enzyme-treated soybean in diets for juvenile largemouth bass (Micropterus salmoides). Aquac. Nutr. 2021, 27, 1569–1577. [Google Scholar] [CrossRef]
- Berge, G.E.; Lied, E.; Espe, M. Absorption and incorporation of dietary free and protein bound (U14C)-lysine in Atlantic cod (Gadus morhua). Comp. Biochem. Physiol. Part A Physiol. 1994, 109, 681–688. [Google Scholar] [CrossRef]
- Öner, M.; Atli, G.; Canli, M. Changes in serum biochemical parameters of freshwater fish Oreochromis niloticus following prolonged metal (Ag, Cd, Cr, Cu, Zn) exposures. Environ. Toxicol. Chem. Int. J. 2008, 27, 360–366. [Google Scholar] [CrossRef]
- Ren, M.C.; Liu, B.; Habte-Tsion, H.M.; Ge, X.P.; Xie, J.; Zhou, Q.L.; Liang, H.L.; Zhao, Z.X.; Pan, L.K. Dietary phenylalanine requirement and tyrosine replacement value for phenylalanine of juvenile blunt snout bream, Megalobrama amblycephala. Aquaculture 2015, 442, 51–57. [Google Scholar] [CrossRef]
- Liu, Y.L.; Lu, Q.S.; Xi, L.W.; Gong, Y.L.; Su, J.Z.; Han, D.; Zhang, Z.M.; Liu, H.K.; Jin, J.Y.; Yang, Y.X.; et al. Effects of replacement of dietary fishmeal by cottonseed protein concentrate on growth performance, liver health, and intestinal histology of largemouth bass (Micropterus salmoides). Front. Physiol. 2021, 12, 764987. [Google Scholar] [CrossRef] [PubMed]
- He, G.Z.; Zhang, T.T.; Zhou, X.M.; Liu, X.P.; Sun, H.; Chen, Y.J.; Tan, B.P.; Lin, S.M. Effects of cottonseed protein concentrate on growth performance, hepatic function and intestinal health in juvenile largemouth bass, Micropterus salmoides. Aquac. Rep. 2022, 23, 101052. [Google Scholar] [CrossRef]
- Xu, X.Y.; Yang, H.; Zhang, C.Y.; Bian, Y.H.; Yao, W.X.; Xu, Z.; Wang, Y.Y.; Li, X.Q.; Leng, X.J. Effects of replacing fishmeal with cottonseed protein concentrate on growth performance, flesh quality and gossypol deposition of largemouth bass (Micropterus salmoides). Aquaculture 2022, 548, 737551. [Google Scholar] [CrossRef]
- Xie, X.Z.; Wang, J.; Guan, Y.; Xing, S.J.; Liang, X.F.; Xue, M.; Wang, J.J.; Chang, Y.; Leclercq, E. Cottonseed protein concentrate as fishmeal alternative for largemouth bass (Micropterus salmoides) supplemented a yeast-based paraprobiotic: Effects on growth performance, gut health and microbiome. Aquaculture 2022, 551, 737898. [Google Scholar] [CrossRef]
- Bian, F.; Zhou, H.; He, G.; Wang, C.; Peng, H.; Pu, X.; Jiang, X.; Wang, X.; Mai, K. Effects of replacing fishmeal with different cottonseed meals on growth, feed utilization, haematological indexes, intestinal and liver morphology of juvenile turbot (Scophthalmus maximus L.). Aquac. Nutr. 2017, 23, 1429–1439. [Google Scholar] [CrossRef]
- Soyingbe, A.A.; Ogunyanwo, O.O.; Hammed, T.B.; Adesope, A.O. Effects of sublethal concentrations of diazinon on total protein in tilapia fish (Oreochromis niloticus). IOSR J. Environ. Sci. Toxicol. Food Technol. 2012, 1, 22–25. [Google Scholar] [CrossRef]
- Ren, M.C.; Liang, H.L.; He, J.; Masagounder, K.; Yue, Y.; Yang, H.; Ge, X.P.; Xi, B.W. Effects of DL-methionine supplementation on the success of fish meal replacement by plant proteins in practical diets for juvenile gibel carp (Carassius auratus gibelio). Aquac. Nutr. 2017, 23, 934–941. [Google Scholar] [CrossRef]
