
Lipid Serum Profiling of Boar-Tainted and Untainted Pigs Using GC×GC–TOFMS: An Exploratory Study

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Chemicals
2.2. Instrumental Method
2.3. Sample Preparation
2.4. Data Processing
3. Results
3.1. Optimization of Derivatization and Extraction Conditions via Experimental Design
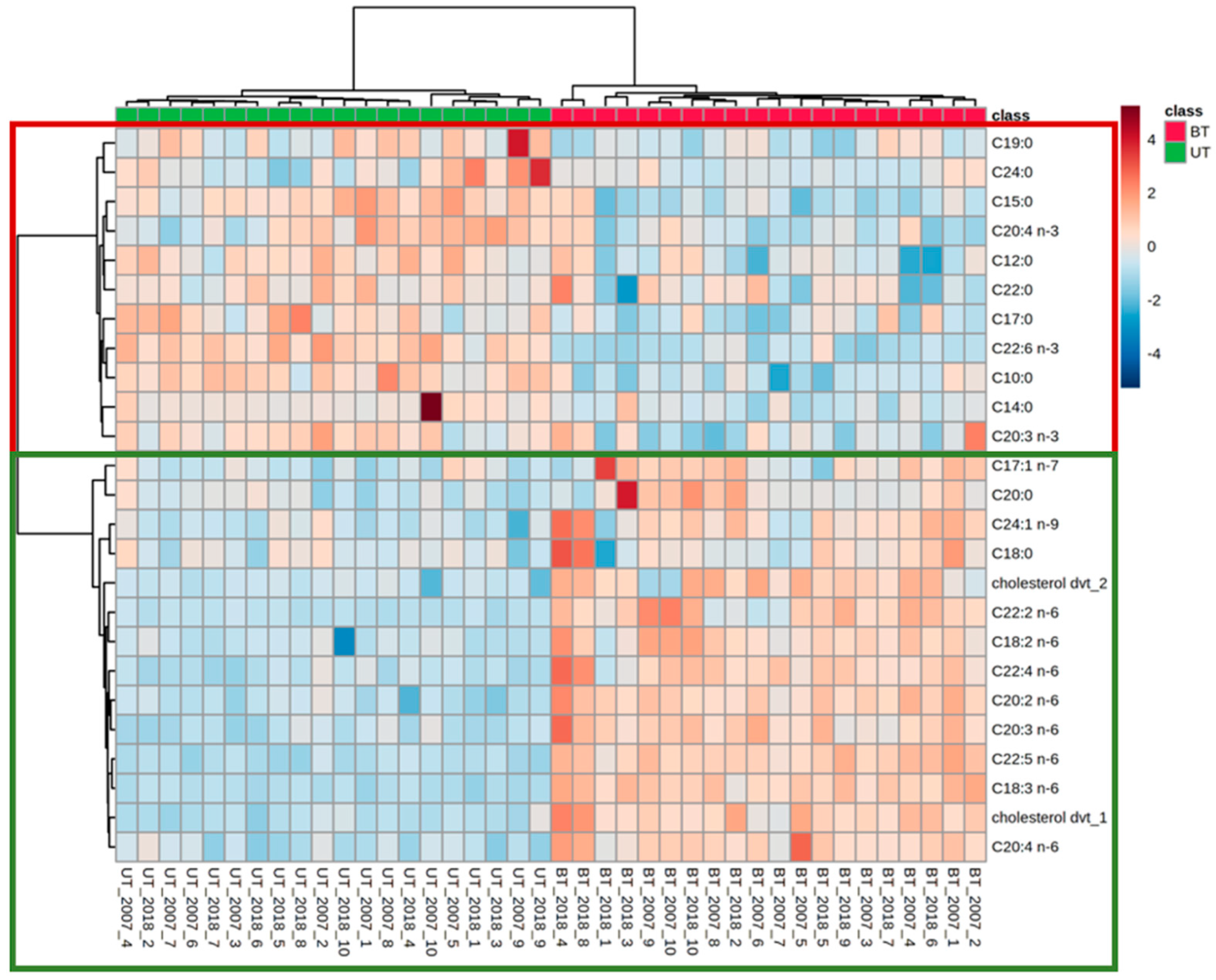
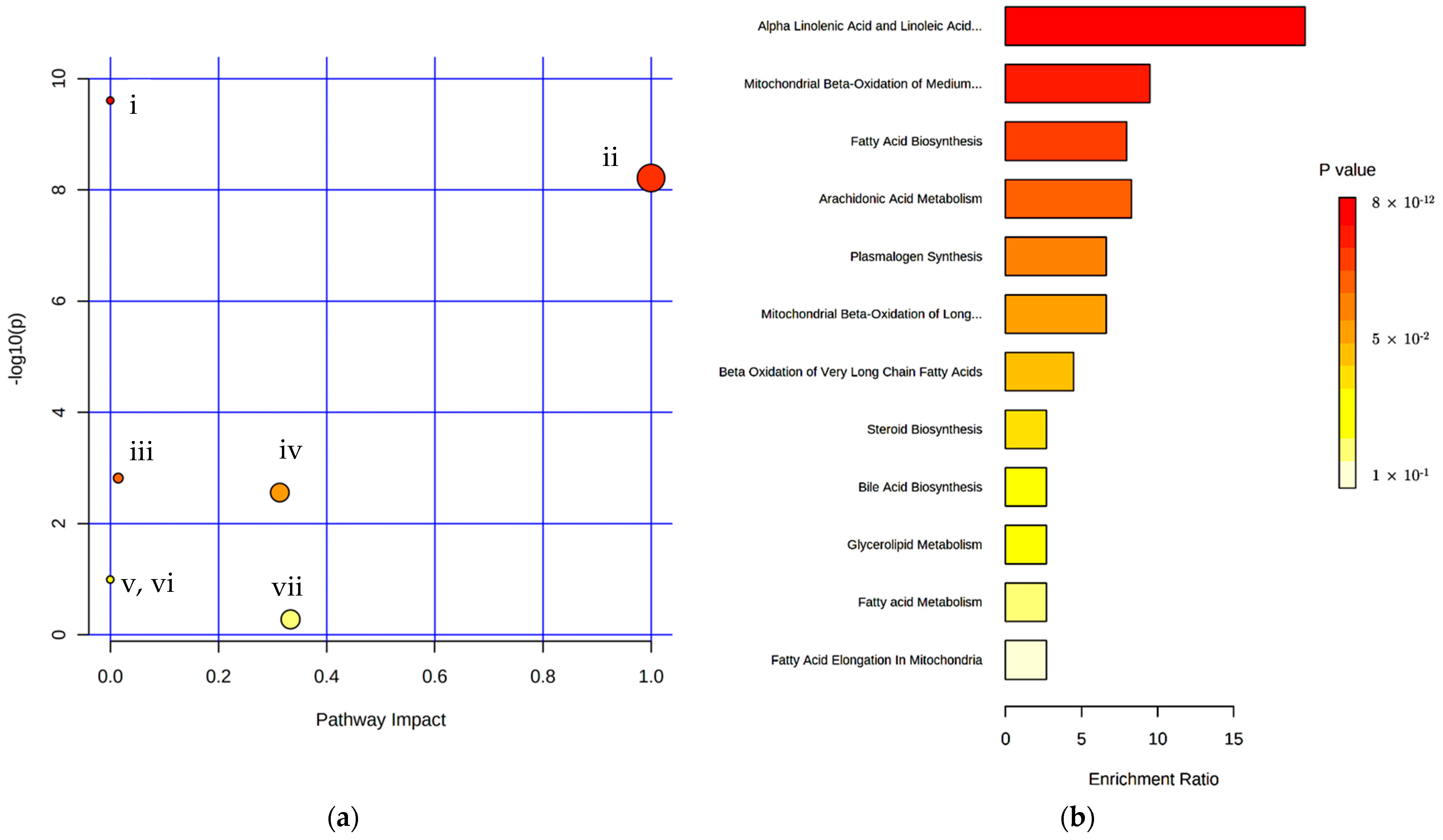
3.2. Identification of Pigs Responsible for Boar Taint by Lipid Profiling of Serum Using Optimized Derivatization and Separation Conditions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spener, F.; Lagarde, M.; Géloën, A.; Record, M. What Is Lipidomics? Eur. J. Lipid Sci. Technol. 2003, 105, 481. [Google Scholar] [CrossRef]
- Han, X.; Gross, R.W. Global Analyses of Cellular Lipidomes Directly from Crude Extracts of Biological Samples by ESI Mass Spectrometry: A Bridge to Lipidomics. J. Lipid Res. 2003, 44, 1071–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Li, B.; Lam, S.M.; Shui, G. Integration of Lipidomics and Metabolomics for In-Depth Understanding of Cellular Mechanism and Disease Progression. J. Genet. Genom. 2020, 47, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Holčapek, M.; Liebisch, G.; Ekroos, K. Lipidomic Analysis. Anal. Chem. 2018, 90, 4249–4257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Lees, M.; Stanley, G.H.S. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Ostermann, A.I.; Müller, M.; Willenberg, I.; Schebb, N.H. Determining the Fatty Acid Composition in Plasma and Tissues as Fatty Acid Methyl Esters Using Gas Chromatography—A Comparison of Different Derivatization and Extraction Procedures. Prostaglandins Leukot. Essent Fat. Acids 2014, 91, 235–241. [Google Scholar] [CrossRef]
- Morrison, W.R.; Smith, L.M. Preparation of Fatty Acid Methyl Esters and Dimethylacetals from Lipids with Boron. Fluorid. Methanol. 1964, 5, 600–608. [Google Scholar] [CrossRef]
