
Revealing the Phenolic Acids in Cardamine violifolia Leaves by Transcriptome and Metabolome Analyses
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Measurement of Total Phenolic Acid Content
2.3. Widely Targeted Metabolome Detection and Analysis
2.4. Transcriptome Sequencing and Analysis
2.5. Conjoint Analysis of Transcriptome and Metabolome
2.6. Real-Time Quantitative PCR (RT-qPCR) Validation
2.7. Statistical Analysis
3. Results
3.1. Content of Total Phenolic Acids
3.2. Overview of the Metabolome Detection
3.3. Analysis of Phenolic Acids
3.4. Transcriptome Sequencing and Annotation
3.5. Analysis of DEGs
3.6. Conjoint Analysis of Transcriptome and Metabolome
3.7. Correlation between Transcriptome and Real-Time PCR (RT-qPCR) Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rao, S.; Yu, T.; Cong, X.; Lai, X.; Xiang, J.; Cao, J.; Liao, X.; Gou, Y.; Chao, W.; Xue, H.; et al. Transcriptome, proteome, and metabolome reveal the mechanism of tolerance to selenate toxicity in Cardamine violifolia. J. Hazard. Mater. 2021, 406, 124283. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Cong, X.; Li, M.; Rao, S.; Liu, Y.; Guo, J.; Zhu, S.; Chen, S.; Xu, F.; Cheng, S.; et al. Effects of different exogenous selenium on Se accumulation, nutrition quality, elements uptake, and antioxidant response in the hyperaccumulation plant Cardamine violifolia. Ecotox. Environ. Safe. 2020, 204, 111045. [Google Scholar] [CrossRef]
- Rao, S.; Yu, T.; Cong, X.; Xu, F.; Lai, X.; Zhang, W.; Liao, Y.; Cheng, S. Integration analysis of PacBio SMRT- and Illumina RNA-seq reveals candidate genes and pathway involved in selenium metabolism in hyperaccumulator Cardamine violifolia. BMC Plant Biol. 2020, 20, 492. [Google Scholar] [CrossRef]
- Both, E.B.; Stonehouse, G.C.; Lima, L.W.; Fakra, S.C.; Aguirre, B.; Wangeline, A.L.; Xiang, J.; Yin, H.; Jókai, Z.; Soós, Á.; et al. Selenium tolerance, accumulation, localization and speciation in a Cardamine hyperaccumulator and a non-hyperaccumulator. Sci. Total Environ. 2020, 703, 135041. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhu, Y.; Lin, Z.; Banuelos, G.; Li, W.; Yin, X. A novel Selenocystine-Accumulating plant in Selenium-Mine drainage area in enshi, China. PLoS ONE 2013, 8, e65615. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Xu, J.; He, X.; Yang, Z.; Fang, W.; Tao, J. Role of exogenous melatonin involved in phenolic acid metabolism of germinated hulless barley under NaCl stress. Plant Physiol. Biochem. 2022, 170, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Apea-Bah, F.B.; Drawbridge, P.; Beta, T. A generalized method for determining free soluble phenolic acid composition and antioxidant capacity of cereals and legumes. J. Vis. Exp. 2022, 184, e62467. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e370. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Huang, F.; Deng, C.; Wang, Y.; Kai, G. Bioactivities, biosynthesis and biotechnological production of phenolic acids in Salvia miltiorrhiza. Crit. Rev. Food Sci. 2018, 59, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Sachithanandam, V.; Parthiban, A.; Lalitha, P.; Muthukumaran, J.; Jain, M.; Elumalai, D.; Jayabal, K.; Sridhar, R.; Ramachandran, P.; Ramachandran, R. Biological evaluation of gallic acid and quercetin derived from Ceriops tagal: Insights from extensive in vitro and in silico studies. J. Biomol. Struct. Dyn. 2022, 40, 1490–1502. [Google Scholar] [CrossRef]
- Menard, G.N.; Langdon, M.; Bhunia, R.K.; Shankhapal, A.R.; Noleto-Dias, C.; Lomax, C.; Ward, J.L.; Kurup, S.; Eastmond, P.J. Diverting phenylpropanoid pathway flux from sinapine to produce industrially useful 4-vinyl derivatives of hydroxycinnamic acids in Brassicaceous oilseeds. Metab. Eng. 2022, 70, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Vries, S.; Fürst Jansen, J.M.R.; Irisarri, I.; Dhabalia Ashok, A.; Ischebeck, T.; Feussner, K.; Abreu, I.N.; Petersen, M.; Feussner, I.; Vries, J. The evolution of the phenylpropanoid pathway entailed pronounced radiations and divergences of enzyme families. Plant J. 2021, 107, 975–1002. [Google Scholar] [CrossRef] [PubMed]
- Valiñas, M.A.; Lanteri, M.L.; Ten Have, A.; Andreu, A.B. Chlorogenic acid biosynthesis appears linked with suberin production in potato tuber (Solanum tuberosum). J. Agric. Food Chem. 2015, 63, 4902–4913. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ding, Y.; Wang, Q.; Wang, P.; Han, Y.; Gu, Z.; Yang, R. NaCl treatment on physio-biochemical metabolism and phenolics accumulation in barley seedlings. Food Chem. 2020, 331, 127282. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, D.; Yang, D.; Xue, Z.; Li, J.; Xing, B.; Yan, K.; Han, R.; Liang, Z. SmKFB5 protein regulates phenolic acid biosynthesis by controlling the degradation of phenylalanine ammonia-lyase in Salvia miltiorrhiza. J. Exp. Bot. 2021, 72, 4915–4929. [Google Scholar] [CrossRef]