- Khosravi, S.; Bui, H.T.D.; Rahimnejad, S.; Herault, M.; Fournier, V.; Jeong, J.B.; Lee, K.J. Effect of dietary hydrolysate supplementation on growth performance, non-specific immune response and disease resistance of olive flounder (Paralichthys olivaceus) challenged with Edwardsiella tarda. Aquac. Nutr. 2015, 21, 321–331. [Google Scholar] [CrossRef]
- Zawistowski, J.; Kopec, A.; Kitts, D.D. Effects of a black rice extract (Oryza sativa L. indica) on cholesterol levels and plasma lipid parameters in Wistar Kyoto rats. J. Funct. Foods 2009, 1, 50–56. [Google Scholar] [CrossRef]
- Reinhart, K.M.; Talati, R.; White, C.M.; Coleman, C.I. The impact of garlic on lipid parameters: A systematic review and meta-analysis. Nutr. Res. Rev. 2009, 22, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.D.; Li, H.Y.; Wang, J.Y.; Li, P.Y.; Sun, Y.Z.; Zhang, L.M. Effects of fishmeal replacement with soy protein hydrolysates on growth performance, blood biochemistry, gastrointestinal digestion and muscle composition of juvenile starry flounder (Platichthys stellatus). Aquaculture 2014, 426, 96–104. [Google Scholar] [CrossRef]
- Ji, K.; Liang, H.L.; Ren, M.C.; Ge, X.P.; Mi, H.F.; Pan, L.K.; Yu, H. The immunoreaction and antioxidant capacity of juvenile blunt snout bream (Megalobrama amblycephala) involves the PI3K/Akt/Nrf2 and NF-κB signal pathways in response to dietary methionine levels. Fish Shellfish Immunol. 2020, 105, 126–134. [Google Scholar] [CrossRef]
- Zheng, Q.M.; Wen, X.B.; Han, C.Y.; Li, H.B.; Xie, X.H. Effect of replacing soybean meal with cottonseed meal on growth, hematology, antioxidant enzymes activity and expression for juvenile grass carp, Ctenopharyngodon idellus. Fish Physiol. Biochem. 2012, 38, 1059–1069. [Google Scholar] [CrossRef] [Green Version]
- Gaweł, S.; Wardas, M.; Niedworok, E.; Wardas, P. Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad. Lek. 2004, 57, 453–455. [Google Scholar]
- Yuan, X.Y.; Liu, W.B.; Wang, C.C.; Huang, Y.Y.; Dai, Y.J.; Cheng, H.H.; Jiang, G.Z. Evaluation of antioxidant capacity and immunomodulatory effects of cottonseed meal protein hydrolysate and its derivative peptides for hepatocytes of blunt snout bream (Megalobrama amblycephala). Fish Shellfish Immunol. 2020, 98, 10–18. [Google Scholar] [CrossRef]
- Gao, D.D.; Cao, Y.S.; Li, H.X. Antioxidant activity of peptide fractions derived from cottonseed protein hydrolysate. J. Sci. Food Agric. 2010, 90, 1855–1860. [Google Scholar] [CrossRef]
- Song, W.G.; Kong, X.Z.; Hua, Y.F.; Li, X.F.; Zhang, C.M.; Chen, Y.M. Antioxidant and antibacterial activity and in vitro digestion stability of cottonseed protein hydrolysates. LWT 2020, 118, 108724. [Google Scholar] [CrossRef]
- Bu, X.Y.; Chen, A.J.; Lian, X.Q.; Chen, F.Y.; Zhang, Y.; Muhammad, I.; Ge, X.P.; Yang, Y.H. An evaluation of replacing fish meal with cottonseed meal in the diet of juvenile Ussuri catfish Pseudobagrus ussuriensis: Growth, antioxidant capacity, nonspecific immunity and resistance to Aeromonas hydrophila. Aquaculture 2017, 479, 829–837. [Google Scholar] [CrossRef]