- Metcalfe, L.D.; Schmitz, A.A.; Pelka, J.R. Rapid Preparation of Fatty Acid Esters from Lipids for Gas Chromatographic Analysis. Anal. Chem. 1966, 38, 514–515. [Google Scholar] [CrossRef]
- Lands, W.E.M.; Libelt, B.; Morris, A.; Kramer, N.C.; Prewitt, T.E.; Bowen, P.; Schmeisser, D.; Davidson, M.H.; Burns, J.H. Maintenance of Lower Proportions of (n − 6) Eicosanoid Precursors in Phospholipids of Human Plasma in Response to Added Dietary (n − 3) Fatty Acids. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 1992, 1180, 147–162. [Google Scholar] [CrossRef]
- Ma, J.; Folsom, A.R.; Shahar, E.; Eckfeldt, J.H. Plasma Fatty Acid Composition as an Indicator of Habitual Dietary Fat Intake in Middle-Aged Adults. The Atherosclerosis Risk in Communities (ARIC) Study Investigators. Am. J. Clin. Nutr. 1995, 62, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Schmidt, S.; Kressel, G.; Willenberg, I.; Hammock, B.D.; Hahn, A.; Schebb, N.H. Modulation of Blood Oxylipin Levels by Long-Chain Omega-3 Fatty Acid Supplementation in Hyper- and Normolipidemic Men. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djuricic, I.; Calder, P.C. Beneficial Outcomes of Omega-6 and Omega-3 Polyunsaturated Fatty Acids on Human Health: An Update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef] [PubMed]
- Micalizzi, G.; Ragosta, E.; Farnetti, S.; Dugo, P.; Tranchida, P.Q.; Mondello, L.; Rigano, F. Rapid and Miniaturized Qualitative and Quantitative Gas Chromatography Profiling of Human Blood Total Fatty Acids. Anal. Bioanal. Chem. 2020, 412, 2327–2337. [Google Scholar] [CrossRef] [PubMed]
- Duarte, D.A.S.; Schroyen, M.; Mota, R.R.; Vanderick, S.; Gengler, N. Recent Genetic Advances on Boar Taint Reduction as an Alternative to Castration: A Review. J. Appl. Genet. 2021, 62, 137–150. [Google Scholar] [CrossRef]
