
Congruence and Complementarity of Differential Mobility Spectrometry and NMR Spectroscopy for Plasma Lipidomics
 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Consumables
2.2. Collection of Plasma Samples
2.3. Sample Preparation
2.4. NMR Experiments and Data Processing
2.5. DMS-SLA Experiments and Data Processing
3. Results and Discussion
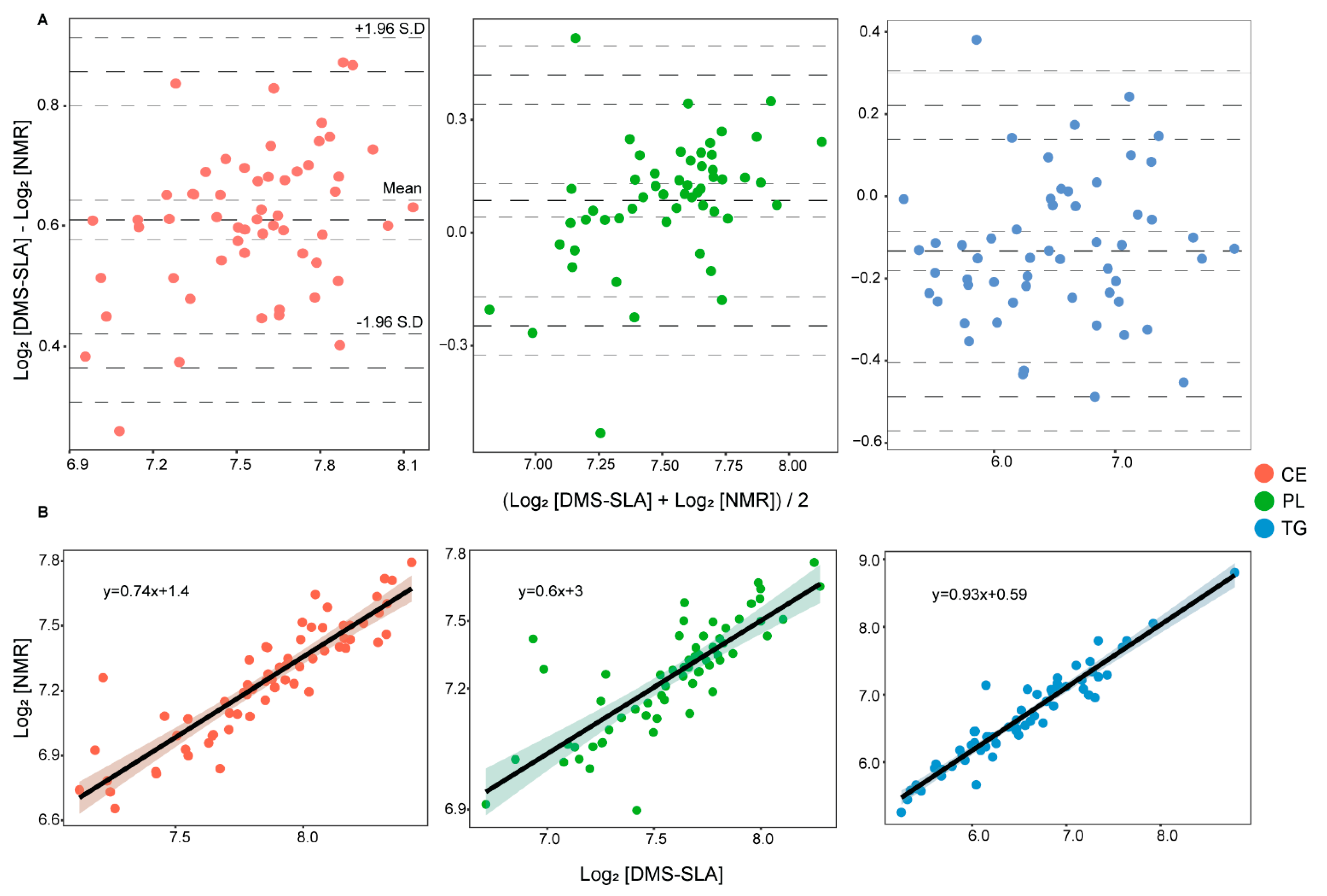
3.1. Comparison of NMR and DMS-SLA Platform
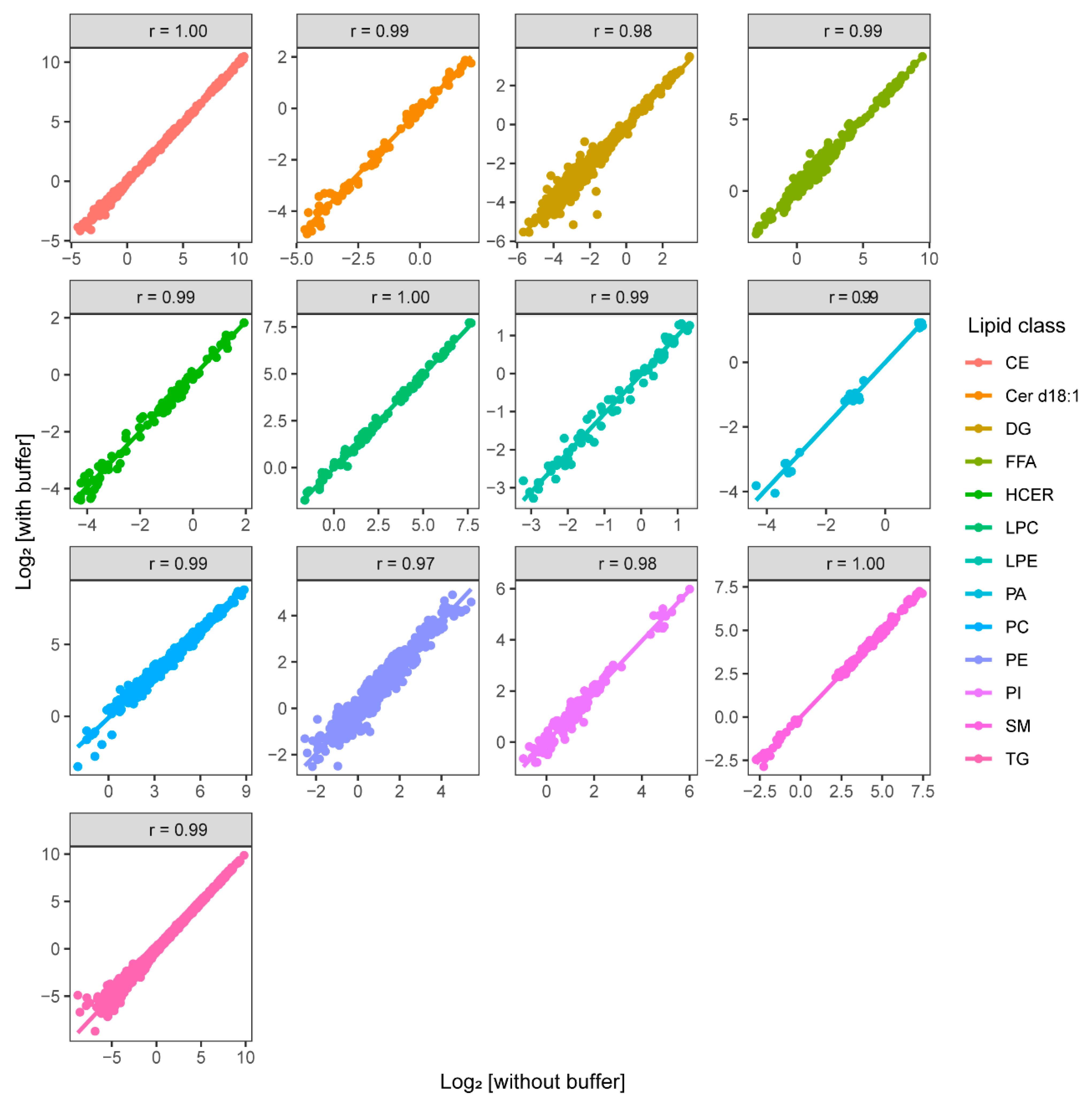
3.2. Effect of Buffer Addition on DMS-SLA Measurements
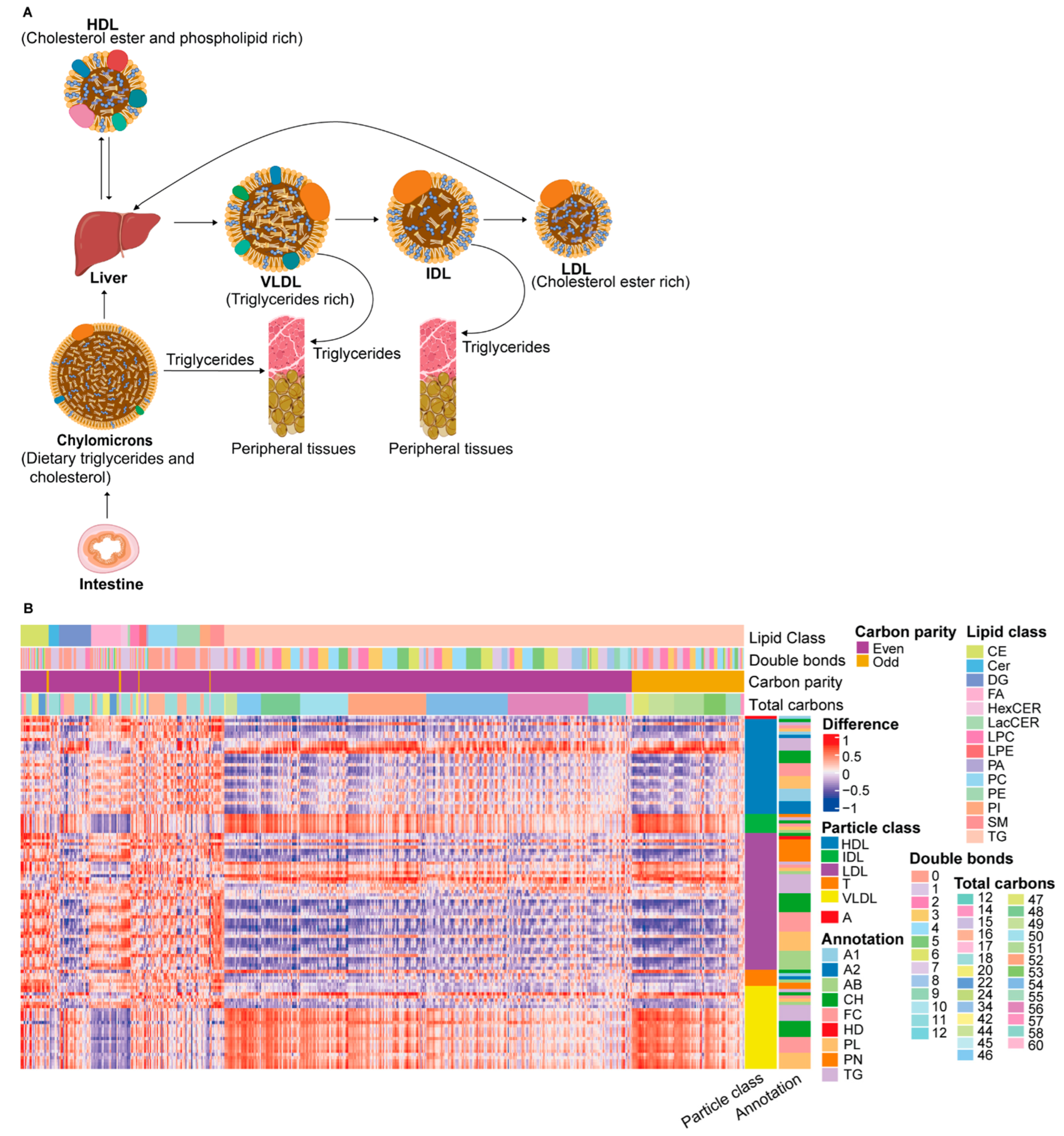
3.3. Correlation between Lipids and Lipoprotein Particles
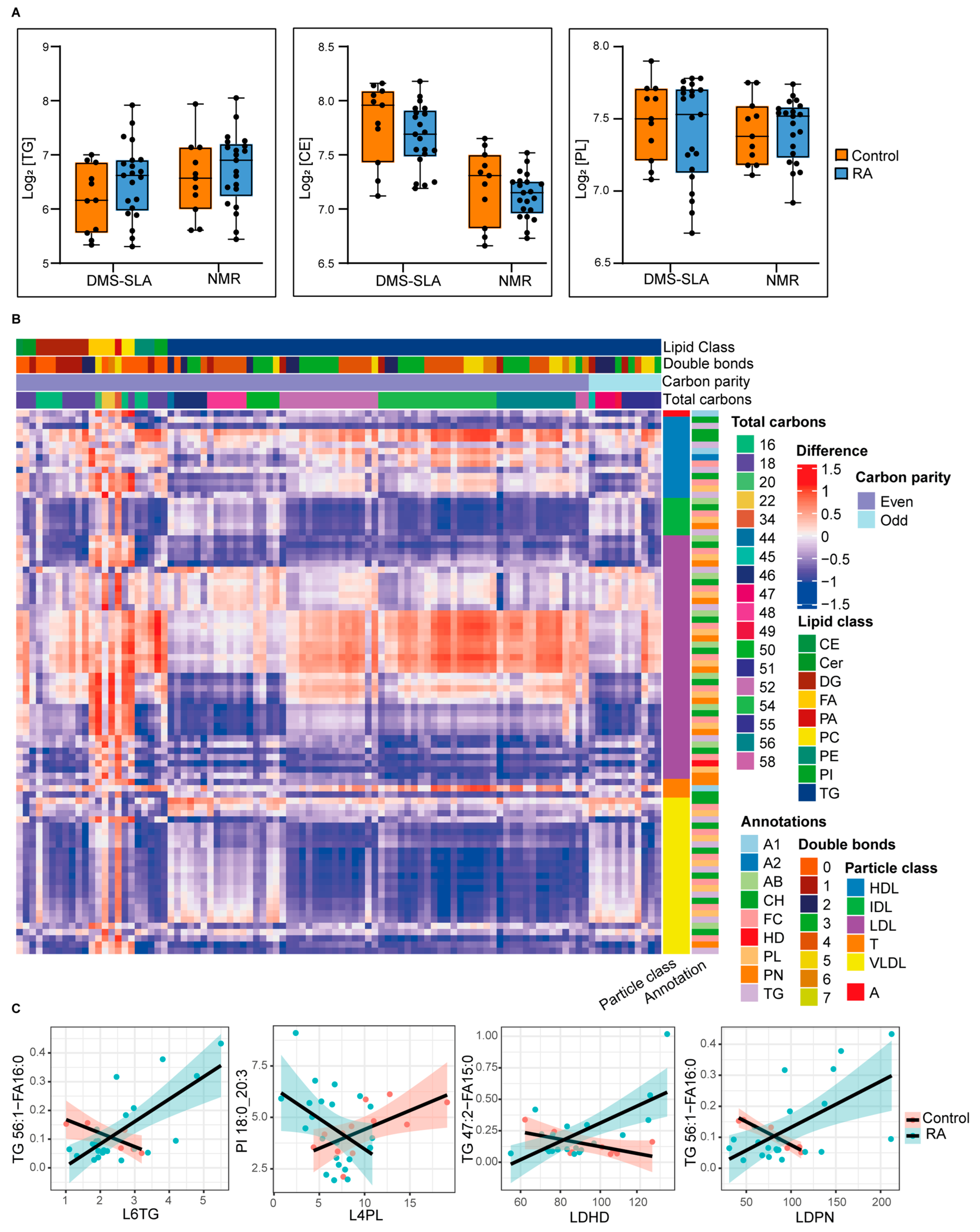
3.4. Comparative Analysis among Healthy Control and RA Patient Plasma
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fahy, E.; Cotter, D.; Sud, M.; Subramaniam, S. Lipid classification, structures and tools. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2011, 1811, 637–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunstone, F.D. Fatty Acid and Lipid Chemistry; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Quehenberger, O.; Armando, A.M.; Brown, A.H.; Milne, S.B.; Myers, D.S.; Merrill, A.H.; Bandyopadhyay, S.; Jones, K.N.; Kelly, S.; Shaner, R.L. Lipidomics reveals a remarkable diversity of lipids in human plasma 1. J. Lipid Res. 2010, 51, 