- Boboev, A.; Hasanov, A.; Yotova, L.; Hasanov, H. Antioxidant activity of peptides obtained from wheat and cottonseed proteins. Bulg. J. Agric. Sci. 2012, 18, 103–111. [Google Scholar]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Li, C.W.; Li, L.L.; Chen, S.; Zhang, J.X.; Lu, W.L. Antioxidant nanotherapies for the treatment of inflammatory diseases. Front. Bioeng. Biotechnol. 2020, 8, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Dong, X.H.; Tan, B.P.; Du, T.; Zhang, S.; Yang, Y.Z.; Chi, S.Y.; Yang, Q.H.; Liu, H.Y. Effects of fish meal replacement by low-gossypol cottonseed meal on growth performance, digestive enzyme activity, intestine histology and inflammatory gene expression of silver sillago (Sillago sihama Forsskál)(1775). Aquac. Nutr. 2020, 26, 1724–1735. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis of the Association of Official Analytical Chemists, 15th ed.; Association of Official Analytical Chemists Inc.: Arlington, TX, USA, 1998. [Google Scholar]
- Huang, D.Y.; Liang, H.L.; Ge, X.P.; Zhu, J.; Li, S.L.; Wang, Y.L.; Ren, M.C.; Chen, X.L. Effects of Dietary Lysine Levels on Growth Performance and Glycolipid Metabolism via the AKT/FoxO1 Pathway in Juvenile Largemouth Bass, Micropterus salmoides. Aquac. Nutr. 2022, 2022, 1372819. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.L.; Liang, J.; Chen, F.K.; Tang, X.H.; Liao, L.; Liu, Q.; Luo, J.; Du, Z.J.; Li, Z.Q.; Luo, W.; et al. High carbohydrate diet induced endoplasmic reticulum stress and oxidative stress, promoted inflammation and apoptosis, impaired intestinal barrier of juvenile largemouth bass (Micropterus salmoides). Fish Shellfish Immunol. 2021, 119, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Liang, H.L.; Ge, X.P.; Zhu, J.; Wang, Y.L.; Ren, M.C.; Chen, X.R. Dietary chlorella (Chlorella vulgaris) supplementation effectively improves body color, alleviates muscle inflammation and inhibits apoptosis in largemouth bass (Micropterus salmoides). Fish Shellfish Immunol. 2022, 127, 140–147. [Google Scholar] [CrossRef]
- Gu, J.Z.; Liang, H.L.; Ge, X.P.; Xia, D.; Pan, L.K.; Mi, H.F.; Ren, M.C. A study of the potential effect of yellow mealworm (Tenebrio molitor) substitution for fish meal on growth, immune and antioxidant capacity in juvenile largemouth bass (Micropterus salmoides). Fish Shellfish Immunol. 2022, 120, 214–221. [Google Scholar] [CrossRef]
- Yang, P.; Wang, W.Q.; Chi, S.Y.; Mai, K.S.; Song, F.; Wang, L. Effects of dietary lysine on regulating GH-IGF system, intermediate metabolism and immune response in largemouth bass (Micropterus salmoides). Aquac. Rep. 2020, 17, 100323. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Diets | IW (g) 2 | FW (g) 3 | WGR (%) 4 | SGR (%/day) 5 | FI (g/fish) 6 | FCR 7 | PER 8 | SR (%) 9 |
|---|---|---|---|---|---|---|---|---|
| ECP0 | 14.88 ± 0.06 | 72.82 ± 2.66 ab | 389.32 ± 19.31 a | 2.65 ± 0.07 a | 72.09 ± 1.15 ab | 1.26 ± 0.06 | 1.59 ± 0.08 | 95.00 ± 5.00 |
| ECP3.6 | 15.03 ± 0.12 | 76.77 ± 0.86 a | 411.75 ± 0.88 a | 2.72 ± 0.00 a | 70.95 ± 2.94 ab | 1.25 ± 0.10 | 1.61 ± 0.12 | 91.67 ± 5.77 |
