
Human Milk from Tandem Feeding Dyads Does Not Differ in Metabolite and Metataxonomic Features When Compared to Single Nursling Dyads under Six Months of Age
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participant Recruitment
2.2. Sample Collection, Handling, and Processing
2.3. DNA Extraction and Metataxonomic Analysis
2.4. Metabolic Fingerprinting Using Laser Assisted Rapid Evaporative Ionisation Mass Spectrometry (LA-REIMS)
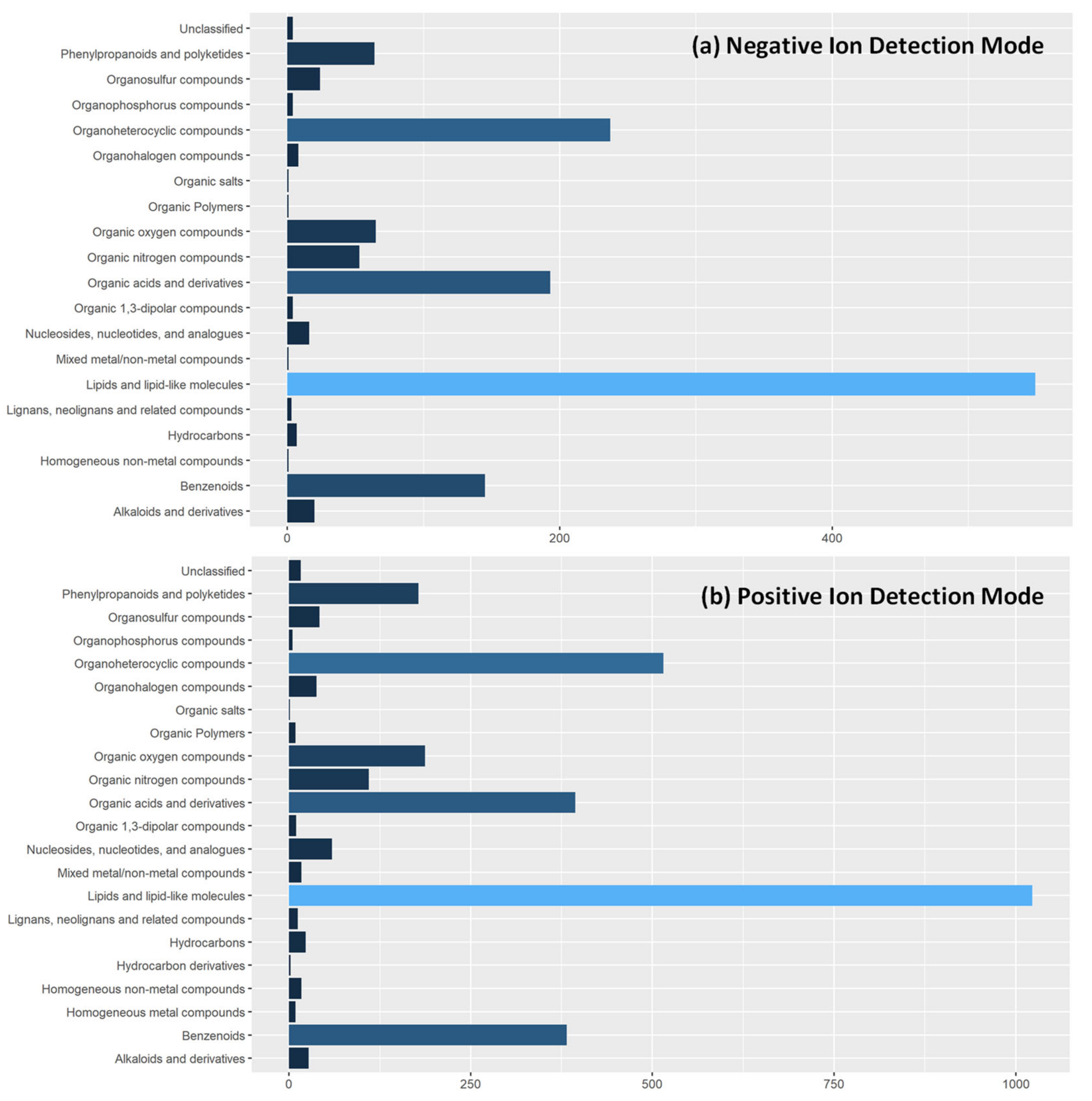
2.5. Processing and Tentative Identifications of LA-REIMS Metabolic Fingerprinting Data
2.6. Statistical Analysis
2.7. Availability of Raw Data
3. Results and Discussion
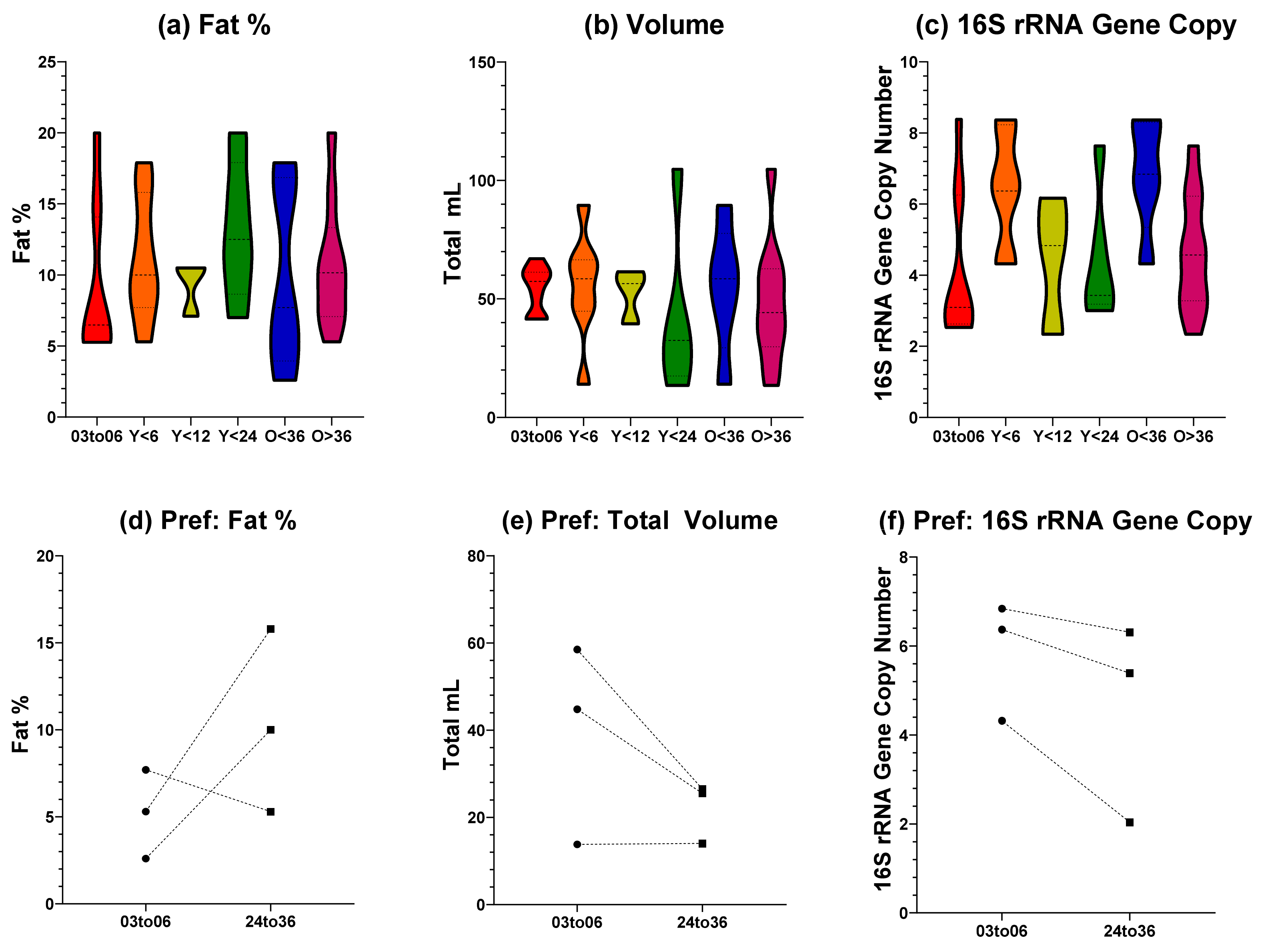
3.1. Fat, Volume, and Estimated Bacterial Load Does Not Differ between Human Milk from Tandem Feeding Dyads and Sole Nurslings
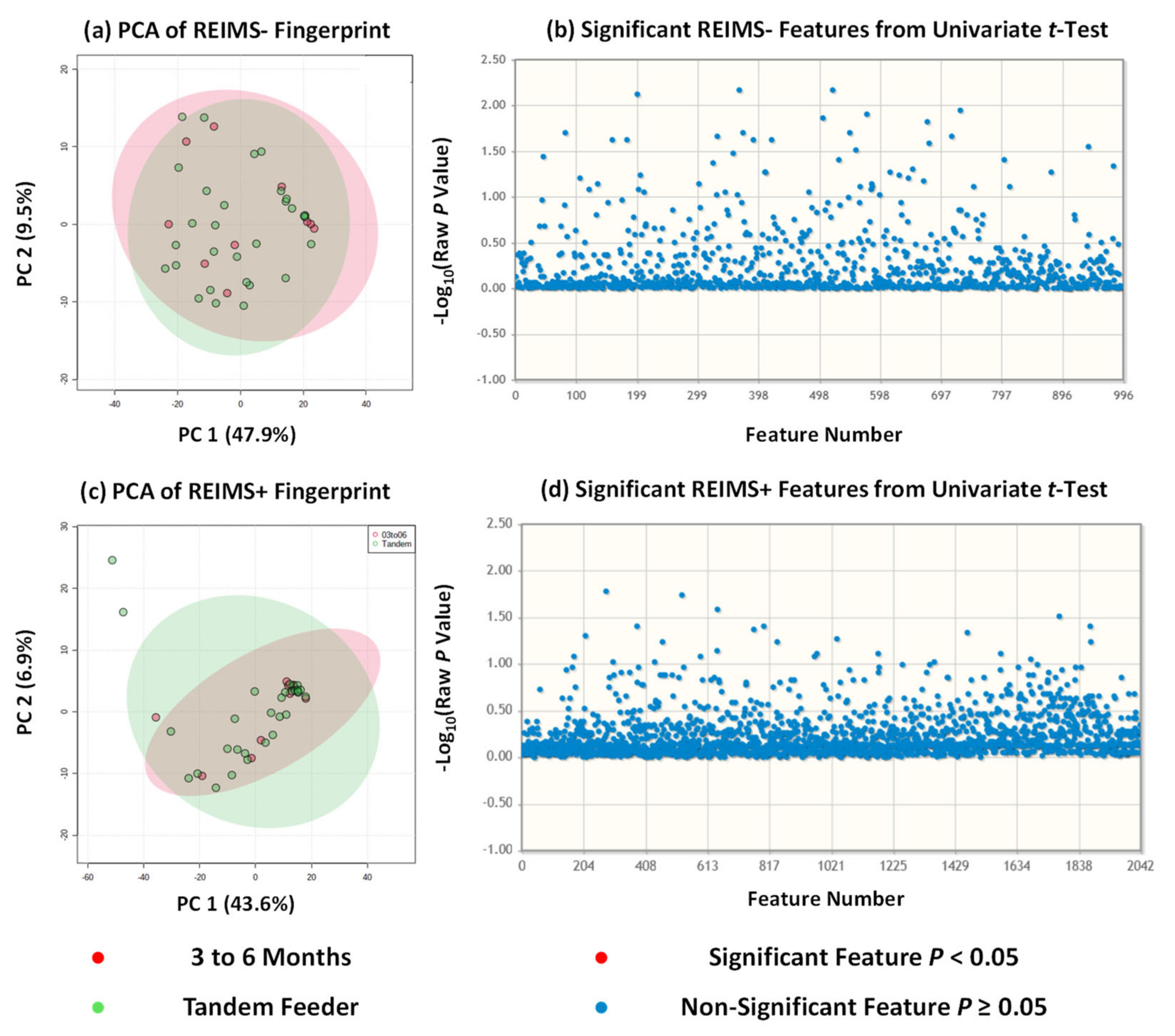
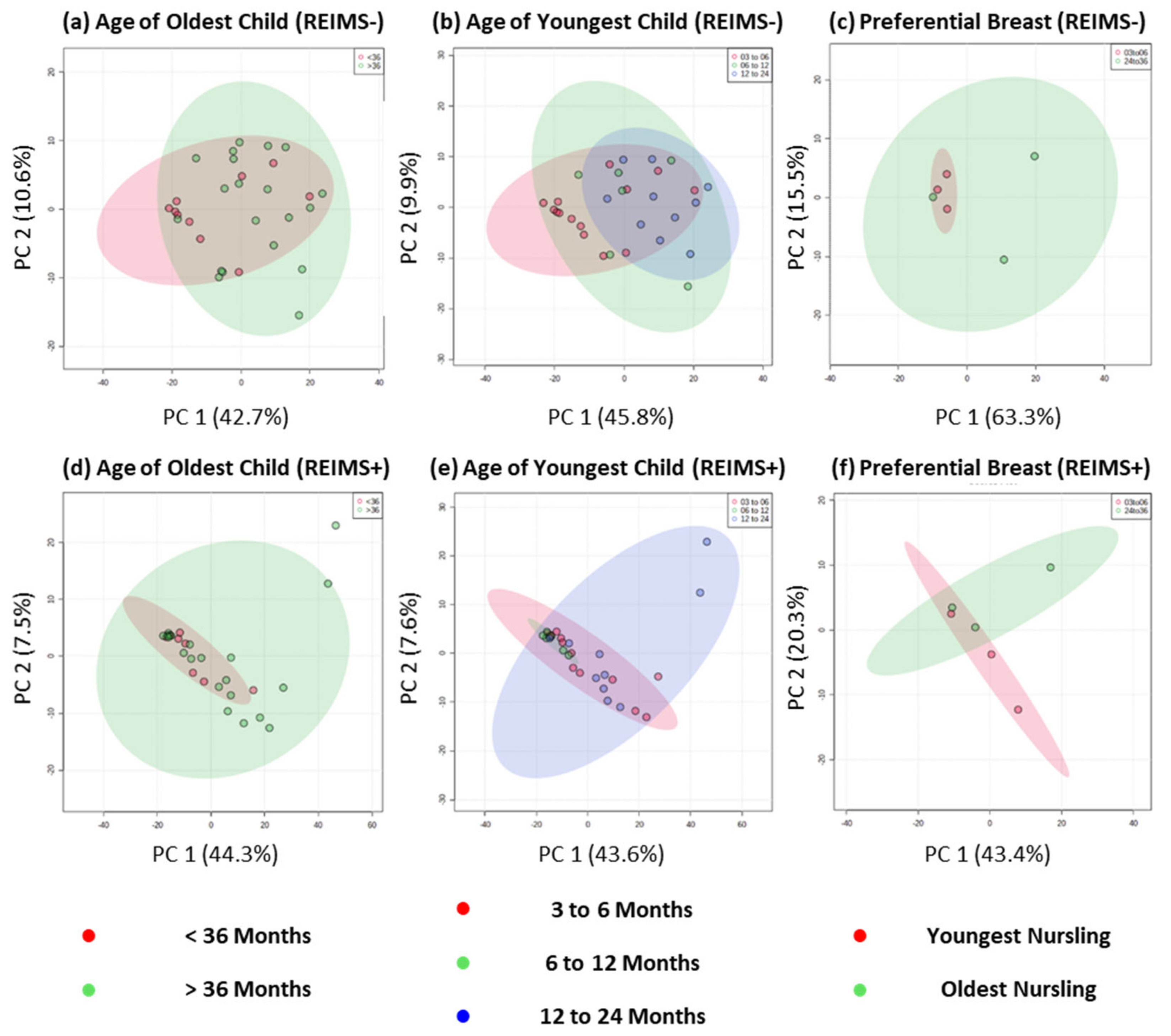
3.2. Metabolite Fingerprinting Using REIMS Shows no Multivariate or Univariate Differences for Dyad Groups
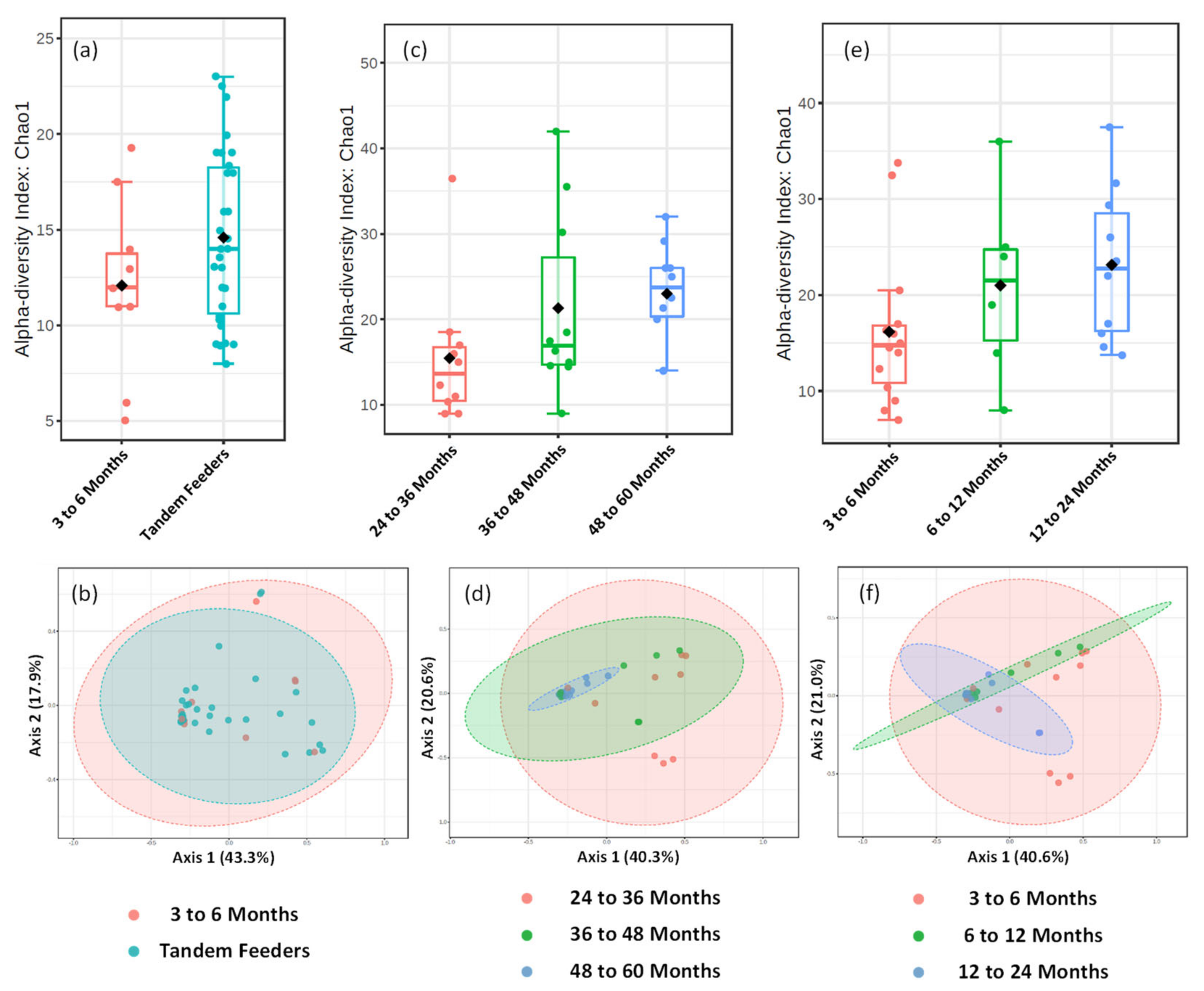
3.3. Metataxonomic Alpha or Beta Diversity Metrics Do Not Differ between Dyad Groups
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bryant, T. Tandem Nursing: A Review and Guidelines. Int. J. Childbirth Educ. 2012, 27, 84–88. [Google Scholar]
- Renfrew, M.; McAndrew, F.; Thompson, J.; Fellows, L.; Large, A.; Speed, M. Infant Feeding Survey 2010; Health and Social Care Information Centre: Leeds, UK, 2012; p. 2. [Google Scholar]