- Wang, B.; Sun, W.; Li, Q.; Li, Y.; Luo, H.; Song, J.; Sun, C.; Qian, J.; Zhu, Y.; Hayward, A.; et al. Genome-wide identification of phenolic acid biosynthetic genes in Salvia miltiorrhiza. Planta 2015, 241, 711–725. [Google Scholar] [CrossRef]
- Chu, C.; Du, Y.; Yu, X.; Shi, J.; Yuan, X.; Liu, X.; Liu, Y.; Zhang, H.; Zhang, Z.; Yan, N. Dynamics of antioxidant activities, metabolites, phenolic acids, flavonoids, and phenolic biosynthetic genes in germinating Chinese wild rice (Zizania latifolia). Food Chem. 2020, 318, 126483. [Google Scholar] [CrossRef]
- Wu, X.; Wei, W.; Yang, X.; Zhang, Y.; Xu, W.; Yang, Y.; Zhong, G.; Liu, H.; Yang, S. Anti-Inflammatory phenolic acid esters from the roots and rhizomes of Notopterygium incisium and their permeability in the human caco-2 monolayer cell model. Molecules 2017, 22, 935. [Google Scholar] [CrossRef] [Green Version]
- Clifford, M.N. Chlorogenic acids and other cinnamates—Nature, occurrence, dietary burden, absorption and metabolism. J. Sci. Food Agric. 2000, 80, 1033–1043. [Google Scholar] [CrossRef]
- Cavò, E.; Taviano, M.F.; Davì, F.; Cacciola, F.; Oulad El Majdoub, Y.; Mondello, L.; Ragusa, M.; Condurso, C.; Merlino, M.; Verzera, A.; et al. Phenolic and Volatile Composition and Antioxidant Properties of the Leaf Extract of Brassica fruticulosa subsp. Fruticulosa (Brassicaceae) Growing Wild in Sicily (Italy). Molecules 2022, 27, 2768. [Google Scholar]
- Linić, I.; Aamec, D.; Grúz, J.; Vujčić Bok, V.; Strnad, M.; Salopek-Sondi, B. Involvement of phenolic acids in Short-Term adaptation to salinity stress is Species-Specific among Brassicaceae. Plants 2019, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Yu, T.; Cong, X.; Zhang, W.; Zhu, Z.; Liao, Y.; Ye, J.; Cheng, S.; Xu, F. Effects of selenate applied at two growth stages on the nutrient quality of Cardamine violifolia. Sci. Hortic. 2021, 288, 110352. [Google Scholar] [CrossRef]
