


Effects of Plant-Derived Glycerol Monolaurate (GML) Additive on the Antioxidant Capacity, Anti-Inflammatory Ability, Muscle Nutritional Value, and Intestinal Flora of Hybrid Grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂)
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Diets Preparation
2.2. Experimental Animals and Breeding Management
2.3. Sample Collection
2.4. Liver Antioxidant Enzyme Activity Analysis
2.5. Gene Expression Analysis
2.6. Vibrio parahaemolyticus Challenge Test
2.7. Statistical Analysis
3. Results
3.1. Effects of Dietary GML Levels on the Antioxidant Index in Liver and Serum for Grouper
3.2. Effects of Dietary GML Levels on the Antioxidant Index in Liver and Serum for Grouper
3.3. Effects of Dietary GML Levels on Muscle Fatty Acid Composition for Grouper
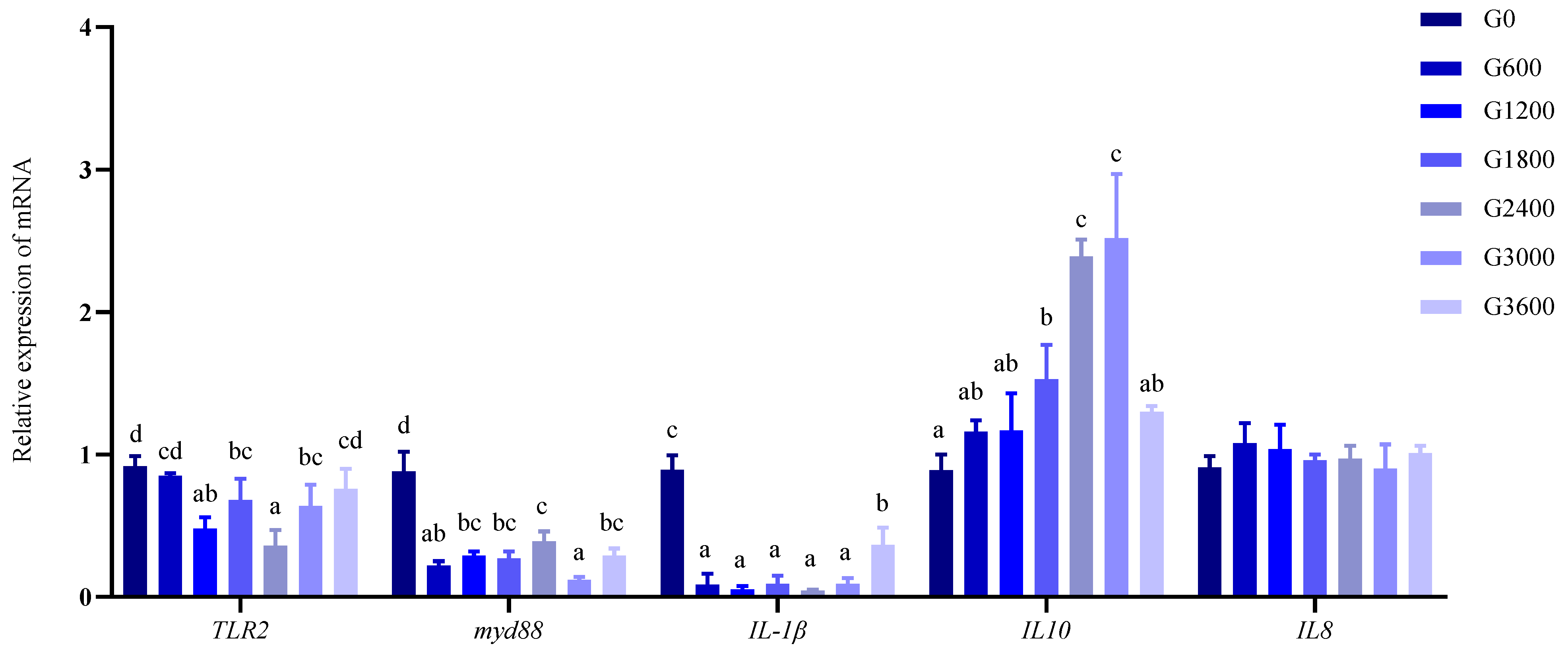
3.4. Effects of Dietary GML Levels on the Relative Expression of myd88 and IL10 in Head Kidney for Grouper
3.5. Vibrio parahaemolyticus Challenge Test Results
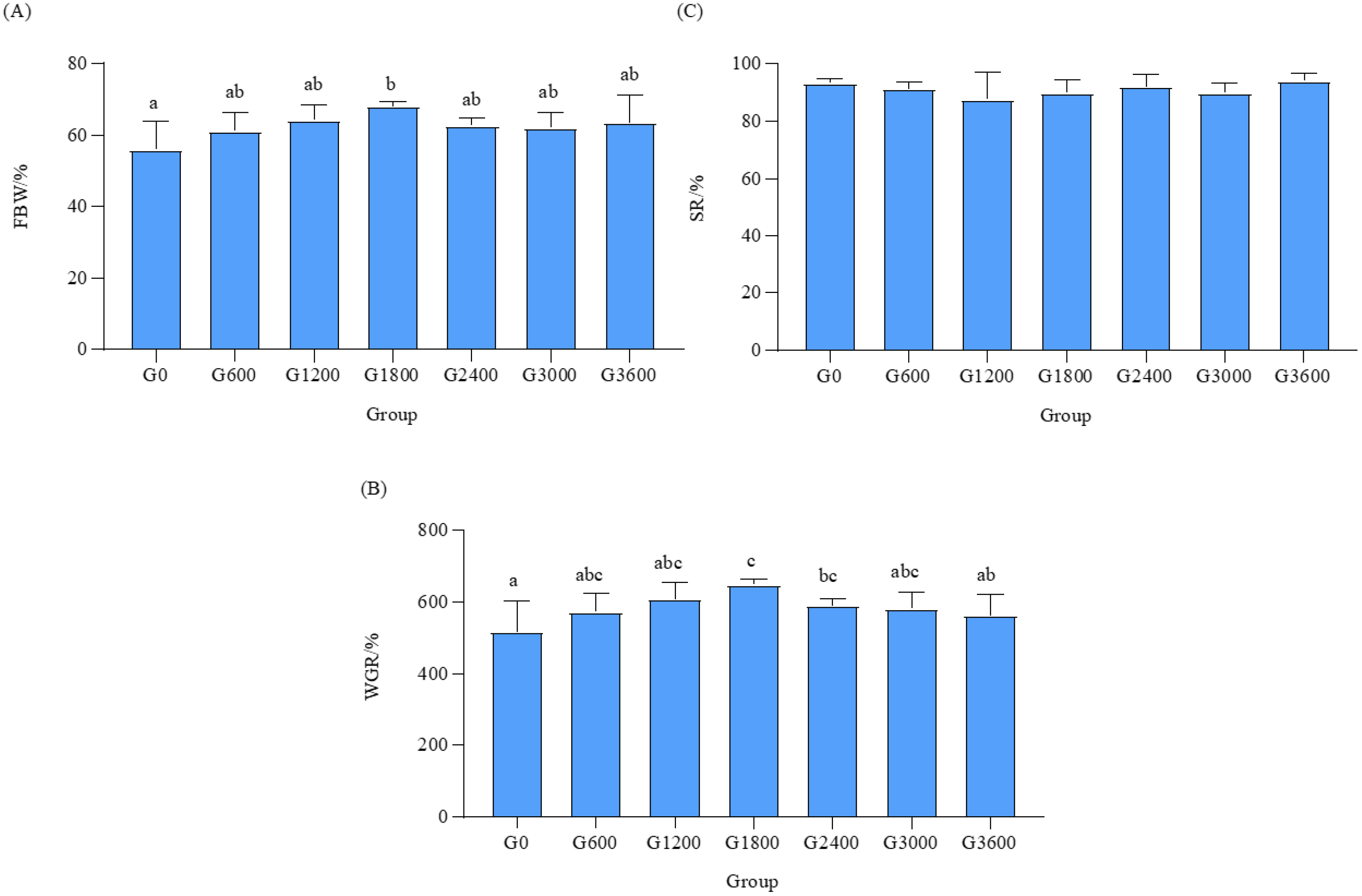
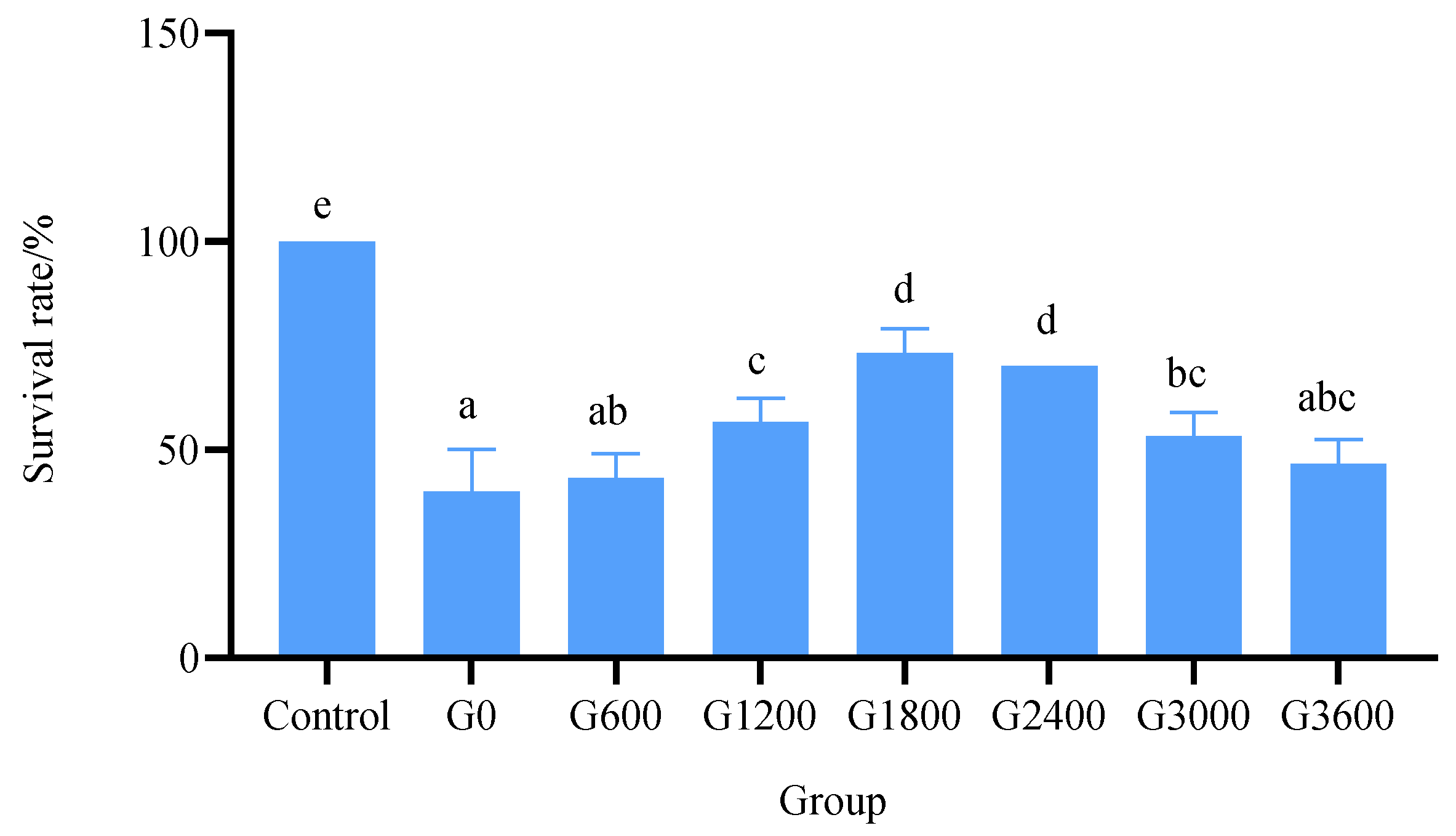
3.5.1. Effects of Dietary GML Levels on Survival Rate in Vibrio parahaemolyticus Challenge Test
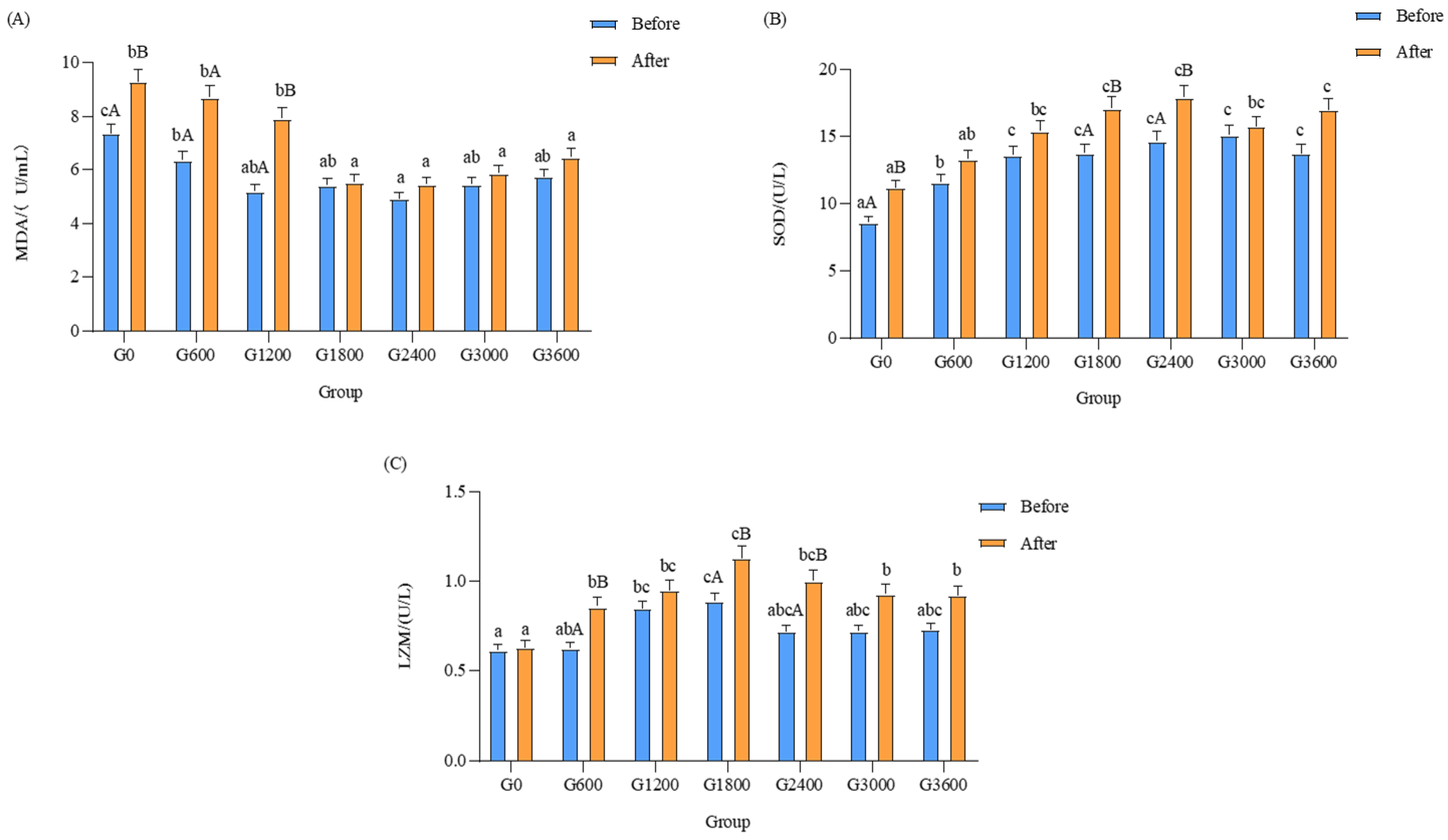
3.5.2. Effects of Dietary GML Levels on Enzyme Activity of LZM in Bacterial Challenge Test
3.5.3. Effects of Dietary GML Levels on Enzyme Activity of SOD in Vibrio parahaemolyticus Challenge Test
3.5.4. Effects of Dietary GML Levels on MDA Content in Vibrio parahaemolyticus Challenge Test
3.6. Effects of Dietary GML Levels on Intestinal Flora for Hybrid Grouper
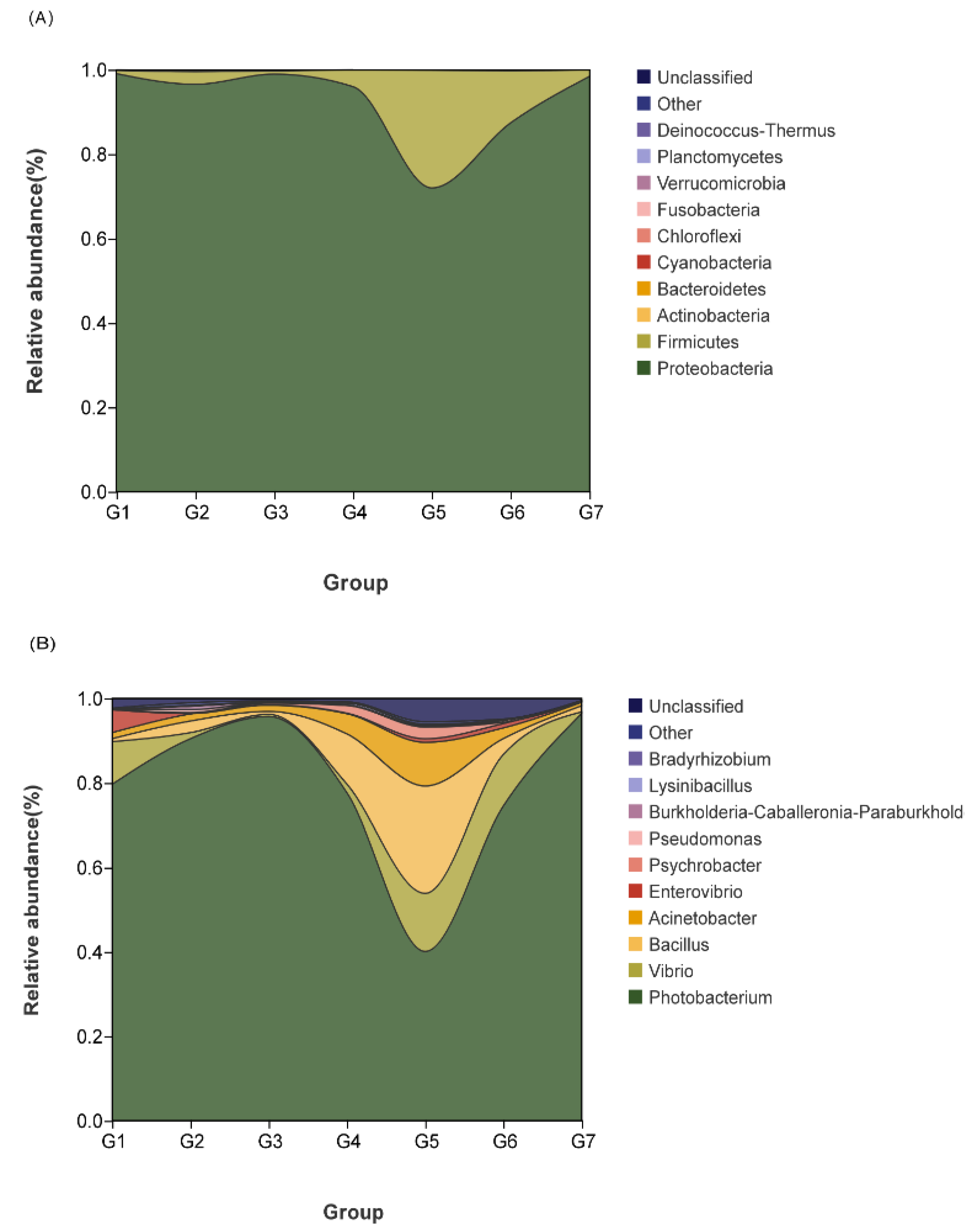
3.6.1. Effects of Dietary GML Levels on Relative Abundance of Species for Hybrid Grouper
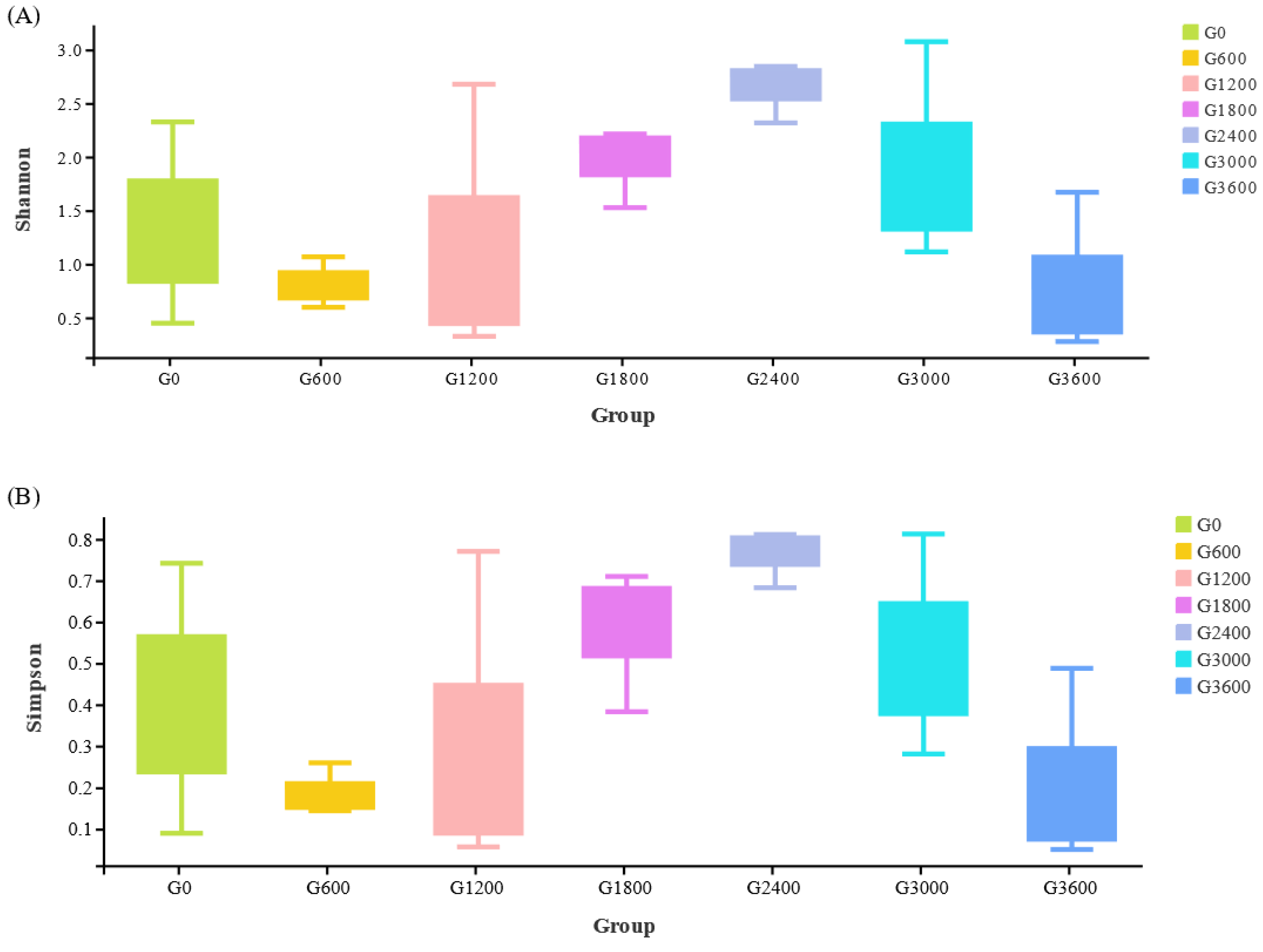
3.6.2. Effects of Dietary GML Levels on Shannon and Simpson Indices for Juvenile Grouper
3.6.3. Analysis of the Association between GML and Intestinal Flora
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tuerxun, M.; Muhda, A.; Yin, L. The molecular mechanisms of signal pathway activating effect of E2F-1/NF-kappaB/GSK-3beta on cognitive dysfunction of alzheimer rats. Bioengineered 2021, 12, 10000–10008. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Kwon, Y.W.; Lee, S.C.; Park, S.D.; Lee, J.H. Herbal formula SC-E1 suppresses lipopolysaccharide-stimulated inflammatory responses through activation of Nrf2/HO-1 signaling pathway in RAW 264.7 macrophages. BMC Complement. Altern. Med. 2017, 17, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeon, L.J.; Jin, S.S.; Sik, S.J.; Kang, S.S.; Hwa, C.S. Bone-healing capacity of pcl/plga/duck beak scaffold in critical bone defects in a rabbit model. Biomed Res. Int. 2016, 2016, 2136215. [Google Scholar]
- Hjelmgren, O.; Gummesson, A.; Bergstrm, G.; Schmidt, C. Beta-cell function, self-rated health, and lifestyle habits in 64-year-old swedish women with metabolically healthy obesity phenotype. J. Obes. Metab. Syndr. 2020, 29, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Tang, Z.; Nan, X.; Sun, F.; Xiong, B. Effects of Lonicera japonica extract on performance, blood biomarkers of inflammation and oxidative stress during perinatal period in dairy cows. Asian-Australas. J. Anim. Sci. 2019, 33, 1096. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Li, D.; Guan, X.; Yang, Y.; Yan, J.; Shi, J.; Ma, R.; Shu, Q. MsrA suppresses inflammatory activation of microglia and oxidative stress to prevent demyelination via inhibition of the NOX2-MAPKs/NF-kappaB signaling pathway. Drug Des. Dev. Ther. 2020, 14, 1377–1389. [Google Scholar] [CrossRef] [Green Version]