- Gupta, P.M.; Perrine, C.G.; Chen, J.; Elam-Evans, L.D.; Flores-Ayala, R. Monitoring the World Health Organization global target 2025 for exclusive breastfeeding: Experience from the United States. J. Hum. Lact. 2017, 33, 578–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Zhao, M.; Magnussen, C.G.; Xi, B. Global prevalence of WHO infant feeding practices in 57 LMICs in 2010–2018 and time trends since 2000 for 44 LMICs. EClinicalMedicine 2021, 37, 100971. [Google Scholar]
- O’Rourke, M.P.; Spatz, D.L. Women’s experiences with tandem breastfeeding. MCN Am. J. Matern. Child Nurs. 2019, 44, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Binns, C.; Lee, M.; Low, W.Y. The long-term public health benefits of breastfeeding. Asia Pac. J. Public Health 2016, 28, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattari, M.; Serwint, J.R.; Levine, D.M. Maternal implications of breastfeeding: A review for the internist. Am. J. Med. 2019, 132, 912–920. [Google Scholar] [CrossRef]
- Baranowska, B.; Malinowska, M.; Stanaszek, E.; Sys, D.; Bączek, G.; Doroszewska, A.; Tataj-Puzyna, U.; Rabijewski, M. Extended breastfeeding in Poland: Knowledge of health care providers and attitudes on breastfeeding beyond infancy. J. Hum. Lact. 2019, 35, 371–380. [Google Scholar] [CrossRef]
- Merše Lovrincevic, K.; Lepicnik, V. tandem breastfeeding—Research on the knowledge of nursing and dietetics students. Obz. Zdr. Nege 2018, 52, 38–44. [Google Scholar]
- Shenker, N.S.; Perdones-Montero, A.; Burke, A.; Stickland, S.; McDonald, J.A.; Alexander-Hardiman, K.; Flanagan, J.; Takats, Z.; Cameron, S.J. Metabolomic and Metataxonomic Fingerprinting of Human Milk Suggests Compositional Stability over a Natural Term of Breastfeeding to 24 Months. Nutrients 2020, 12, 3450. [Google Scholar] [CrossRef]