- Fischer, J.; Elsinghorst, P.W.; Bücking, M.; Tholen, E.; Petersen, B.; Wüst, M. Development of a Candidate Reference Method for the Simultaneous Quantitation of the Boar Taint Compounds Androstenone, 3α-Androstenol, 3β-Androstenol, Skatole, and Indole in Pig Fat by Means of Stable Isotope Dilution Analysis-Headspace Solid-Phase Microextraction-Gas Chromatography/Mass Spectrometry. Anal. Chem. 2011, 83, 6785–6791. [Google Scholar] [CrossRef]
- Bekaert, K.M.; vanden Bussche, J.; François, S.; Tuyttens, F.A.M.; de Brabander, H.F.; Vandendriessche, F.; Vanhaecke, L. A Validated Ultra-High Performance Liquid Chromatography Coupled to High Resolution Mass Spectrometry Analysis for the Simultaneous Quantification of the Three Known Boar Taint Compounds. J. Chromatogr. A 2012, 1239, 49–55. [Google Scholar] [CrossRef]
- Buttinger, G.; Wenzl, T. Validation by Collaborative Trial of a Method for the Determination by GC–MS and LC–MS/MS of Boar Taint Marker Compounds in Pork Tissue. Food Chem X 2020, 6, 100083. [Google Scholar] [CrossRef]
- Burgeon, C.; Debliquy, M.; Lahem, D.; Rodriguez, J.; Ly, A.; Fauconnier, M.L. Past, Present, and Future Trends in Boar Taint Detection. Trends Food Sci. Technol. 2021, 112, 283–297. [Google Scholar] [CrossRef]
- Trautmann, J.; Gertheiss, J.; Wicke, M.; Mörlein, D. How Olfactory Acuity Affects the Sensory Assessment of Boar Fat: A Proposal for Quantification. Meat. Sci. 2014, 98, 255–262. [Google Scholar] [CrossRef]
- Sørensen, K.M.; Engelsen, S.B. Measurement of Boar Taint in Porcine Fat Using a High-Throughput Gas Chromatography-Mass Spectrometry Protocol. J. Agric. Food Chem. 2014, 62, 9420–9427. [Google Scholar] [CrossRef] [PubMed]
- Verplanken, K.; Wauters, J.; van Durme, J.; Claus, D.; Vercammen, J.; de Saeger, S.; Vanhaecke, L. Rapid Method for the Simultaneous Detection of Boar Taint Compounds by Means of Solid Phase Microextraction Coupled to Gas Chromatography/Mass Spectrometry. J. Chromatogr. A 2016, 1462, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Hansen-Møller, J. Rapid High-Performance Liquid Chromatographic Method for Simultaneous Determination of Androstenone, Skatole and Indole in Back Fat from Pigs. J. Chromatogr. B Biomed. Sci. Appl. 1994, 661, 219–230. [Google Scholar] [CrossRef]