- Federico, A.; Morgillo, F.; Tuccillo, C.; Ciardiello, F.; Loguercio, C. Chronic inflammation and oxidative stress in human carcinogenesis. Int. J. Cancer 2010, 121, 2381–2386. [Google Scholar] [CrossRef]
- Rajkumar, K.; Bhattacharya, A.; David, S.; Balaji, S.H.; Hariharan, R.; Jayakumar, M.; Balaji, N. Socio-demographic study on extent of knowledge, awareness, attitude, and risks of zoonotic diseases among livestock owners in Puducherry region. Vet. World 2016, 9, 1018–1024. [Google Scholar] [CrossRef] [Green Version]
- Mazzoli, A.; Donadio, G.; Lanzilli, M.; Saggese, A.; Isticato, R. Bacillus megaterium SF185 spores exert protective effects against oxidative stress in vivo and in vitro. Sci. Rep. 2019, 9, 12082. [Google Scholar] [CrossRef] [Green Version]
- Sadat, A.; Shata, R.R.; Farag, A.M.; Ramadan, H.; Alkhedaide, A.; Soliman, M.M.; Elbadawy, M.; Abugomaa, A.; Awad, A. Prevalence and characterization of PVL-positive staphylococcus aureus isolated from raw cow’s milk. Toxins 2022, 14, 97. [Google Scholar] [CrossRef]
- Lindberg, J.E. Fiber effects in nutrition and gut health in pigs. J. Anim. Sci. Biotechnol. 2014, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Kurjogi, M.; Mohammad, Y.; Alghamdi, S.; Abdelrahman, M.; Jogaiah, S. Detection and determination of stability of the antibiotic residues in cow’s milk. PLoS ONE 2019, 14, e0223475. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.W.; Li, X.X.; Zhang, L.Y.; Wu, J.P.; Zhao, S.G.; Jiao, T. Effect of oregano oil and cobalt lactate on sheep in vitro digestibility, fermentation characteristics and rumen microbial community. Animals 2022, 12, 118. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Wu, H.; Wang, X.; He, J.; Yin, Y. Resveratrol attenuates oxidative stress-induced intestinal barrier injury through PI3K/Akt-Mediated Nrf2 signaling pathway. Oxid. Med. Cell. Longev. 2019, 2019, 7591840. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Zhao, M.; Zhang, H.; Li, Y.; Liu, M.; Feng, F. Antimicrobial emulsifier-glycerol monolaurate induces metabolic syndrome, gut microbiota dysbiosis and systemic low-grade inflammation in low-fat diet fed mice. Mol. Nutr. Food Res. 2018, 62, 1700547. [Google Scholar] [CrossRef]
- Schlievert, P.M.; Kilgore, S.H.; Seo, K.S.; Leung, D. Glycerol monolaurate contributes to the antimicrobial and anti-inflammatory activity of human milk. Sci. Rep. 2019, 9, 14550. [Google Scholar] [CrossRef] [Green Version]
- Welch, J.L.; Xiang, J.; Okeoma, C.M.; Schlievert, P.M.; Stapleton, J.T. Glycerol monolaurate, an analogue to a factor secreted by Lactobacillus, is virucidal against enveloped viruses, including HIV-1. mBio 2020, 11, e00686-20. [Google Scholar] [CrossRef]
- Seleem, D.; Chen, E.; Benso, B.; Pardi, V.; Murata, R.M. In vitro evaluation of antifungal activity of monolaurin against Candida albicans biofilms. Peer J. 2016, 4, e2148. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wei, H.; Cui, Y.; Zhao, G.; Feng, F. Antibacterial interactions of monolaurin with commonly used antimicrobials and food components. J. Food Sci. 2009, 74, 418–427. [Google Scholar] [CrossRef]
- Zhu, Z.K.; Tan, Z.H.; Wang, S.J.; Ning, N.; Wang, M.L.; Wang, L. Effects of glycerol monolaurate on antioxidant function and lipid metabolism gene expression in liver of piglets infected with porcine epidemic diarrhea virus. Chin. J. Anim. Sci. 2022, 58, 209–213. [Google Scholar]