- Perrin, M.T.; Fogleman, A.D.; Newburg, D.S.; Allen, J.C. A longitudinal study of human milk composition in the second year postpartum: Implications for human milk banking. Matern. Child Nutr. 2017, 13, e12239. [Google Scholar] [CrossRef] [Green Version]
- Verd, S.; Ginovart, G.; Calvo, J.; Ponce-Taylor, J.; Gaya, A. Variation in the protein composition of human milk during extended lactation: A narrative review. Nutrients 2018, 10, 1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czosnykowska-Łukacka, M.; Królak-Olejnik, B.; Orczyk-Pawiłowicz, M. Breast Milk Macronutrient Components in Prolonged Lactation. Nutrients 2018, 10, 1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, J.E.; Hallam, J. Against all odds—Why UK mothers’ breastfeeding beyond infancy are turning to their international peers for emotional and informative support. Health Care Women Int. 2021, 42, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Sinkiewicz-Darol, E.; Bernatowicz-Łojko, U.; Łubiech, K.; Adamczyk, I.; Twarużek, M.; Baranowska, B.; Skowron, K.; Spatz, D.L. Tandem breastfeeding: A descriptive analysis of the nutritional value of milk when feeding a younger and older child. Nutrients 2021, 13, 277. [Google Scholar] [CrossRef] [PubMed]
- Cameron, S.; Huws, S.; Hegarty, M.J.; Smith, D.; Mur, L.A.J. The human salivary microbiome exhibits temporal stability in bacterial diversity. FEMS Microbiol. Ecol. 2015, 91, fiv091. [Google Scholar] [CrossRef]
- Cameron, S.J.; Perdones-Montero, A.; van Meulebroek, L.; Burke, A.; Alexander-Hardiman, K.; Simon, D.; Schaffer, R.; Balog, J.; Karancsi, T.; Rickards, T.; et al. Sample Preparation Free Mass Spectrometry Using Laser-Assisted Rapid Evaporative Ionization Mass Spectrometry: Applications to Microbiology, Metabolic Biofluid Phenotyping, and Food Authenticity. J. Am. Soc. Mass Spectrom. 2021, 32, 1393–1401. [Google Scholar] [CrossRef]