3299–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghorasaini, M.; Mohammed, Y.; Adamski, J.; Bettcher, L.; Bowden, J.A.; Cabruja, M.; Contrepois, K.; Ellenberger, M.; Gajera, B.; Haid, M. Cross-laboratory standardization of preclinical lipidomics using differential mobility spectrometry and multiple reaction monitoring. Anal. Chem. 2021, 93, 16369–16378. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Simons, K. Lipidomics: Coming to grips with lipid diversity. Nat. Rev. Mol. Cell Biol. 2010, 11, 593–598. [Google Scholar] [CrossRef]
- Alabdulkarim, B.; Bakeet, Z.A.N.; Arzoo, S. Role of some functional lipids in preventing diseases and promoting health. J. King Saud Univ. Sci. 2012, 24, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Su, B.; Bettcher, L.F.; Hsieh, W.-Y.; Hornburg, D.; Pearson, M.J.; Blomberg, N.; Giera, M.; Snyder, M.P.; Raftery, D.; Bensinger, S.J. A DMS shotgun lipidomics workflow application to facilitate high-throughput, comprehensive lipidomics. J. Am. Soc. Mass Spectrom. 2021, 32, 2655–2663. [Google Scholar] [CrossRef]
- Medina, J.; Borreggine, R.; Teav, T.; Gao, L.; Ji, S.; Carrard, J.; Jones, C.; Blomberg, N.; Jech, M.; Atkins, A. ‘Omic-scale quantitative HILIC-MS/MS approach for circulatory lipid phenotyping in clinical research. Chemrxiv 2022. [Google Scholar] [CrossRef]
