

Bioactive Compounds from Endophytic Bacteria Bacillus subtilis Strain EP1 with Their Antibacterial Activities

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Purification
2.2. DNA Extraction, Sequencing, and Identification
2.3. Extraction of Biomass
2.4. Extraction and Isolation of Pure Compounds
2.5. Antibacterial Activities
2.6. Molecular Validation of the Antibacterial Activities
2.6.1. RNA Extraction
2.6.2. cDNA Synthesis and QRT PCR Analysis
3. Results and Discussion
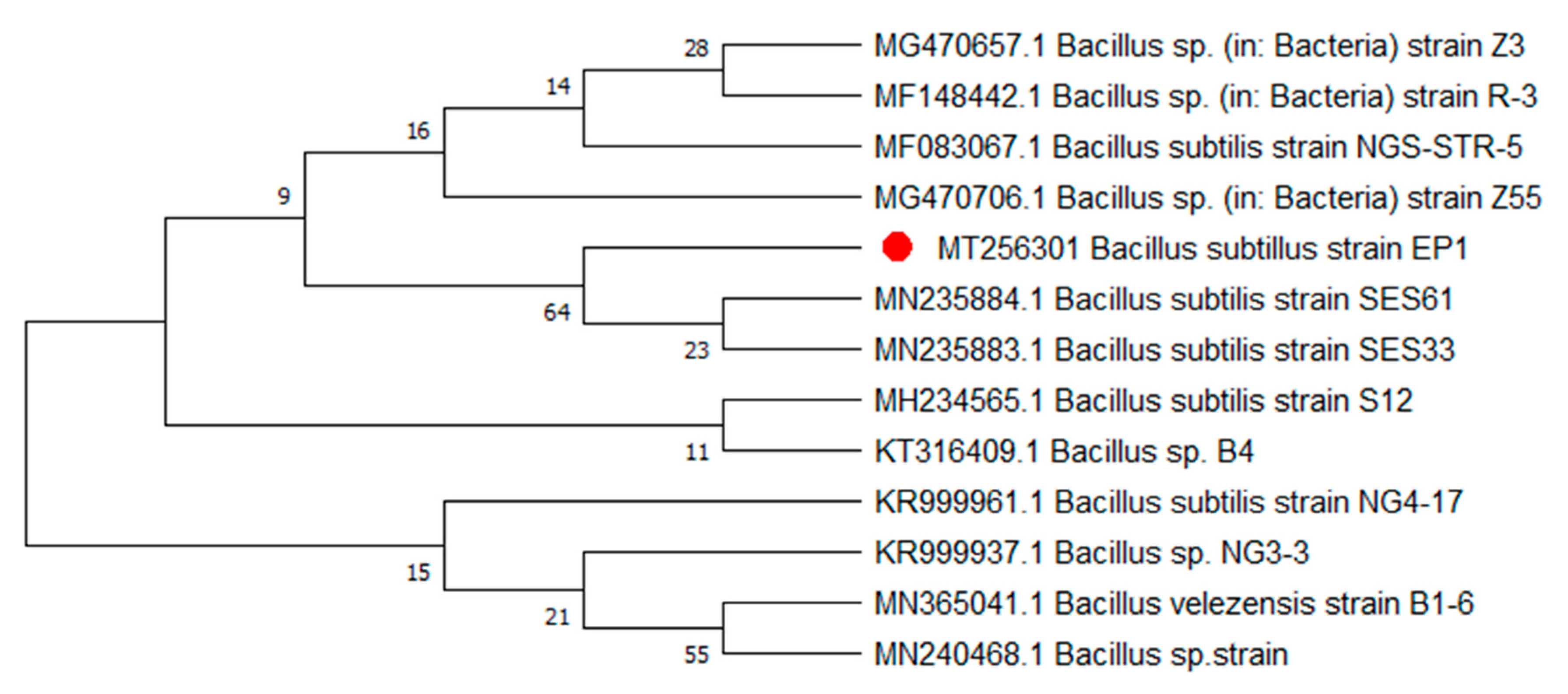
3.1. Strain Identification
3.2. Characterization of Compounds through NMR and Mass Spectroscopy
3.3. Antibacterial Significance
3.4. Beta-Lactam Gene Inhibition
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Rho, H.; Hsieh, M.; Kandel, S.L.; Cantillo, J.; Doty, S.L.; Kim, S.-H. Do endophytes promote growth of host plants under stress? A meta-analysis on plant stress mitigation by endophytes. Micro. Ecol. 2018, 75, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Kia, S.H.; Glynou, K.; Nau, T.; Thines, M.; Piepenbring, M.; Maciá-Vicente, J.G. Influence of phylogenetic conservatism and trait convergence on the interactions between fungal root endophytes and plants. ISME J. 2017, 11, 777–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Defez, R.; Andreozzi, A.; Bianco, C. The overproduction of indole-3-acetic acid (IAA) in endophytes upregulates nitrogen fixation in both bacterial cultures and inoculated rice plants. Microb. Ecol. 2017, 74, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Passari, A.K.; Mishra, V.K.; Singh, G.; Singh, P.; Kumar, B.; Gupta, V.K.; Sarma, R.K.; Saikia, R.; Singh, B.P. Insights into the functionality of endophytic actinobacteria with a focus on their biosynthetic potential and secondary metabolites production. Sci. Rep. 2017, 7, 11809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isah, T.; Umar, S.; Mujib, A.; Sharma, M.P.; Rajasekharan, P.; Zafar, N.; Frukh, A. Secondary metabolism of pharmaceuticals in the plant in vitro cultures: Strategies, approaches, and limitations to achieving higher yield. Plant Cell Tiss. Org. Cult. 2018, 132, 239–265. [Google Scholar] [CrossRef]
- Saha, P.; Talukdar, A.D.; Choudhury, M.D.; Nath, D. Bioprospecting for Fungal-Endophyte-Derived Natural Products for Drug Discovery. Adv. Endo. Fungal Res. 2019, 2, 35–49. [Google Scholar]
- Rajamanikyam, M.; Vadlapudi, V.; Upadhyayula, S.M. Endophytic fungi as novel resources of natural therapeutics. Brazilian Arch. Biol. Technol. 2017, 60, e17160542. [Google Scholar] [CrossRef] [Green Version]
- Numan, M.; Bashir, S.; Mumtaz, R.; Tayyab, S.; Rehman, N.U.; Khan, A.L.; Shinwari, Z.K.; Al-Harrasi, A. Therapeutic applications of bacterial pigments: A review of current status and future opportunities. 3 Biotech. 2018, 8, 207. [Google Scholar] [CrossRef]
- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J.; Dowling, D.N. Bacterial endophytes: Recent developments and applications. FEMS Microbiol. Lett. 2008, 278, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mumtaz, R.; Bashir, S.; Numan, M.; Shinwari, Z.K.; Ali, M. Pigments from Soil Bacteria and Their Therapeutic Properties: A Mini Review. Curr. Microbiol. 2019, 76, 783–790. [Google Scholar] [CrossRef]
- Singh, R.K.; Shrivastava, A.; Yadav, A.; Srivastava, A.K. Endophytic bacteria as a source of bioactive compounds. In Microbial Endophytes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 175–188. [Google Scholar]
- Takahashi, Y.; Nakashima, T. Actinomycetes, an inexhaustible source of naturally occurring antibiotics. Antibiotics 2018, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Rahman, H.; Khan, A.; Bibi, S.; Ullah, O.; Ullah, S.; Ur Rehman, N.; Murad, W.; Al-Harrasi, A. Identification of α-Glucosidase Inhibitors from Scutellaria edelbergii: ESI-LC-MS and Computational Approach. Molecules 2022, 27, 1322. [Google Scholar] [CrossRef]
- Mohamad, O.A.; Li, L.; Ma, J.B.; Hatab, S.; Xu, L.; Guo, J.W.; Rasulov, B.A.; Liu, Y.H.; Hedlund, B.P.; Li, W.J. Evaluation of the antimicrobial activity of endophytic bacterial populations from Chinese traditional medicinal plant licorice and characterization of the bioactive secondary metabolites produced by Bacillus atrophaeus against Verticillium dahliae. Front. Microbiol. 2018, 9, 924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götmark, F.; Cafaro, P.; O’Sullivan, J. Aging human populations: Good for us, good for the Earth. Trends Ecol. Evol. 2018, 33, 851–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra, S.A.; Rather, M.I.; Para, P.A.; Ganguly, S. The emergence of drug resistant bacteria: Effects on human health. J. Environ. Life Sci. 2017, 2, 77–79. [Google Scholar]
- Windels, E.M.; Michiels, J.E.; Fauvart, M.; Wenseleers, T.; Van den Bergh, B.; Michiels, J. Bacterial persistence promotes the evolution of antibiotic resistance by increasing survival and mutation rates. ISME J. 2019, 13, 1239–1251. [Google Scholar] [CrossRef]
- Subramani, R.; Narayanasamy, M.; Feussner, K.-D. Plant-derived antimicrobials to fight against multi-drug-resistant human pathogens. 3 Biotech. 2017, 7, 172. [Google Scholar] [CrossRef]
- Mubin, S.; Rehman, N.U.; Murad, W.; Shah, M.; Al-Harrasi, A.; Afza, R. Scutellaria petiolata Hemsl. ex Lace & Prain (Lamiaceae).: A New Insight in Biomedical Therapies. Antioxidants 2022, 11, 1446. [Google Scholar]
- Singh, M.; Kumar, A.; Singh, R.; Pandey, K.D. Endophytic bacteria: A new source of bioactive compounds. 3 Biotech. 2017, 7, 315. [Google Scholar] [CrossRef]
- Shah, M.; Mubin, S.; Tagde, P.; Ullah, O.; Rahman, M.; Al-Harrasi, A.; Rehman, N.U.; Murad, W. Phytochemical Profiling and Bio-Potentiality of Genus Scutellaria: Biomedical Approach. Biomolecules 2022, 12, 936. [Google Scholar] [CrossRef]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial metabolites in nutrition, healthcare and agriculture. 3 Biotech. 2017, 7, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.; Murad, W.; Mubin, S.; Ullah, O.; Rehman, N.U.; Rahman, M. Multiple health benefits of curcumin and its therapeutic potential. Environ. Sci. Pollut. Res. 2022, 29, 43732–43744. [Google Scholar] [CrossRef]