- Kong, L.; Wang, Z.; Xiao, C.; Zhu, Q.; Song, Z. Glycerol monolaurate ameliorated intestinal barrier and immunity in broilers by regulating intestinal inflammation, antioxidant balance, and intestinal microbiota. Front. Immunol. 2021, 12, 3913. [Google Scholar] [CrossRef] [PubMed]
- Valentini, J.; Silva, A.; Fortuoso, B.F.; Reis, J.H.; Tavernari, F.C. Chemical composition, lipid peroxidation, and fatty acid profile in meat of broilers fed with glycerol monolaurate additive. Food Chem. 2020, 330, 127187. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Cai, H.; Liu, M.; Deng, L.; Li, Y.; Zhang, H.; Feng, F. Effects of dietary glycerol monolaurate on productive performance, egg quality, serum biochemical indices, and intestinal morphology of laying hens. J. Zhejiang Univ. Sci. B 2019, 20, 877. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Z.; Li, X.L.; Ma, T.; Ding, H.B. Effects of monoglyceride laurate supplementation on growthperformance serum biochemical indexes and antioxidant capacity of weaned lambs. Chin. J. Anim. Nutr. 2021, 33, 6593–6600. [Google Scholar]
- Wang, Y.; Zhang, C.; Li, Y.; Zhang, H.; Wang, J.; Feng, F. Effects of dietary glycerol monolaurate on the growth performance, digestive enzymes, body composition and non-specific immune response of white shrimp (Litopenaeus vannamei). Aquacult. Rep. 2020, 18, 100535. [Google Scholar] [CrossRef]
- Wang, Y.; Zhong, H.; Wang, J.; Feng, F. Dietary glycerol monolaurate improved the growth, activity of digestive enzymes and gut microbiota in zebrafish (Danio rerio). Aquacult. Rep. 2021, 20, 100670. [Google Scholar] [CrossRef]
- Wang, Y.C.; Du, J.; Li, Y.; Zhang, H.; Feng, F.Q. Effects of glycerol monolaurate on growth, health and nutritional quality ofchinese soft-shelled turtle (Pelodiscus sinensis). Chin. J. Anim. Nutr. 2019, 31, 428–436. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1995; Volume 2. [Google Scholar]
- Ray, G.W.; Liang, D.; Yang, Q.; Tan, B.; Li, R. Effects of replacing fishmeal with dietary soybean protein concentrate (SPC) on growth, serum biochemical indices, and antioxidative functions for juvenile shrimp Litopenaeus vannamei. Aquaculture 2019, 516, 734630. [Google Scholar] [CrossRef]
- Lin, H.; Ding, M.; Ray, G.W.; Tan, B.; Yang, Q. Vitamin D promotes growth, feed utilization and gene expression related to lipid metabolism for juvenile orange-spotted grouper Epinephelus coioides. Aquacult. Rep. 2022, 23, 101074. [Google Scholar] [CrossRef]
- Yang, Q.; Ding, M.; Tan, B.; Dong, X.; Chi, S.; Shuang, Z.; Liu, H. Effects of dietary vitamin A on growth, feed utilization, lipid metabolism enzyme activities, and fatty acid synthase and hepatic lipase mRNA expression levels in the liver of juvenile orange spotted grouper, Epinephelus coioides. Aquaculture 2017, 479, 501–507. [Google Scholar] [CrossRef]
- He, Y.; Guo, X.; Tan, B.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S.; Chi, S. Partial fishmeal protein replacement with peptides from swine blood modulates the nutritional status, immune response, and intestinal microbiota of hybrid groupers (female Epinephelus fuscoguttatus × male E. lanceolatus). Aquaculture 2021, 533, 736154. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [PubMed]
- Welker, A.F.; Campos, E.G.; Cardoso, L.A.; Hermes-Lima, M. Role of catalase on the hypoxia/reoxygenation stress in the hypoxia-tolerant Nile tilapia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R1111. [Google Scholar] [PubMed] [Green Version]