- Costa, C.; Tsatsakis, A.; Mamoulakis, C.; Teodoro, M.; Briguglio, G.; Caruso, E.; Tsoukalas, D.; Margina, D.; Dardiotis, E.; Kouretas, D.; et al. Current evidence on the effect of dietary polyphenols intake on chronic diseases. Food Chem. Toxicol. 2017, 110, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Hu, W.; Jiang, A.; Xu, Y.; Sa, R.; Feng, K.; Zhao, M.; Yu, J.; Ji, Y.; Hou, M.; et al. Effect of Methyl Jasmonate on Phenolic Accumulation in Wounded Broccoli. Molecules 2019, 24, 3537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seljåsen, R.; Kusznierewicz, B.; Bartoszek, A.; Mølmann, J.; Vågen, I.M. Effects of Post-Harvest Elicitor Treatments with Ultrasound, UV- and Photosynthetic Active Radiation on Polyphenols, Glucosinolates and Antioxidant Activity in a Waste Fraction of White Cabbage (Brassica oleracea var. Capitata). Molecules 2022, 27, 5256. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Liang, L.; Su, M.; Yang, T.; Mao, X.; Wang, Y. Variations in phenolic acids and antioxidant activity of navel orange at different growth stages. Food Chem. 2021, 360, 129980. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Li, X.; Qin, Y.; Li, M.; Han, Q.; Zhou, J.; Lin, S.; Zhao, L.; Zhang, Q.; Qin, W.; et al. Physicochemical properties, phenolic profiles, antioxidant capacities, and inhibitory effects on digestive enzymes of okra (Abelmoschus esculentus) fruit at different maturation stages. J. Food Sci. Technol. 2019, 56, 1275–1286. [Google Scholar] [CrossRef]
- Bai, N.; He, K.; Roller, M.; Lai, C.; Bai, L.; Pan, M. Flavonolignans and Other Constituents from Lepidium meyenii with Activities in Anti-inflammation and Human Cancer Cell Lines. J. Agric. Food Chem. 2015, 63, 2458–2463. [Google Scholar] [CrossRef]
- Zhou, W.; Jia, M.; Zhang, G.; Sun, J.; Li, Q.; Wang, X.; Hua, J.; Luo, S. Up-regulation of phenylpropanoid biosynthesis system in peach species by peach aphids produces anthocyanins that protect the aphids against UVB and UVC radiation. Tree Physiol. 2021, 41, 428–443. [Google Scholar] [CrossRef]
- Benković, V.; Marčina, N.; Horvat Knežević, A.; Aikić, D.; Rajevac, V.; Milić, M.; Kopjar, N. Potential radioprotective properties of arbutin against ionising radiation on human leukocytes in vitro. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2021, 872, 503413. [Google Scholar] [CrossRef]
- Wu, L.; Li, P.; Zhao, Q.; Piao, J.; Jiao, Y.; Kadowaki, M.; Kondo, T. Arbutin, an intracellular hydroxyl radical scavenger, protects radiation-induced apoptosis in human lymphoma U937 cells. Apoptosis 2014, 19, 1654–1663. [Google Scholar] [CrossRef] [PubMed]