- Liu, X.; Schmidt, H.; Mörlein, D. Feasibility of Boar Taint Classification Using a Portable Raman Device. Meat. Sci. 2016, 116, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A Web Server for Metabolomic Data Analysis and Interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugan, M.E.R.; Vahmani, P.; Turner, T.D.; Mapiye, C.; Juárez, M.; Prieto, N.; Beaulieu, A.D.; Zijlstra, R.T.; Patience, J.F.; Aalhus, J.L. Pork as a Source of Omega-3 (n-3) Fatty Acids. J. Clin. Med. 2015, 4, 1999–2011. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | −1 | 0 | +1 |
|---|---|---|---|
| T1 (°C) | 85 | 95 | 105 |
| t1 (min) | 5 | 15 | 25 |
| T2 (°C) | 85 | 95 | 105 |
| t2 (min) | 5 | 15 | 25 |
| SFA | MUFA | Omega-6 | Omega-3 | Omega-3 | |
|---|---|---|---|---|---|
| Analyte | C12:0 | C14:1 | C18:3 n-6 | C20:4 n-3 | C22:6 n-3 |
| 1tR (min) | 16.39 | 23.46 | 39.46 | 48.26 | 55.06 |
| 2tR (s) | 4.07 | 5.08 | 6.36 | 6.986 | 7.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhatt, K.; Dejong, T.; Dubois, L.M.; Markey, A.; Gengler, N.; Wavreille, J.; Stefanuto, P.-H.; Focant, J.-F. Lipid Serum Profiling of Boar-Tainted and Untainted Pigs Using GC×GC–TOFMS: An Exploratory Study. Metabolites 2022, 12, 1111. https://doi.org/10.3390/metabo12111111
Bhatt K, Dejong T, Dubois LM, Markey A, Gengler N, Wavreille J, Stefanuto P-H, Focant J-F. Lipid Serum Profiling of Boar-Tainted and Untainted Pigs Using GC×GC–TOFMS: An Exploratory Study. Metabolites. 2022; 12(11):1111. https://doi.org/10.3390/metabo12111111
Chicago/Turabian StyleBhatt, Kinjal, Thibaut Dejong, Lena M. Dubois, Alice Markey, Nicolas Gengler, José Wavreille, Pierre-Hugues Stefanuto, and Jean-François Focant. 2022. "Lipid Serum Profiling of Boar-Tainted and Untainted Pigs Using GC×GC–TOFMS: An Exploratory Study" Metabolites 12, no. 11: 1111. https://doi.org/10.3390/metabo12111111