- Kong, S.Z.; Li, J.C.; Li, S.D.; Liao, M.N.; Li, C.P.; Zheng, P.J.; Guo, M.H.; Tan, W.X.; Zheng, Z.H.; Zhang, H. Anti-Aging Effect of chitosan oligosaccharide on d-galactose-induced subacute aging in mice. Mar. Drugs 2018, 16, 181. [Google Scholar]
- Qian, X.; Zhu, F. Use of glycerol monolaurate as a treatment against white spot syndrome virus in crayfish (Procambarus clarkii). Aquaculture 2021, 541, 736853. [Google Scholar]
- Liu, M.; Chen, X.; Zhao, H.; Feng, F. Effect of dietary supplementation with glycerol monolaurate on growth performance, digestive ability and chicken nutritional components of broilers. Food Sci. 2018, 39, 67–71. [Google Scholar]
- Sun, C.Y.; Dong, H.B.; Wang, W.H.; Li, Y.; Gu, Q.H.; Duan, Y.F.; Zhang, G.S.; Xu, X.D. Effects of glycerol monolaurate on lipid metabolism of Lateolabrax maculatus. South China Fish. Sci. 2021, 17, 67–75. [Google Scholar]
- Famurewa, A.C.; Aja, P.M.; Maduagwuna, E.K.; Ekeleme-Egedigwe, C.A.; Ufebe, O.G.; Azubuike-Osu, S.O. Antioxidant and anti-inflammatory effects of virgin coconut oil supplementation abrogate acute chemotherapy oxidative nephrotoxicity induced by anticancer drug methotrexate in rats. Biomed. Pharm. 2017, 96, 905–911. [Google Scholar]
- Jack, O. New insights into the utilization of medium-chain triglycerides by the neonate: Observations from a piglet model. J. Nutr. 1997, 127, 1061–1067. [Google Scholar]
- An, Y.; Cui, B.; Wang, Y.; Jin, W.; Geng, X.; Yan, X.; Li, B. Functional properties of ovalbumin glycosylated with carboxymethyl cellulose of different substitution degree. Food Hydrocoll. 2014, 40, 1–8. [Google Scholar]
- Zhang, T.; Niu, Q.; Wang, T.; Zheng, X.; Li, H.; Gao, X.; Chen, Y.; Gao, H.; Zhang, L.; Liu, G.E. Comparative transcriptomic analysis reveals diverse expression pattern underlying fatty acid composition among different beef cuts. Foods 2022, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.Q. The Effect of Glycerol Monolaurate on Growth, Health and Food Quality of Cultured Large Yellow Croaker. Master’s Thesis, Zhejiang University, Hangzhou, China, 2021. [Google Scholar]
- Kim, K.S.; Oh, D.H.; Kim, J.Y.; Lee, B.G.; You, J.S.; Chang, K.J.; Chung, H.; Yoo, M.C.; Yang, H.-I.; Kang, J.-H. Taurine ameliorates hyperglycemia and dyslipidemia by reducing insulin resistance and leptin level in otsukal ong-evans tokushima fatty (OLETF) rats with long-term diabetes. Exp. Mol. Med. 2012, 44, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Tyra, M. The effect of replacement of soybean meal with corn dried distillers grains with solubles (cddgs) and differentiation of dietary fat sources on pig meat quality and fatty acid profile. Animals 2021, 11, 1277. [Google Scholar]
- Cui, Z.; Wang, X.; Hou, Z.; Liao, S.; Tan, B. Low-protein diet supplemented with medium-chain fatty acid glycerides improves the growth performance and intestinal function in post-weaning piglets. Animals 2020, 10, 1852. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.C. Personalised medicine in child and Adolescent Psychiatry: Focus on omega-3 polyunsaturated fatty acids and ADHD. Brain Behav. Immun.-Health 2021, 16, 100310. [Google Scholar] [CrossRef]
- Huang, C.B.; George, B.; Ebersole, J.L. Antimicrobial activity of n-6, n-7 and n-9 fatty acids and their esters for oral microorganisms. Arch. Oral Biol. 2010, 55, 555–560. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ning, X.; He, X.; Sun, X.; Wu, Y. Fatty acid composition analyses of commercially important fish species from the Pearl River Estuary, China. PLoS ONE 2020, 15, e0228276. [Google Scholar] [CrossRef] [Green Version]