- Gibb, S.; Strimmer, K. MALDIquant: A versatile R package for the analysis of mass spectrometry data. Bioinformatics 2012, 28, 2270–2271. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Jewison, T.; Guo, A.C.; Wilson, M.; Knox, C.; Liu, Y.; Djoumbou, Y.; Mandal, R.; Aziat, F.; Dong, E.; et al. HMDB 3.0: The Human Metabolome Database in 2013. Nucleic Acids Res. 2013, 41, D801–D807. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making Metabolomics More Meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [Green Version]
- Bravi, F.; Wiens, F.; Decarli, A.; Dal Pont, A.; Agostoni, C.; Ferraroni, M. Impact of maternal nutrition on breast-milk composition: A systematic review. Am. J. Clin. Nutr. 2016, 104, 646–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czosnykowska-Łukacka, M.; Orczyk-Pawiłowicz, M.; Broers, B.; Królak-Olejnik, B. Lactoferrin in Human Milk of Prolonged Lactation. Nutrients 2019, 11, 2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer, K.C.; Dénes, J.; Albrecht, K.; Szaniszló, T.; Balog, J.; Skoumal, R.; Katona, M.; Tóth, M.; Balogh, L.; Takáts, Z. In vivo, in situ tissue analysis using rapid evaporative ionization mass spectrometry. Angew. Chem. Int. Ed. 2009, 48, 8240–8242. [Google Scholar] [CrossRef] [PubMed]
- Bardin, E.E.; Cameron, S.J.; Perdones-Montero, A.; Hardiman, K.; Bolt, F.; Alton, E.W.; Bush, A.; Davies, J.C.; Takáts, Z. Metabolic Phenotyping and Strain Characterisation of Pseudomonas aeruginosa Isolates from Cystic Fibrosis Patients Using Rapid