- Das, A.; Rahman, M.I.; Ferdous, A.S.; Amin, A.; Rahman, M.M.; Nahar, N.; Uddin, M.A.; Islam, M.R.; Khan, H. An endophytic Basidiomycete, Grammothele lineata, isolated from Corchorus olitorius, produces paclitaxel that shows cytotoxicity. PLoS ONE 2017, 12, e0178612. [Google Scholar] [CrossRef]
- Zabalgogeazcoa, I.; Alvarez, A.; Herrero, N.; Vazquez-de-Aldana, B.R. Production of fumonisins by endophytic strains of Tolypocladium cylindrosporum and its relation to fungal virus infection. Mycotoxin Res. 2018, 34, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Christina, A.; Christapher, V.; Bhore, S.J. Endophytic bacteria as a source of novel antibiotics: An overview. Pharmacogn. Rev. 2013, 7, 11. [Google Scholar]
- Jain, A.; Dixit, P. Multidrug resistant to extensively drug resistant tuberculosis: What is next? J. Biosci. 2008, 33, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Barzanti, R.; Ozino, F.; Bazzicalupo, M.; Gabbrielli, R.; Galardi, F.; Gonnelli, C.; Mengoni, A. Isolation and characterization of endophytic bacteria from the nickel hyperaccumulator plant Alyssum bertolonii. Microb. Ecol. 2007, 53, 306–316. [Google Scholar] [CrossRef]
- Arnold, A.E.; Engelbrecht, B.M. Fungal endophytes nearly double minimum leaf conductance in seedlings of a neotropical tree species. J. Trop. Ecol. 2007, 23, 369–372. [Google Scholar] [CrossRef] [Green Version]
- Numan, M.; Bashir, S.; Mumtaz, R.; Tayyab, S.; Ullah, I.; Khan, A.L.; Shinwari, Z.K.; Al-Harrasi, A. Chemical profile and in-vitro pharmacological activities of yellow pigment extracted from Arthrobacter gandavensis. Process Biochem. 2018, 75, 74–82. [Google Scholar] [CrossRef]
- Radjasa, O.K.; Kencana, D.S.; Sabdono, A.; Hutagalung, R.A.; Lestari, E.S. Antibacterial activity of marine bacteria associated with sponge Aaptos sp. against multi drugs resistant (MDR) Strains. J. Mat. Sains 2009, 12, 147–152. [Google Scholar]
- Radjasa, O.K.; Sabdono, A.; Junaidi, J.; Zocchi, E. Richness of Secondary Metabolite-Producing Marine Bacteria Associated with Sponge Hatictona sp. Int. J. Pharmacol. 2007, 3, 275–279. [Google Scholar]
- Li, F.; Xue, F.; Yu, X. GC–MS, FTIR and Raman Analysis of Antioxidant Components of Red Pigments from Stemphylium lycopersici. Curr. Microbiol. 2017, 74, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Raina, J.-B.; Tapiolas, D.; Motti, C.A.; Foret, S.; Seemann, T.; Tebben, J.; Willis, B.L.; Bourne, D.G. Isolation of an antimicrobial compound produced by bacteria associated with reef-building corals. PeerJ 2016, 4, e2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ríos-Marco, P.; Romero-López, C.; Berzal-Herranz, A. The cis-acting replication element of the Hepatitis C virus genome recruits host factors that influence viral replication and translation. Sci. Rep. 2016, 6, 25729. [Google Scholar] [CrossRef] [Green Version]