- Mahley, R.W.; Innerarity, T.L.; Rall, S.C., Jr.; Weisgraber, K.H. Plasma lipoproteins: Apolipoprotein structure and function. J. Lipid Res. 1984, 25, 1277–1294. [Google Scholar] [CrossRef]
- Osborne, J.C., Jr.; Brewer, H.B., Jr. The plasma lipoproteins. Adv. Protein Chem. 1977, 31, 253–337. [Google Scholar]
- Straat, M.E.; Martinez-Tellez, B.; Nahon, K.J.; Janssen, L.G.; Verhoeven, A.; van der Zee, L.; Mulder, M.T.; Kooijman, S.; Boon, M.R.; van Lennep, J.E.R. Comprehensive (apo) lipoprotein profiling in patients with genetic hypertriglyceridemia using LC-MS and NMR spectroscopy. J. Clin. Lipidol. 2022, 16, 472–482. [Google Scholar] [CrossRef]
- Julkunen, H.; Cichońska, A.; Tiainen, M.; Koskela, H.; Nybo, K.; Mäkelä, V.; Nokso-Koivisto, J.; Kristiansson, K.; Perola, M.; Salomaa, V. Atlas of plasma nuclear magnetic resonance biomarkers for health and disease in 118,461 individuals from the UK Biobank. medRxiv 2022. [Google Scholar] [CrossRef]
- Alarcon-Barrera, J.C.; von Hegedus, J.H.; Brouwers, H.; Steenvoorden, E.; Ioan-Facsinay, A.; Mayboroda, O.A.; Ondo-Mendez, A.; Giera, M. Lipid metabolism of leukocytes in the unstimulated and activated states. Anal. Bioanal. Chem. 2020, 412, 2353–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loef, M.; von Hegedus, J.H.; Ghorasaini, M.; Kroon, F.P.; Giera, M.; Ioan-Facsinay, A.; Kloppenburg, M. Reproducibility of targeted lipidome analyses (lipidyzer) in plasma and erythrocytes over a 6-week period. Metabolites 