- Halade, G.V.; Norris, P.C.; Kain, V.; Serhan, C.N.; Ingle, K.A. Splenic leukocytes define the resolution of inflammation in heart failure. Sci. Signal. 2018, 11, eaao1818. [Google Scholar] [CrossRef] [Green Version]
- Schlievert, P.M.; Peterson, M.L. Glycerol monolaurate antibacterial activity in broth and biofilm cultures. PLoS ONE 2012, 7, e40350. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Huang, W.; Chen, Y.; Du, Z.; Zhu, F.; Wang, T.; Jiang, B. Ischemic postconditioning ameliorates acute kidney injury induced by limb ischemia/reperfusion via transforming TLR4 and NF-kappaB signaling in rats. J. Orthop. Surg. Res. 2021, 16, 416. [Google Scholar] [CrossRef]
- Martinez, J.; Huang, X.; Yang, Y. Direct TLR2 signaling is critical for NK cell activation and function in response to vaccinia viral infection. PLoS Pathog. 2010, 6, e1000811. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wu, L.; Deng, H.; Chen, Y.; Zhou, H.; Liu, M.; Wang, S.; Zheng, L.; Zhu, L.; Lv, X. Anti-inflammatory protein TSG-6 secreted by bone marrow mesenchymal stem cells attenuates neuropathic pain by inhibiting the TLR2/MyD88/NF-kappaB signaling pathway in spinal microglia. J. Neuroinflamm. 2020, 17, 154. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Jiang, Z.; Cai, H.; Li, Y.; Feng, F. Modulation of the gut microbiota during high-dose glycerol monolaurate-mediated amelioration of obesity in mice fed a high-fat diet. mBio 2020, 11, e00190-20. [Google Scholar] [CrossRef]
- Wei, H.; Li, R.; Yang, Q.; Tan, B.; Ray, G.W.; Dong, X.; Chi, S.; Liu, H.; Zhang, S. Effects of Zn on growth performance, immune enzyme activities, resistance to disease and intestinal flora for juvenile pearl gentian grouper (Epinephelus lanceolatus♂ × Epinephelus fuscoguttatus♀) under low fishmeal diet. Aquacult. Rep. 2021, 21, 100880. [Google Scholar] [CrossRef]
- Bao, J.; Li, X.; Xing, Y.; Feng, C.; Jiang, H. Effects of hypoxia on immune responses and carbohydrate metabolism in the Chinese mitten crab, Eriocheir sinensis. Aquacult. Res. 2020, 51, 2735–2744. [Google Scholar] [CrossRef]
- Zhu, L.Y.; Nie, L.; Zhu, G.; Xiang, L.X.; Shao, J.Z. Advances in research of fish immune-relevant genes: A comparative overview of innate and adaptive immunity in teleosts. Dev. Comp. Immunol. 2013, 39, 39–62. [Google Scholar] [CrossRef] [PubMed]
- Bergsson, G.; Steingrímsson, Ó.; Thormar, H. Bactericidal effects of fatty acids and monoglycerides on Helicobacter pylori. Int. J. Antimicrob. Agents 2002, 20, 258–262. [Google Scholar] [CrossRef]
- Peterson, M.L.; Schlievert, P.M. Glycerol monolaurate inhibits the effects of Gram-positive select agents on eukaryotic cells. Biochemistry 2006, 45, 2387–2397. [Google Scholar] [CrossRef] [Green Version]
- Schlievert, P.M.; Strandberg, K.L.; Brosnahan, A.J.; Peterson, M.L.; Pambuccian, S.E.; Nephew, K.R.; Brunner, K.G.; Schultz-Darken, N.J.; Haase, A.T. Glycerol monolaurate does not alter rhesus macaque (Macaca mulatta) vaginal lactobacilli and is safe for chronic use. Antimicrob. Agents Chemother. 2008, 52, 4448–4454. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Zeng, Y.; Zhang, H.; Ma, Y. The role of gastrointestinal microbiota in functional dyspepsia: A review. Front. Physiol. 2022, 13, 910568. [Google Scholar] [CrossRef]