- Green, M.R.; Sambrook, J. Synthesis of cDNA Probes from mRNA Using Random Oligonucleotide Primers. Cold Spring Harbor Protocols 2019, 2019, pdb-prot100636. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.; Zhao, X.; Tang, C.; Wang, Z.; Kuang, Z.; Li, Z.; Huang, J.; Luo, G. Identification and validation of reference genes for qRT-PCR analysis in mulberry (Morus alba L.). PLoS ONE 2018, 13, e0194129. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Suring, W.; Mariën, J.; Broekman, R.; Van Straalen, N.M.; Roelofs, D. Biochemical pathways supporting beta-lactam biosynthesis in the springtail Folsomia candida. Biol. Open 2016, 5, 1784–1789. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Gao, Y.; Wang, Y.; Wu, Y.; Wāng, Y.; Zhao, Y.; Chen, H.; Bao, D.; Xu, J.; Bian, X. Selection and evaluation of appropriate reference genes for RT-qPCR normalization of Volvariella volvacea gene expression under different conditions. BioMed. Res. Int. 2018, 2018, 6125706. [Google Scholar] [CrossRef] [Green Version]
- Wattana-Amorn, P.; Charoenwongsa, W.; Williams, C.; Crump, M.P.; Apichaisataienchote, B. Antibacterial activity of cyclo (L-Pro-L-Tyr) and cyclo (D-Pro-L-Tyr) from Streptomyces sp. strain 22-4 against phytopathogenic bacteria. Nat. Prod. Res. 2016, 30, 1980–1983. [Google Scholar] [CrossRef] [Green Version]
- Alshaibani, M.M.; MohamadZin, N.; Jalil, J.; Sidik, N.M.; Ahmad, S.J.; Kamal, N.; Edrada-Ebel, R. Isolation, purification, and characterization of five active diketopiperazine derivatives from endophytic Streptomyces SUK 25 with antimicrobial and cytotoxic activities. J. Microbiol. Biotechnol. 2017, 27, 1249–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Hosni, S.; Rizvi, T.S.; Khan, A.; Ali, L.; Rehman, N.U.; Rashid, U.; Hussian, J.; Khan, A.L.; Al-Harrasi, A. Diketopeprazin and Methyl-5-docosenoate from endophytic fungi Aureobasidium pollulan BSS6 with α-glucosidase inhibition and its validation through molecular docking. South Afr. J. Bot. 2020, 134, 322–328. [Google Scholar] [CrossRef]
- Lin, W.-X.; Xie, C.-L.; Zhou, M.; Xia, M.-L.; Zhou, T.-T.; Chen, H.-F.; Yang, X.-W.; Yang, Q. Chemical constituents from the deep sea-derived Streptomyces xiamenensis MCCC 1A01570 and their effects on RXRα transcriptional regulation. Nat. Prod. Res. 2018, 34, 1461–1464. [Google Scholar] [CrossRef]
- Abed, R.M.; Dobretsov, S.; Al-Fori, M.; Gunasekera, S.P.; Sudesh, K.; Paul, V.J. Quorum-sensing inhibitory compounds from extremophilic microorganisms isolated from a hypersaline cyanobacterial mat. J. Ind. Microbiol. Biotechnol. 2013, 40, 759–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, N.; Mohandas, C.; Nambisan, B.; Kumar, D.S.; Lankalapalli, R.S. Isolation of proline-based cyclic dipeptides from Bacillus sp. N strain associated with rhabitid entomopathogenic nematode and its antimicrobial properties. World J. Microbiol. Biotechnol. 