2020, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Vosegaard, T.; Guo, Z. Applications of nuclear magnetic resonance in lipid analyses: An emerging powerful tool for lipidomics studies. Prog. Lipid Res. 2017, 68, 37–56. [Google Scholar] [CrossRef]
- Jiménez, B.; Holmes, E.; Heude, C.; Tolson, R.F.; Harvey, N.; Lodge, S.L.; Chetwynd, A.J.; Cannet, C.; Fang, F.; Pearce, J.T. Quantitative lipoprotein subclass and low molecular weight metabolite analysis in human serum and plasma by 1H NMR spectroscopy in a multilaboratory trial. Anal. Chem. 2018, 90, 11962–11971. [Google Scholar] [CrossRef]
- Lodge, S.; Nitschke, P.; Kimhofer, T.; Coudert, J.D.; Begum, S.; Bong, S.-H.; Richards, T.; Edgar, D.; Raby, E.; Spraul, M. NMR spectroscopic windows on the systemic effects of SARS-CoV-2 infection on plasma lipoproteins and metabolites in relation to circulating cytokines. J. Proteome Res. 2021, 20, 1382–1396. [Google Scholar] [CrossRef]
- Kay, J.; Upchurch, K.S. ACR/EULAR 2010 rheumatoid arthritis classification criteria. Rheumatology 2012, 51 (Suppl. 6), vi5–vi9. [Google Scholar] [CrossRef] [Green Version]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Findeisen, M.; Brand, T.; Berger, S. A 1H-NMR thermometer suitable for cryoprobes. Magn. Reson. Chem. 2007, 45, 175–178. [Google Scholar] [CrossRef]