Evaporative Ionisation Mass Spectrometry. Sci. Rep. 2018, 8, 10952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, S.J.; Bodai, Z.; Temelkuran, B.; Perdones-Montero, A.; Bolt, F.; Burke, A.; Alexander-Hardiman, K.; Salzet, M.; Fournier, I.; Rebec, M.; et al. Utilisation of Ambient Laser Desorption Ionisation Mass Spectrometry (ALDI-MS) Improves Lipid-Based Microbial Species Level Identification. Sci. Rep. 2019, 9, 3006. [Google Scholar] [CrossRef] [Green Version]
- Bolt, F.; Cameron, S.J.; Karancsi, T.; Simon, D.; Schaffer, R.; Rickards, T.; Hardiman, K.; Burke, A.; Bodai, Z.; Perdones-Montero, A.; et al. Automated High-Throughput Identification and Characterization of Clinically Important Bacteria and Fungi using Rapid Evaporative Ionization Mass Spectrometry. Anal. Chem. 2016, 88, 9419–9426. [Google Scholar] [CrossRef]
- Gowers, G.-O.F.; Cameron, S.J.; Perdones-Montero, A.; Bell, D.; Chee, S.M.; Kern, M.; Tew, D.; Ellis, T.; Takáts, Z. Off-Colony Screening of Biosynthetic Libraries by Rapid Laser-Enabled Mass Spectrometry. ACS Synth. Biol. 2019, 8, 2566–2575. [Google Scholar] [CrossRef]
- Cameron, S.J.; Bolt, F.; Perdones-Montero, A.; Rickards, T.; Hardiman, K.; Abdolrasouli, A.; Burke, A.; Bodai, Z.; Karancsi, T.; Simon, D.; et al. Rapid Evaporative Ionisation Mass Spectrometry (REIMS) Provides Accurate Direct from Culture Species Identification within the Genus Candida. Sci. Rep. 2016, 6, 36788. [Google Scholar] [CrossRef] [Green Version]
- Van Meulebroek, L.; Cameron, S.; Plekhova, V.; de Spiegeleer, M.; Wijnant, K.; Michels, N.; de Henauw, S.; Lapauw, B.; Takats, Z.; Vanhaecke, L. Rapid LA-REIMS and comprehensive UHPLC-HRMS for metabolic phenotyping of feces. Talanta 2020, 217, 121043. [Google Scholar] [CrossRef]