- Hazman, Ö.; Sarıova, A.; Bozkurt, M.F.; Ciğerci, İ.H. The anticarcinogen activity of β-arbutin on MCF-7 cells: Stimulation of apoptosis through estrogen receptor-α signal pathway, inflammation and genotoxicity. Mol. Cell. Biochem. 2021, 476, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Radmard, A.; Saeedi, M.; Morteza-Semnani, K.; Hashemi, S.M.H.; Nokhodchi, A. An eco-friendly and green formulation in lipid nanotechnology for delivery of a hydrophilic agent to the skin in the treatment and management of hyperpigmentation complaints: Arbutin niosome (Arbusome). Colloids Surf. B Biointerfaces 2021, 201, 111616. [Google Scholar] [CrossRef]
- Wang, R.; Mu, J. Arbutin attenuates ethanol-induced acute hepatic injury by the modulation of oxidative stress and Nrf-2/HO-1 signaling pathway. J. Biochem. Mol. Toxicol. 2021, 35, e22872. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.J.; Ha, H.W.; Kim, H.G.; Lee, D.H.; Kong, M.J.; Ahn, Y.T.; Kim, D.H.; Shin, B.S.; Kang, W.; Jeong, H.G.; et al. Role of metabolism by intestinal bacteria in arbutin-induced toxicity in vitro. Arch. Pharm. Res. 2011, 34, 687–693. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, Y.; Chen, Q.; Kuang, Y.; Fan, J.; Xu, X.; Zhu, H.; Gao, Q.; Cheng, S.; Cong, X.; et al. Selenium-enriched Cardamine violifolia improves growth performance with potential regulation of intestinal health and antioxidant function in weaned pigs. Front. Vet. Sci. 2022, 9, 964766. [Google Scholar] [CrossRef]
- Wei, J.; Wang, S.; Pei, D.; Qu, L.; Li, Y.; Chen, J.; Di, D.; Gao, K. Antibacterial Activity of Hydroxytyrosol Acetate from Olive Leaves (Olea Europaea L.). Nat. Prod. Res. 2018, 32, 1967–1970. [Google Scholar] [CrossRef]
- Guo, D.; Fu, X.; Sun, Y.; Li, X.; Pan, H. De novo biosynthesis of tyrosol acetate and hydroxytyrosol acetate from glucose in engineered Escherichia coli. Enzym. Microb. Technol. 2021, 150, 109886. [Google Scholar] [CrossRef]
- Nair, R.B.; Joy, R.W.; Kurylo, E.; Shi, X.; Schnaider, J.; Datla, R.S.S.; Keller, W.A.; Selvaraj, G. Identification of a CYP84 family of cytochrome P450-Dependent Mono-Oxygenase genes in brassica napus and perturbation of their expression for engineering sinapine reduction in the seeds. Plant Physiol. 2000, 123, 1623–1634. [Google Scholar] [CrossRef] [Green Version]