- Wu, P.S.; Otting, G. Rapid pulse length determination in high-resolution NMR. J. Magn. Reson. 2005, 176, 115–119. [Google Scholar] [CrossRef]
- Price, W.S. Water signal suppression in NMR spectroscopy. In Annual Reports on NMR Spectroscopy; Elsevier: Amsterdam, The Netherlands, 1999; Volume 38, pp. 289–354. [Google Scholar]
- Kumar, A. Two-dimensional nuclear Overhauser effect experiment in a protein: The first NOESY (1979–80). Magn. Reson. Chem. 2003, 41 (Suppl. 1), S26–S32. [Google Scholar] [CrossRef]
- Eksborg, S. Evaluation of method-comparison data. Clin. Chem. 1981, 27, 1311–1312. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G.; Bland, J.M. Measurement in medicine: The analysis of method comparison studies. J. R. Stat. Soc. Ser. D (Stat.) 1983, 32, 307–317. [Google Scholar] [CrossRef]
- Iavarone, A.T.; Udekwu, O.A.; Williams, E.R. Buffer loading for counteracting metal salt-induced signal suppression in electrospray ionization. Anal. Chem. 2004, 76, 3944–3950. [Google Scholar] [CrossRef] [Green Version]
- Feingold, K.R.; Grunfeld, C. Introduction to Lipids and Lipoproteins; MDText.com, Inc.: South Dartmouth, MA, USA, 2015. [Google Scholar]
- Lewis, G.F. Fatty acid regulation of very low density lipoprotein production. Curr. Opin. Lipidol. 1997, 8, 146–153. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Lee, E.-Y.; Park, J.K.; Song, Y.W.; Kim, J.-R.; Cho, K.