- Paraskevaidi, M.; Cameron, S.J.; Whelan, E.; Bowden, S.; Tzafetas, M.; Mitra, A.; Semertzidou, A.; Athanasiou, A.; Bennett, P.R.; MacIntyre, D.A.; et al. Laser-assisted rapid evaporative ionisation mass spectrometry (LA-REIMS) as a metabolomics platform in cervical cancer screening. EBioMedicine 2020, 60, 103017. [Google Scholar] [CrossRef]
- Cameron, S.J.; Alexander, J.L.; Bolt, F.; Burke, A.; Ashrafian, H.; Teare, J.; Marchesi, J.R.; Kinross, J.; Li, J.V.; Takats, Z. Evaluation of Direct from Sample Metabolomics of Human Faeces using Rapid Evaporative Ionisation Mass Spectrometry (REIMS). Anal. Chem. 2019, 91, 13448–13457. [Google Scholar] [CrossRef] [PubMed]
- Cameron, S.J.; Takáts, Z. Mass Spectrometry Approaches to Metabolic Profiling of Microbial Communities within the Human Gastrointestinal Tract. Methods 2018, 149, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Forstmeier, W.; Wagenmakers, E.J.; Parker, T.H. Detecting and avoiding likely false-positive findings—A practical guide. Biol. Rev. 2017, 92, 1941–1968. [Google Scholar] [CrossRef]
- Pines, N.; Mandel, D.; Mimouni, F.B.; Moran Lev, H.; Mangel, L.; Lubetzky, R. The effect of between-breast differences on human milk macronutrients content. J. Perinatol. 2016, 36, 549–551. [Google Scholar] [CrossRef] [PubMed]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K.; et al. Association between breast milk bacterial communities and establishment and development of the infant gut microbiome. JAMA Pediatr. 2017, 171, 647–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moossavi, S.; Azad, M.B. Origins of human milk microbiota: New evidence and arising questions. Gut Microb. 