- Kolba, N.; Zarei, A.; Cheng, J.; Agarwal, N.; Dadmohammadi, Y.; Khazdooz, L.; Abbaspourrad, A.; Tako, E. Alterations in Intestinal Brush Border Membrane Functionality and Bacterial Populations Following Intra-Amniotic Administration (Gallus gallus) of Nicotinamide Riboside and Its Derivatives. Nutrients 2022, 14, 3130. [Google Scholar] [CrossRef] [PubMed]
- Alkushi, A.; Abdelfattah-Hassan, A.; Eldoumani, H.; Elazab, S.T.; Mohamed, S.A.; Metwally, A.S.; SEl-Shetry, E.; Saleh, A.A.; ElSawy, N.A.; Ibrahim, D. Probiotics-loaded nanoparticles attenuated colon inflammation, oxidative stress, and apoptosis in colitis. Sci. Rep. 2022, 12, 5116. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Xu, S.; Zhang, J.; Hussain, R.; Lu, H.; Ye, Y.; Mehmood, K.; Zhang, H.; Shang, P. First report of fecal microflora of wild bar-headed goose in Tibet Plateau. Front. Vet. Sci. 2021, 8, 791461. [Google Scholar] [CrossRef] [PubMed]
- Ilinskaya, O.N.; Ulyanova, V.V.; Yarullina, D.R.; Gataullin, I.G. Secretome of intestinal Bacilli: A natural guard against pathologies. Front. Microbiol. 2017, 8, 1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar]
- Cha, J.H.; Rahimnejad, S.; Yang, S.Y.; Kim, K.W.; Lee, K.J. Evaluations of Bacillus spp. as dietary additives on growth performance, innate immunity and disease resistance of olive flounder (Paralichthys olivaceus) against Streptococcus iniae and as water additives. Aquaculture 2013, 402–403, 50–57. [Google Scholar] [CrossRef]
- Aly, S.M.; Abdel-Galil Ahmed, Y.; Abdel-Aziz Ghareeb, A.; Mohamed, M.F. Studies on Bacillus subtilis and Lactobacillus acidophilus, as potential probiotics, on the immune response and resistance of Tilapia nilotica (Oreochromis niloticus) to challenge infections. Fish Shellfish Immunol. 2008, 25, 128–136. [Google Scholar] [CrossRef]
- Tseng, D.Y.; Ho, P.L.; Huang, S.Y.; Cheng, S.C.; Shiu, Y.L.; Chiu, C.S.; Liu, C.H. Enhancement of immunity and disease resistance in the white shrimp, Litopenaeus vannamei, by the probiotic, Bacillus subtilis E20. Fish Shellfish Immunol. 2009, 26, 339–344. [Google Scholar] [CrossRef]
- Yu, J.F.; Xia, B.H.; Hao, Q.R.; Zou, H.B.; Wang, P.; Chen, Z.X.; Gao, L.; Wu, S.; Huang, L.; Qin, D.L.; et al. Screening, identification and culture medium optimization of a hypoglycemic Lysinibacillus fusiformis strain from common carp. J. Dalian Ocean Univ. 2022, 37, 611–619. [Google Scholar]
- Guan, L.P.; Xie, J.J.; Li, W.H.; Chen, H.L.; Wen, Z.Z.; Sun, C.B. Effects of a Lysinibacillus strain on the growth and water quality of Litopenaeus Vannamei. Fish. Mod. 2022, 49, 1–7. [Google Scholar]
- Saratale, R.G.; Cho, S.K.; Saratale, G.D.; Ghodake, G.S.; Bharagava, R.N.; Kim, D.S.; Nair, S.; Shin, H.S. Efficient bioconversion of sugarcane bagasse into polyhydroxybutyrate (PHB) by Lysinibacillus sp. and its characterization. Bioresour. Technol. 2021, 324, 124673. [Google Scholar] [CrossRef] [PubMed]
- Asif, T.; Javed, U.; Zafar, S.B.; Ansari, A.; Ul Qader, S.A.; Aman, A. Bioconversion of colloidal chitin using novel chitinase from glutamicibacter uratoxydans exhibiting anti-fungal potential by hydrolyzing chitin within fungal cell wall. Waste Biomass Valorization 2019, 11, 4129–4143. [Google Scholar] [CrossRef]
- Breuil, C.; Kushner, D.J. Lipase and esterase formation by psychrophilic and mesophilic Acinetobacter species. Can. J. Microbiol. 1975, 21, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Park, Y.H.; Lee, J.S.; Jeong, H.I.; Lee, K.W.; Kang, T.H. Anti-obesity effect of DKB-117 through the inhibition of pancreatic lipase and alpha-amylase activity. Nutrients 2020, 12, 3053. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | |||||||