| ECP7.2 | 15.00 ± 0.13 | 71.11 ± 0.61 b | 374.12 ± 8.08 a | 2.59 ± 0.03 a | 73.41 ± 1.33 a | 1.33 ± 0.05 | 1.52 ± 0.05 | 95.00 ± 8.66 |
| ECP10.8 | 15.08 ± 0.03 | 60.78 ± 0.75 c | 303.20 ± 5.93 b | 2.33 ± 0.02 b | 66.90 ± 2.21 b | 1.37 ± 0.14 | 1.46 ± 0.15 | 98.33 ± 2.89 |
| ECP14.4 | 14.97 ± 0.18 | 64.22 ± 2.07 c | 329.22 ± 16.57 b | 2.43 ± 0.07 b | 70.20 ± 2.08 ab | 1.46 ± 0.02 | 1.37 ± 0.02 | 88.33 ± 5.77 |
| Diets | Moisture (%) | Crude Protein (%) | Crude Lipid (%) | Ash (%) |
|---|---|---|---|---|
| ECP0 | 67.53 ± 1.46 | 17.52 ± 0.01 | 7.95 ± 0.73 | 4.62 ± 0.19 |
| ECP3.6 | 67.40 ± 1.04 | 17.73 ± 0.47 | 8.55 ± 0.94 | 4.55 ± 0.36 |
| ECP7.2 | 67.74 ± 1.01 | 17.64 ± 0.26 | 8.06 ± 0.34 | 4.55 ± 0.16 |
| ECP10.8 | 68.59 ± 0.27 | 17.79 ± 0.06 | 8.07 ± 0.45 | 4.95 ± 0.23 |
| ECP14.4 | 66.33 ± 1.57 | 18.68 ± 0.18 | 8.97 ± 0.21 | 4.89 ± 0.27 |
| Diets | TP (g/L) 2 | ALB (g/L) 2 | TC (mmol/L) 2 | TG (mmol/L) 2 | LDL (mmol/L) 2 | HDL (mmol/L) 2 | ALP (U/L) 2 |
|---|---|---|---|---|---|---|---|
| ECP0 | 55.72 ± 7.21 c | 13.61 ± 2.10 c | 12.13 ± 2.24 | 10.00 ± 1.97 ab | 4.91 ± 1.60 a | 2.37 ± 0.28 c | 194.87 ± 52.78 a |
| ECP3.6 | 61.48 ± 4.68 bc | 15.62 ± 1.81 bc | 12.02 ± 1.45 | 9.56 ± 1.23 ab | 4.63 ± 0.87 ab | 2.76 ± 0.22 b | 161.16 ± 35.10 abc |
| ECP7.2 | 69.72 ± 4.04 a | 17.34 ± 1.14 a | 12.55 ± 1.06 | 10.97 ± 1.29 a | 4.78 ± 0.68 a | 3.01 ± 0.13 ab | 169.93 ± 31.01 ab |
| ECP10.8 | 67.74 ± 5.98 ab | 17.42 ± 1.39 ab | 11.63 ± 1.35 | 8.20 ± 0.92 b | 3.92 ± 0.83 ab | 3.18 ± 0.38 a | 143.72 ± 19.23 bc |
| ECP14.4 | 67.10 ± 4.30 ab | 17.92 ± 1.54 a | 10.66 ± 1.65 | 6.09 ± 1.20 c | 3.24 ± 1.17 b | 2.96 ± 0.12 ab | 119.13 ± 32.61 c |
| Ingredients (%) | Diets | ||||
|---|---|---|---|---|---|
| ECP0 | ECP3.6 | ECP7.2 | ECP10.8 | ECP14.4 | |
| Fish meal 1 | 45.00 | 41.50 | 38.00 | 34.50 | 31.00 |
| Enzymatic cottonseed protein concentrate 1 | 0.00 | 3.60 | 7.20 | 10.80 | 14.40 |
| Blood meal 1 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Soybean meal 1 | 13.00 | 13.00 | 13.00 | 13.00 | 13.00 |
| Corn gluten meal1 | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 |
| Enzymatic hydrolysis of chicken powder 1 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 |
| Wheat meal | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 |
| Cassava starch | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 |
| Rice bran | 6.13 | 6.13 | 6.13 | 6.13 | 6.13 |
| Microcrystalline cellulose | 3.27 | 2.41 | 1.58 | 0.82 | 0.00 |
| Squid Ointment | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Fish oil | 4.10 | 4.45 | 4.75 | 5.00 | 5.30 |
| Mineral premix 2 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Vitamin premix 2 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Monocalcium phosphate | 1.00 | 1.30 | 1.60 | 1.90 | 2.20 |
| Choline chloride | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| L-Lysine 3 | 0.00 | 0.08 | 0.17 | 0.25 | 0.33 |
| L-Methionine 3 | 0.00 | 0.03 | 0.07 | 0.10 | 0.14 |
| Taurine (mg/kg) | 0.00 | 13.30 | 26.60 | 39.90 | 53.20 |
| Proximate Composition (dry basis) | |||||
| Crude protein (%) | 48.59 | 48.53 | 48.61 | 48.47 | 48.49 |
| Crude lipid (%) | 9.25 | 9.24 | 8.98 | 9.28 | 9.23 |