2020, 12, 1667722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moossavi, S.; Sepehri, S.; Robertson, B.; Bode, L.; Goruk, S.; Field, C.J.; Lix, L.M.; de Souza, R.J.; Becker, A.B.; Mandhane, P.J.; et al. Composition and variation of the human milk microbiota are influenced by maternal and early-life factors. Cell Host Microbe 2019, 25, 324–335.e4. [Google Scholar] [CrossRef] [Green Version]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and Laboratory Contamination can Critically Impact Sequence-Based Microbiome Analyses. BMC Biol. 2014, 12, 87. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.E.; Carrothers, J.M.; Lackey, K.A.; Beatty, N.F.; York, M.A.; Brooker, S.L.; Shafii, B.; Price, W.J.; Settles, M.L.; McGuire, M.A.; et al. Human milk microbial community structure is relatively stable and related to variations in macronutrient and micronutrient intakes in healthy lactating women. J. Nutr. 2017, 147, 1739–1748. [Google Scholar]
- Gonzalez, E.; Brereton, N.J.B.; Li, C.; Leyva, L.L.; Solomons, N.W.; Agellon, L.B.; Scott, M.E.; Koski, K.G. Distinct Changes Occur in the Human Breast Milk Microbiome Between Early and Established Lactation in Breastfeeding Guatemalan Mothers. Front. Microbiol. 2021, 12, 557180. [Google Scholar] [CrossRef]
- Cameron, S.J.; Lewis, K.E.; Huws, S.A.; Hegarty, M.J.; Lewis, P.D.; Pachebat, J.A.; Mur, L.A. A Pilot Study using Metagenomic Sequencing of the Sputum Microbiome Suggests Potential Bacterial Biomarkers for Lung Cancer. PLoS ONE 2017, 12, e0177062. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tandem Feeders | 3 to 6 Months | Preferential Side | p Value | ||