|---|---|---|---|---|---|---|---|
| G0 | G600 | G1200 | G1800 | G2400 | G3000 | G3600 | |
| Brown fish meal | 30.00 | 30.00 | 30.00 | 30.00 | 30.00 | 30.00 | 30.00 |
| American soybean meal | 18.00 | 18.00 | 18.00 | 18.00 | 18.00 | 18.00 | 18.00 |
| Corn gluten meal | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 |
| Peanut meal | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Chicken powder | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 |
| Wheat flour | 18.59 | 18.59 | 18.59 | 18.59 | 18.59 | 18.59 | 18.59 |
| fish oil | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Soybean oil | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Soybean lecithin | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Ca(H2PO4)2 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Premix | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Choline chloride | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| vitamin C | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Microcrystalline cellulose | 0.36 | 0.30 | 0.24 | 0.18 | 0.12 | 0.06 | 0.00 |
| GML | 0.00 | 0.06 | 0.12 | 0.18 | 0.24 | 0.30 | 0.36 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| Nutrient levels | |||||||
| Crude protein | 49.74 | 49.19 | 49.65 | 49.66 | 49.19 | 49.32 | 49.40 |
| Crude lipid | 9.42 | 9.46 | 9.38 | 9.37 | 9.28 | 9.29 | 9.18 |
| Nucleotide Sequence (5′–3′) | Genbank Accession No. | |
|---|---|---|
| β-actin | F: GATCTGGCATCACACCTTCT R: CATCTTCTCCCTGTTGGCTT | AY510710.2 |
| TLR2 | F: AGGGTTCAGAAGGGTTGCTAT R: CAGGAAGGAAGTCCCGTTTGT | HM357230.1 |
| myd88 | F: AGCTGGAGCAGACGGAGTG R: GAGGCTGAGAGCAAACTTGGTC | JF271883.1 |
| IL-1β | F: AACCTCATCATCGCCACACA R: AGTTGCCTCACAACCGAACAC | XP_049460451.1 |
| IL-10 | F: ACACAGCGCTGCTAGACGAG R: GGGCAGCACCGTGTTCAGAT | KJ741852.1 |
| IL8 | F: GGCCGTCAGTGAAGGGAGTC R: TCAGAGTGGCAATGATCTCA | GU988706.1 |
| G0 | G600 | G1200 | G1800 | G2400 | G3000 | G3600 | |
|---|---|---|---|---|---|---|---|
| Liver | |||||||
| MDA/nmol/mL | 1.57 ± 0.26 d | 1.27 ± 0.21 bc | 1.08 ± 0.02 b | 0.86 ± 0.11 a | 1.28 ± 0.01 bc | 1.40 ± 0.01 cd | 1.14 ± 0.13 c |
| SOD/U/mL | 16.91 ± 0.46 a | 17.31 ± 0.31 a | 22.04 ± 0.42 c | 26.01 ± 0.73 d | 23.54 ± 1.14 cd | 21.38 ± 4.01 b | 18.64 ± 1.57 ab |
| CAT/U/mL | 8.29 ± 1.15 | 9.37 ± 0.11 | 10.19 ± 1.49 | 9.64 ± 0.79 | 9.31 ± 0.51 | 9.51 ± 0.97 | 8.84 ± 0.50 |
| GSH-Px/U/mL | 94.56 ± 12.43 a | 94.90 ± 13.80 a | 96.16 ± 5.01 ab | 113.53 ± 3.60 c | 108.86 ± 7.95 c | 106.96 ± 7.04 bc | 97.64 ± 8.87 ab |
| Serum | |||||||
| MDA/nmol/mL | 1.66 ± 0.13 e | 1.73 ± 0.07 e | 1.12 ± 0.17 c | 0.79 ± 0.07 a | 1.00 ± 0.11 bc | 0.91 ± 0.11 ab | 1.36 ± 0.07 d |
| SOD/U/mL | 8.61 ± 0.56 a | 13.33 ± 1.94 b | 13.62 ± 2.12 b | 17.14 ± 0.67 c | 17.95 ± 1.71 c | 15.74 ± 2.34 bc | 17.01 ± 3.03 c |
| G0 | G600 | G1200 | G1800 | G2400 | G3000 | G3600 | |
|---|---|---|---|---|---|---|---|
| C12:0 | 0.00 ± 0.00 a | 0.27 ± 0.01 b | 0.61 ± 0.01 c | 0.75 ± 0.04 d | 0.83 ± 0.01 e | 1.25 ± 0.12 f | 1.37 ± 0.03 g |
| C14:0 | 2.56 ± 0.07 a | 2.77 ± 0.00 bc | 2.68 ± 0.08 ab | 2.70 ± 0.25 ab | 2.94 ± 0.01 c | 2.94 ± 0.01 c | 2.78 ± 0.02 bc |
| C16:0 | 19.87 ± 0.16 b | 20.41 ± 0.09 c | 19.73 ± 0.08 b | 19.11 ± 0.05 a | 20.48 ± 0.32 c | 20.36 ± 0.21 c | 19.81 ± 0.07 b |
| C16:1n7 | 2.85 ± 0.03 a | 3.41 ± 0.02 f | 3.04 ± 0.02 c | 2.95 ± 0.03 b | 3.29 ± 0.02 e | 3.24 ± 0.02 d | 3.03 ± 0.03 c |