- Tetreault, H.M.; Gries, T.; Palmer, N.A.; Funnell-Harris, D.L.; Sato, S.; Ge, Z.; Sarath, G.; Sattler, S.E. Overexpression of ferulate 5-hydroxylase increases syringyl units in Sorghum bicolor. Plant Mol. Biol. 2020, 103, 269–285. [Google Scholar] [CrossRef]
- Maruta, T.; Noshi, M.; Nakamura, M.; Matsuda, S.; Tamoi, M.; Ishikawa, T.; Shigeoka, S. Ferulic acid 5-hydroxylase 1 is essential for expression of anthocyanin biosynthesis-associated genes and anthocyanin accumulation under photooxidative stress in Arabidopsis. Plant Sci. 2014, 219, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.; Li, Y.; Mo, H.; Chapple, C. Assembly of an evolutionarily new pathway for α-Pyrone biosynthesis in Arabidopsis. Science 2012, 337, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, P.D.; Khan, B.M.; Gaikwad, S.M. Conformational transitions of cinnamoyl CoA reductase 1 from Leucaena leucocephala. Int. J. Biol. Macromol. 2014, 64, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wu, J.; Lin, L.; Li, P.; Li, S.; Wang, Y.; Li, J.; Sun, Q.; Liang, J.; Wang, Y. Overexpression of Cinnamoyl-CoA Reductase 2 in Brassica napus Increases Resistance to Sclerotinia sclerotiorum by Affecting Lignin Biosynthesis. Front. Plant Sci. 2021, 12, 732733. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
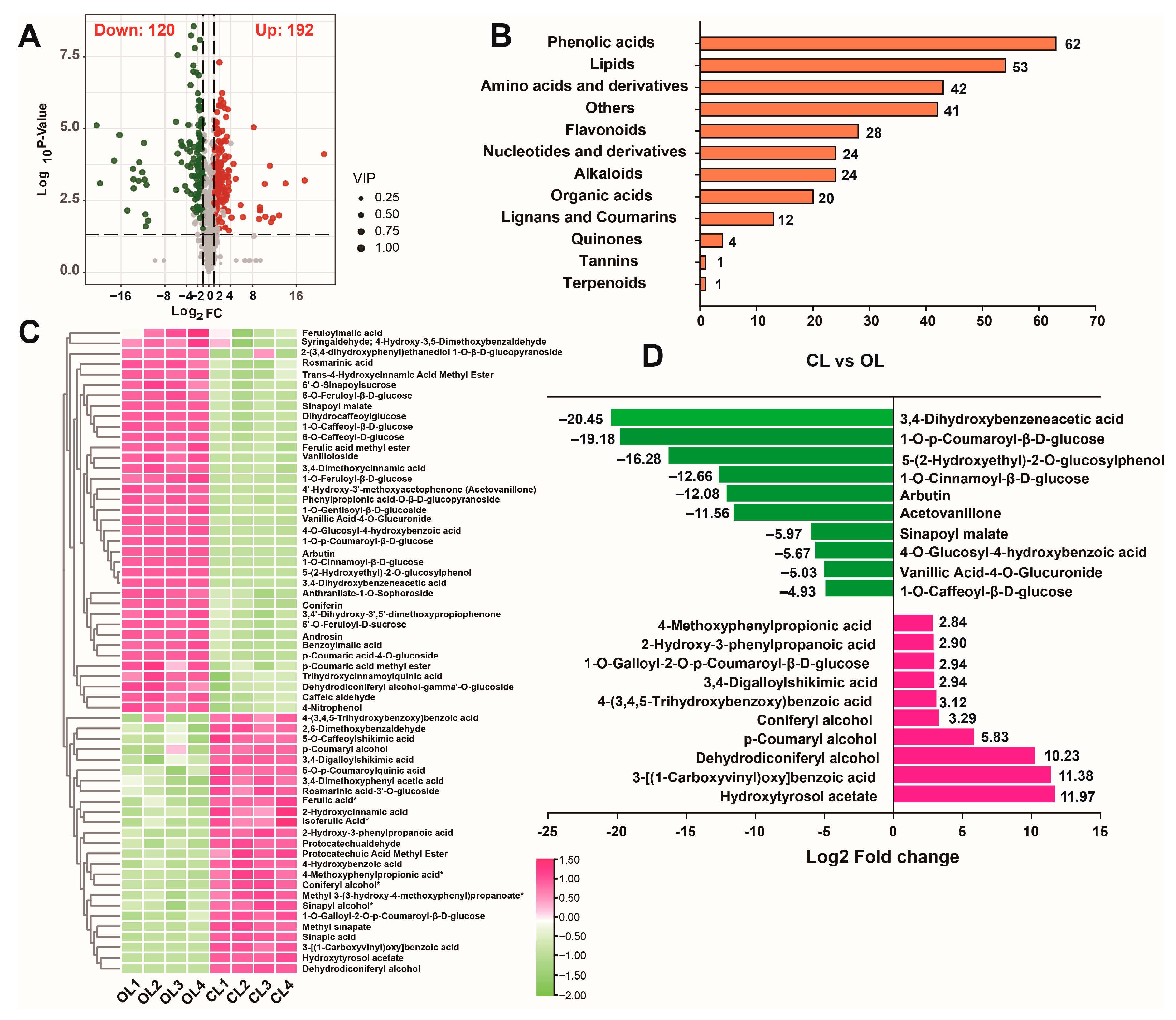{kind=link}
{kind=link}
{kind=link}
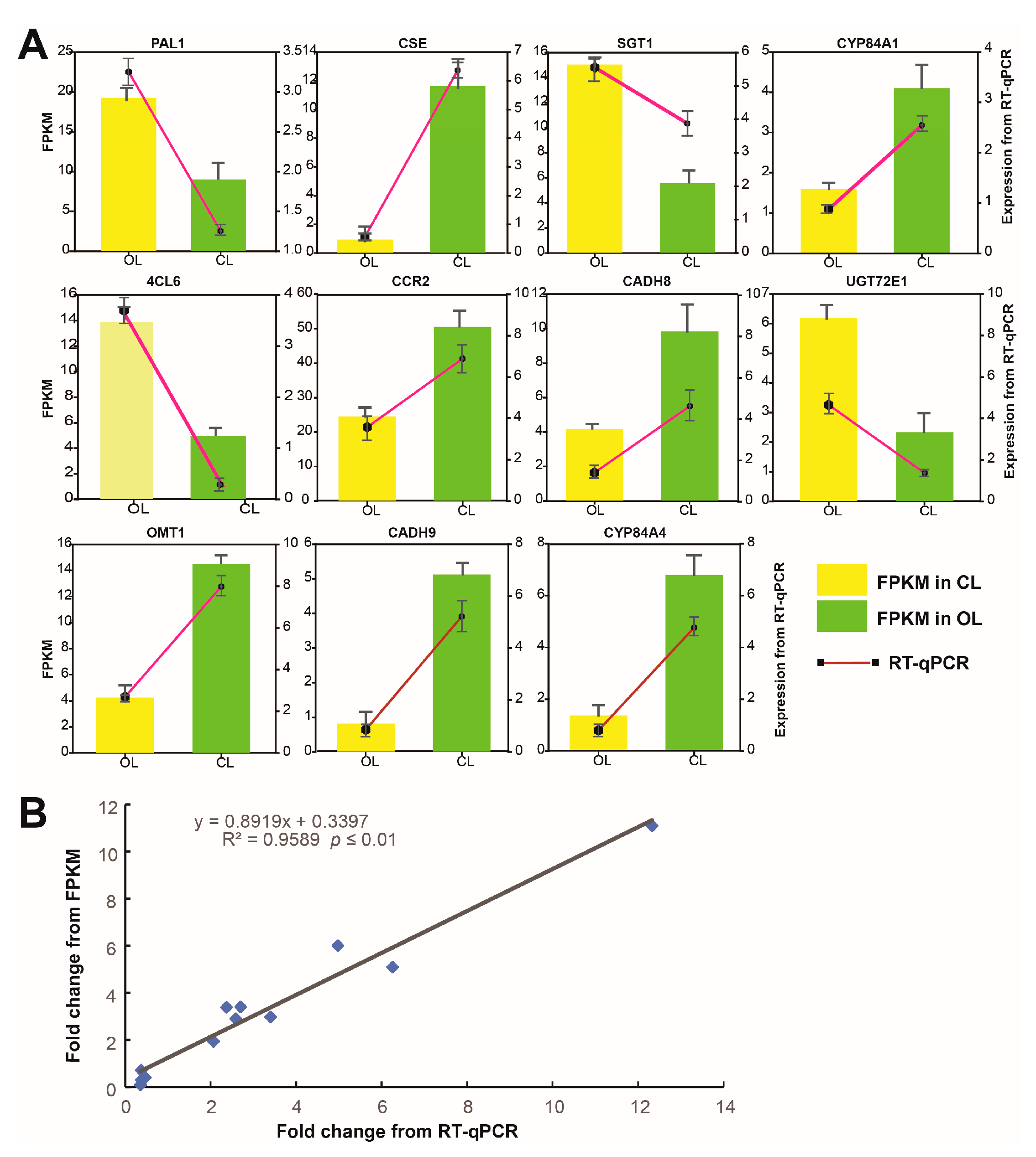{kind=link}
| Database | KEGG | Nr | Swissprot | TrEMBL | KOG | GO | Pfam | Total |
|---|---|---|---|---|---|---|---|---|
| Number | 60,173 | 82,706 | 62,987 | 82,521 | 50,137 | 72,691 | 57,208 | 84,140 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, S.; Cong, X.; Liu, H.; Hu, Y.; Yang, W.; Cheng, H.; Cheng, S.; Zhang, Y. Revealing the Phenolic Acids in Cardamine violifolia Leaves by Transcriptome and Metabolome Analyses. Metabolites 2022, 12, 1024. https://doi.org/10.3390/metabo12111024
Rao S, Cong X, Liu H, Hu Y, Yang W, Cheng H, Cheng S, Zhang Y. Revealing the Phenolic Acids in Cardamine violifolia Leaves by Transcriptome and Metabolome Analyses. Metabolites. 2022; 12(11):1024. https://doi.org/10.3390/metabo12111024
Chicago/Turabian StyleRao, Shen, Xin Cong, Haodong Liu, Yili Hu, Wei Yang, Hua Cheng, Shuiyuan Cheng, and Yue Zhang. 2022. "Revealing the Phenolic Acids in Cardamine violifolia Leaves by Transcriptome and Metabolome Analyses" Metabolites 12, no. 11: 1024. https://doi.org/10.3390/metabo12111024