|---|---|---|---|---|---|
| Total Participants | 15 | 10 | 3 | N/A | |
| Age (Years) | 25.3 (4.9) | 26.9 (5.9) | 27.4 (4.7) | 0.675 | |
| Ethnicity | Black-Caribbean | 1 | 0 | 0 | <0.001 |
| Latin American | 1 | 0 | 1 | ||
| White | 13 | 10 | 2 | ||
| Body Mass Index | Pre-Pregnancy | 23.1 (3.6) | 23.9 (5.5) | 24.5 (5.2) | 0.864 |
| Post-Pregnancy | 25.3 (4.9) | 23.9 (5.5) | 24.5 (5.2) | 0.786 | |
| Gender | Boy | 11 | 4 | 3 | 0.131 |
| Girl | 4 | 6 | 0 | ||
| Diet | Meat Eater | 8 | 9 | 2 | <0.001 |
| Pescetarian | 1 | 1 | 0 | ||
| Vegetarian | 4 | 0 | 0 | ||
| Vegan | 2 | 0 | 1 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shenker, N.S.; Perdones-Montero, A.; Burke, A.; Stickland, S.; McDonald, J.A.K.; Cameron, S.J.S. Human Milk from Tandem Feeding Dyads Does Not Differ in Metabolite and Metataxonomic Features When Compared to Single Nursling Dyads under Six Months of Age. Metabolites 2022, 12, 1069. https://doi.org/10.3390/metabo12111069
Shenker NS, Perdones-Montero A, Burke A, Stickland S, McDonald JAK, Cameron SJS. Human Milk from Tandem Feeding Dyads Does Not Differ in Metabolite and Metataxonomic Features When Compared to Single Nursling Dyads under Six Months of Age. Metabolites. 2022; 12(11):1069. https://doi.org/10.3390/metabo12111069
Chicago/Turabian StyleShenker, Natalie S., Alvaro Perdones-Montero, Adam Burke, Sarah Stickland, Julie A. K. McDonald, and Simon J. S. Cameron. 2022. "Human Milk from Tandem Feeding Dyads Does Not Differ in Metabolite and Metataxonomic Features When Compared to Single Nursling Dyads under Six Months of Age" Metabolites 12, no. 11: 1069. https://doi.org/10.3390/metabo12111069