| C17:0 | 0.35 ± 0.01 b | 0.35 ± 0.01 b | 0.39 ± 0.01 c | 0.41 ± 0.01 c | 0.40 ± 0.01 c | 0.41 ± 0.02 c | 0.29 ± 0.03 a |
| C18:0 | 8.18 ± 0.13 f | 6.89 ± 0.03 a | 7.77 ± 0.09 e | 7.45 ± 0.03 c | 7.25 ± 0.04 b | 7.74 ± 0.04 e | 7.62 ± 0.01 d |
| C18:1n9c | 17.84 ± 0.28 a | 19.94 ± 0.14 e | 18.91 ± 0.03 d | 18.63 ± 0.06 bc | 19.11 ± 0.13 d | 18.62 ± 0.26 bc | 18.32 ± 0.06 b |
| C18:2n6c | 24.09 ± 0.08 c | 25.25 ± 0.10 e | 24.98 ± 0.05 d | 25.30 ± 0.08 e | 23.91 ± 0.22 bc | 23.14 ± 0.14 a | 23.79 ± 0.21 b |
| C18:3n3 | 0.00 ± 0.00 a | 3.98 ± 0.14 c | 3.54 ± 0.04 b | 3.55 ± 0.03 b | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| C20:0 | 0.45 ± 0.01 e | 0.31 ± 0.00 abc | 0.33 ± 0.02 bcd | 0.30 ± 0.01 ab | 0.34 ± 0.02 cd | 0.34 ± 0.01 d | 0.29 ± 0.02 a |
| C20:1 | 3.32 ± 0.04 b | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 3.56 ± 0.02 d | 3.50 ± 0.06 d | 3.42 ± 0.07 c |
| C20:2 | 3.32 ± 0.04 b | 0.00 ± 0.00 c | 0.00 ± 0.00 d | 0.00 ± 0.00 e | 3.56 ± 0.02 a | 3.50 ± 0.06 bc | 3.42 ± 0.07 d |
| C20:3n6 | 0.34 ± 0.01 d | 0.23 ± 0.02 b | 0.28 ± 0.03 c | 0.26 ± 0.01 c | 0.28 ± 0.01 c | 0.38 ± 0.01 e | 0.20 ± 0.01 a |
| C20:4n6 | 1.31 ± 0.04 e | 1.02 ± 0.02 ab | 1.04 ± 0.04 abc | 1.09 ± 0.03 e | 0.99 ± 0.06 a | 1.10 ± 0.01 c | 1.17 ± 0.05 d |
| C20:5n3 | 6.12 ± 0.10 d | 5.80 ± 0.02 b | 5.58 ± 0.04 a | 5.96 ± 0.07 bc | 5.90 ± 0.04 bc | 5.89 ± 0.04 bc | 6.11 ± 0.06 d |
| C22:0 | 0.45 ± 0.00 d | 0.30 ± 0.02 b | 0.38 ± 0.03 c | 0.32 ± 0.01 c | 0.31 ± 0.02 b | 0.34 ± 0.02 b | 0.24 ± 0.01 a |
| C22:1n9 | 0.36 ± 0.01 bc | 0.39 ± 0.00 c | 0.35 ± 0.03 b | 0.37 ± 0.03 bc | 0.31 ± 0.01 a | 0.34 ± 0.02 ab | 0.35 ± 0.02 b |
| C22:6n3 | 8.92 ± 0.14 c | 7.50 ± 0.13 a | 8.39 ± 0.05 b | 9.23 ± 0.08 d | 8.86 ± 0.02 c | 8.92 ± 0.14 c | 9.62 ± 0.20 e |
| C24:1n9 | 0.87 ± 0.07 d | 0.53 ± 0.03 a | 1.08 ± 0.03 e | 0.59 ± 0.03 ab | 0.62 ± 0.02 bc | 0.68 ± 0.06 c | 0.66 ± 0.06 bc |
| ∑SFA | 31.86 ± 0.21 b | 31.29 ± 0.08 a | 31.88 ± 0.11 b | 31.04 ± 0.16 a | 32.56 ± 0.31 c | 33.38 ± 0.18 d | 32.41 ± 0.08 c |
| ∑n-3 | 15.04 ± 0.12 b | 17.28 ± 0.02 d | 17.51 ± 0.09 d | 18.74 ± 0.16 e | 14.75 ± 0.05 a | 14.81 ± 0.19 ab | 15.73 ± 0.24 c |
| ∑n-6 | 25.74 ± 0.12 c | 26.49 ± 0.07 de | 26.30 ± 0.08 d | 26.65 ± 0.06 e | 25.19 ± 0.27 b | 24.62 ± 0.13 a | 25.15 ± 0.19 b |
| ∑n-3/∑n-6 | 0.58 ± 0.00 a | 0.65 ± 0.00 d | 0.67 ± 0.00 d | 0.70 ± 0.01 e | 0.59 ± 0.01 a | 0.60 ± 0.01 b | 0.63 ± 0.01 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Yi, Y.; Wu, J.; Yang, Q.; Tan, B.; Chi, S. Effects of Plant-Derived Glycerol Monolaurate (GML) Additive on the Antioxidant Capacity, Anti-Inflammatory Ability, Muscle Nutritional Value, and Intestinal Flora of Hybrid Grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂). Metabolites 2022, 12, 1089. https://doi.org/10.3390/metabo12111089
Li X, Yi Y, Wu J, Yang Q, Tan B, Chi S. Effects of Plant-Derived Glycerol Monolaurate (GML) Additive on the Antioxidant Capacity, Anti-Inflammatory Ability, Muscle Nutritional Value, and Intestinal Flora of Hybrid Grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂). Metabolites. 2022; 12(11):1089. https://doi.org/10.3390/metabo12111089
Chicago/Turabian StyleLi, Xuehe, Yuanming Yi, Jiahua Wu, Qihui Yang, Beiping Tan, and Shuyan Chi. 2022. "Effects of Plant-Derived Glycerol Monolaurate (GML) Additive on the Antioxidant Capacity, Anti-Inflammatory Ability, Muscle Nutritional Value, and Intestinal Flora of Hybrid Grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂)" Metabolites 12, no. 11: 1089. https://doi.org/10.3390/metabo12111089