-H. Patients with rheumatoid arthritis show altered lipoprotein profiles with dysfunctional high-density lipoproteins that can exacerbate inflammatory and atherogenic process. PLoS ONE 2016, 11, e0164564. [Google Scholar] [CrossRef] [Green Version]
- Choy, E.; Sattar, N. Interpreting lipid levels in the context of high-grade inflammatory states with a focus on rheumatoid arthritis: A challenge to conventional cardiovascular risk actions. Ann. Rheum. Dis. 2009, 68, 460–469. [Google Scholar] [CrossRef]
- Steiner, G.; Urowitz, M.B. Lipid Profiles in Patients with Rheumatoid Arthritis: Mechanisms and the Impact of Treatment. Semin. Arthritis Rheum. 2009, 38, 372–381. [Google Scholar] [CrossRef]
- Charles-Schoeman, C.; Fleischmann, R.; Davignon, J.; Schwartz, H.; Turner, S.M.; Beysen, C.; Milad, M.; Hellerstein, M.K.; Luo, Z.; Kaplan, I.V. Potential mechanisms leading to the abnormal lipid profile in patients with rheumatoid arthritis versus healthy volunteers and reversal by tofacitinib. Arthritis Rheumatol. 2015, 67, 616–625. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Class | Concentration | Pearson r | p-Value | Spearman ρ | p-Value | Mean (DMS-SLA) | Mean (NMR) | RMSD | NRMSD |
|---|---|---|---|---|---|---|---|---|---|
| Triacyl glycerides | mg/dL | 0.97 | 1.4 × 10−38 | 5.04 × 10−32 | 102.1 | 111.39 | 16.5 | 14.82% | |
| log2 [mg/dL] | 0.95 | 6.15 × 10−34 | 0.95 | 5.04 × 10−32 | 6.48 | 6.62 | 0.20 | 3.07% | |