2013, 29, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.N.; Nambisan, B.; Mohandas, C. Purification and identification of two antifungal cyclic dipeptides from Bacillus cereus subsp. thuringiensis associated with a rhabditid entomopathogenic nematode especially against Fusarium oxysporum. J. Enzym. Inhib. Med. Chem. 2014, 29, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.N.; Mohandas, C.; Nambisan, B. Purification of an antifungal compound, cyclo (L-Pro-D-Leu) for cereals produced by Bacillus cereus subsp. thuringiensis associated with entomopathogenic nematode. Microbiol. Res. 2013, 168, 278–288. [Google Scholar] [CrossRef]
- Gos, F.M.; Savi, D.C.; Shaaban, K.A.; Thorson, J.S.; Aluizio, R.; Possiede, Y.M.; Rohr, J.; Glienke, C. Antibacterial activity of endophytic actinomycetes isolated from the medicinal plant Vochysia divergens (Pantanal, Brazil). Front. Microbiol. 2017, 8, 1642. [Google Scholar] [CrossRef]
- Nishanth Kumar, S.; Mohandas, C.; Siji, J.; Rajasekharan, K.; Nambisan, B. Identification of antimicrobial compound, diketopiperazines, from a B acillus sp. N strain associated with a rhabditid entomopathogenic nematode against major plant pathogenic fungi. J. Appl. Microbiol. 2012, 113, 914–924. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Beta-Lactam Biosynthesis | |||
|---|---|---|---|
| Gene Name | Code | F | Reference |
| δ-(l-α-aminoadipyl)-l-cysteinyl-d-valine synthetase | ACVS F | AGCCGTGAAAAGCCACTTGA | [39] |
| ACVS R | GCTCACTGATGCCAATGGTTC | ||
| aminoadipate aminotransferase | AADAT F | GTGGCATCGGTAGAGAAGCATT | |
| AADAT R | CAACGTGAAGCATTCTCATCCA | ||
| Ubiquitin | UBQ F | CAATCACCTTGGAAGTCGAGTCGTC | [40] |
| UBQ R | CTGGATGTTGTAGTCGGAAAGGGTG | ||
| 1 | 2 | 3 | 4 | |
|---|---|---|---|---|
| Concentration | (Mean ± SD in mm) | |||
| Amoxicillin | 24 ± 1 | 23 ± 1 | 20 ± 1 | 26 ± 1 |
| 1 mg/mL | 11 ± 1 | 10 ± 1 | 11 ± 0.5 | 13 ± 1 |
| 500 µg/mL | 9 ± 1 | 10 ± 0.5 | 10 ± 0.5 | 11 ± 0.5 |
| 250 µg/mL | 9 ± 1 | 9 ± 0.5 | 9 ± 0.5 | 10 ± 0.5 |
| 100 µg/mL | 7 ± 0.5 | 9 ± 1 | 10 ± 0.5 | 9 ± 0.5 |
| 50 µg/mL | 8 ± 1 | 8 ± 0.5 | 8 ± 0.5 | 9 ± 0.5 |
| 10 µg/mL | 7 ± 1 | 7 ± 0.5 | 7 ± 0.5 | 8 ± 0.5 |
| DMSO | 6 ± 0.5 | 6 ± 0.5 | 6 ± 0.5 | 6 ± 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Numan, M.; Shah, M.; Asaf, S.; Ur Rehman, N.; Al-Harrasi, A. Bioactive Compounds from Endophytic Bacteria Bacillus subtilis Strain EP1 with Their Antibacterial Activities. Metabolites 2022, 12, 1228. https://doi.org/10.3390/metabo12121228
Numan M, Shah M, Asaf S, Ur Rehman N, Al-Harrasi A. Bioactive Compounds from Endophytic Bacteria Bacillus subtilis Strain EP1 with Their Antibacterial Activities. Metabolites. 2022; 12(12):1228. https://doi.org/10.3390/metabo12121228
Chicago/Turabian StyleNuman, Muhammad, Muddaser Shah, Sajjad Asaf, Najeeb Ur Rehman, and Ahmed Al-Harrasi. 2022. "Bioactive Compounds from Endophytic Bacteria Bacillus subtilis Strain EP1 with Their Antibacterial Activities" Metabolites 12, no. 12: 1228. https://doi.org/10.3390/metabo12121228