| Gross energy (KJ/g) | 19.75 | 19.83 | 19.87 | 19.72 | 19.58 |
| Items | Methods, Assay Kits and Testing Equipment |
|---|---|
| Crude protein | Determined by Hanon K1100 auto kieldahl apparatus (Jinan Hanon Instruments Co., Ltd., Jinan, China). |
| Crude lipid | Determined by Hanon SOX606 auto fat analy (Jinan Hanon Instruments Co., Ltd., Jinan, China). |
| Ash | Determined by burning at 550 ℃ for 5 h in a XL-2A intelligent muffle furnace (Hangzhou Zhuochi Instruments Co., Ltd., Hangzhou, China). |
| Gross energy | Measured by an oxygen bomb calorimeter IKA C6000 ((IKA Works Guangzhou, Guangzhou, China). |
| TP 1 | All plasma parameters were determined by Assay kits (Mindray Bio Medical Co., Ltd., Shenzhen, China) with a Mindray BS-400 automatic biochemical analyzer (Mindray Medical International Ltd., Shenzhen, China). |
| ALB 1 | |
| TC 1 | |
| TG 1 | |
| LDL 1 | |
| HDL 1 | |
| ALP 1 | |
| T-SOD 2 | All hepatic antioxidant parameters and MDA levels were tested according to the instructions of assay kits purchased from Jian Cheng Bioengineering Institute (Nanjing, China). |
| CAT 2 | |
| GSH 2 | |
| MDA 2 |
| Gene 1 | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) | Source |
|---|---|---|---|
| Nrf2 | CTGGTCCGAGACATACGC | CTCAGCAGACGCTCCTTC | Zhao et al. (2021) [59] |
| Keap1 | CGTACGTCCAGGCCTTACTC | TGACGGAAATAACCCCCTGC | Yu et al. (2022) [60] |
| SOD | TGGCAAGAACAAGAACCACA | CCTCTGATTTCTCCTGTCACC | Gu et al. (2022) [61] |
| CAT | CTATGGCTCTCACACCTTC | TCCTCTACTGGCAGATTCT | Gu et al. (2022) [61] |
| NF-κB | CCACTCAGGTGTTGGAGCTT | TCCAGAGCACGACACACTTC | Yu et al. (2022) [60] |
| IL-8 | CGTTGAACAGACTGGGAGAGATG | AGTGGGATGGCTTCATTATCTTGT | Yang et al. (2020) [62] |
| TNF-α | CTTCGTCTACAGCCAGGCATCG | TTTGGCACACCGACCTCACC | Gu et al. (2022) [61] |
| TGF-β | GCTCAAAGAGAGCGAGGATG | TCCTCTACCATTCGCAATCC | Gu et al. (2022) [61] |
| IL-10 | CGGCACAGAAATCCCAGAGC | CAGCAGGCTCACAAAATAAACATCT | Gu et al. (2022) [61] |
| GAPDH | ACTGTCACTCCTCCATCTT | CACGGTTGCTGTATCCAA | Yu et al. (2022) [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Liang, H.; Xu, P.; Xu, G.; Zhang, L.; Wang, Y.; Ren, M.; Chen, X. Effects of Enzymatic Cottonseed Protein Concentrate as a Feed Protein Source on the Growth, Plasma Parameters, Liver Antioxidant Capacity and Immune Status of Largemouth Bass (Micropterus salmoides). Metabolites 2022, 12, 1233. https://doi.org/10.3390/metabo12121233
Zhang Q, Liang H, Xu P, Xu G, Zhang L, Wang Y, Ren M, Chen X. Effects of Enzymatic Cottonseed Protein Concentrate as a Feed Protein Source on the Growth, Plasma Parameters, Liver Antioxidant Capacity and Immune Status of Largemouth Bass (Micropterus salmoides). Metabolites. 2022; 12(12):1233. https://doi.org/10.3390/metabo12121233
Chicago/Turabian StyleZhang, Qile, Hualiang Liang, Pao Xu, Gangchun Xu, Lu Zhang, Yongli Wang, Mingchun Ren, and Xiaoru Chen. 2022. "Effects of Enzymatic Cottonseed Protein Concentrate as a Feed Protein Source on the Growth, Plasma Parameters, Liver Antioxidant Capacity and Immune Status of Largemouth Bass (Micropterus salmoides)" Metabolites 12, no. 12: 1233. https://doi.org/10.3390/metabo12121233