| Cholesterol esters | mg/dL | 0.89 | 8.06 × 10−23 | 0.92 | 1.55 × 10−26 | 237.41 | 154.79 | 12.03 | 7.77% |
| log2 [mg/dL] | 0.89 | 8.06 × 10−23 | 0.92 | 1.55 × 10−26 | 7.86 | 7.25 | 0.12 | 1.62% | |
| Phospholipids | mg/dL | 0.81 | 1.56 × 10−15 | 0.81 | 1.03 × 10−15 | 197.46 | 183.75 | 17.36 | 9.44% |
| log2 [mg/dL] | 0.79 | 2.30 × 10−14 | 0.81 | 1.03 × 10−15 | 7.59 | 7.50 | 0.146 | 1.95% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghorasaini, M.; Tsezou, K.I.; Verhoeven, A.; Mohammed, Y.; Vlachoyiannopoulos, P.; Mikros, E.; Giera, M. Congruence and Complementarity of Differential Mobility Spectrometry and NMR Spectroscopy for Plasma Lipidomics. Metabolites 2022, 12, 1030. https://doi.org/10.3390/metabo12111030
Ghorasaini M, Tsezou KI, Verhoeven A, Mohammed Y, Vlachoyiannopoulos P, Mikros E, Giera M. Congruence and Complementarity of Differential Mobility Spectrometry and NMR Spectroscopy for Plasma Lipidomics. Metabolites. 2022; 12(11):1030. https://doi.org/10.3390/metabo12111030
Chicago/Turabian StyleGhorasaini, Mohan, Konstantina Ismini Tsezou, Aswin Verhoeven, Yassene Mohammed, Panayiotis Vlachoyiannopoulos, Emmanuel Mikros, and Martin Giera. 2022. "Congruence and Complementarity of Differential Mobility Spectrometry and NMR Spectroscopy for Plasma Lipidomics" Metabolites 12, no. 11: 1030. https://doi.org/10.3390/metabo12111030