
Integrated Metabolomic and Transcriptomic Analyses Reveal the Basis for Carotenoid Biosynthesis in Sweet Potato (Ipomoea batatas (L.) Lam.) Storage Roots
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Analyses of Total Carotenoid Content and Color
2.3. Quantification of Carotenoid Content and ABA Levels
2.4. qPCR
2.5. RNA-Sequencing
2.6. Quality Control and Bioinformatics Analyses
2.7. Statistical Analysis
3. Results
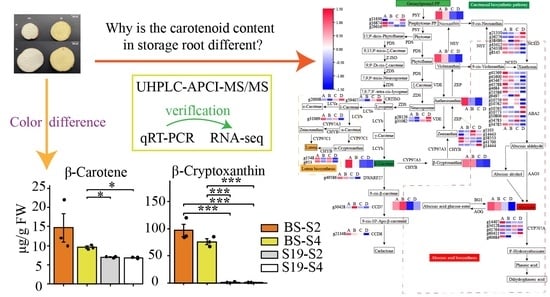
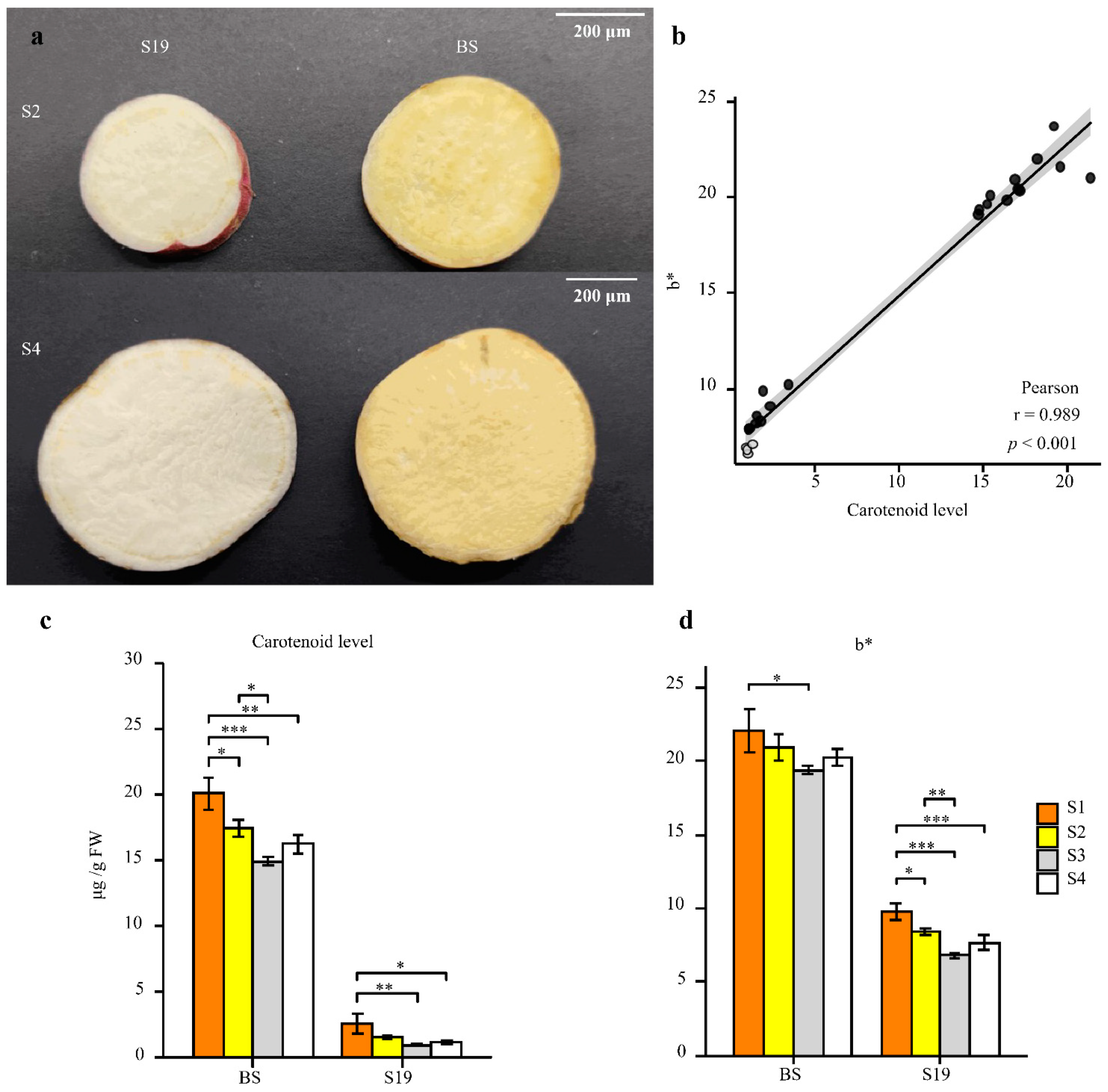
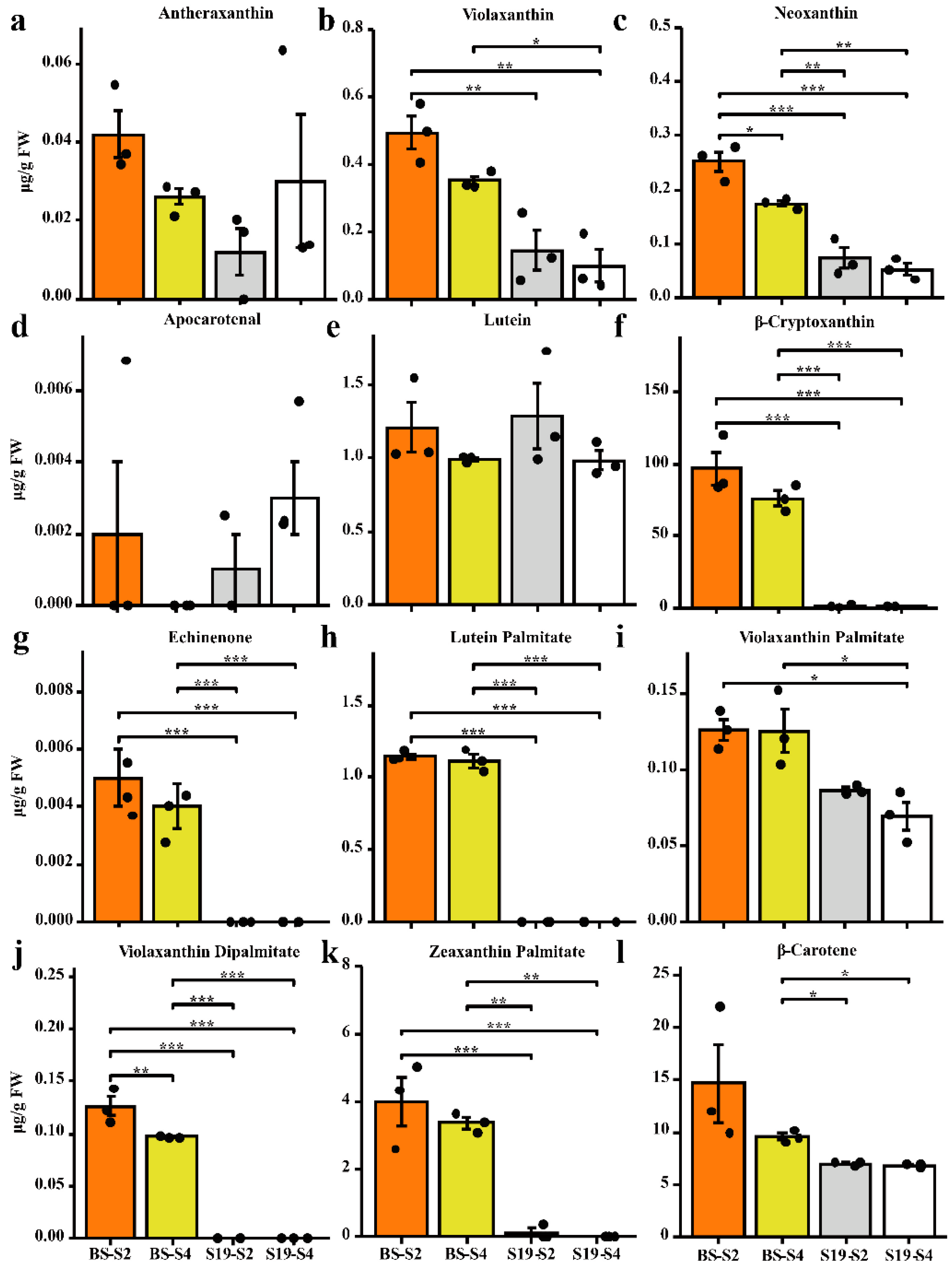
3.1. Carotenoid Accumulation in Storage Roots Varies in Different Sweet Potato Types
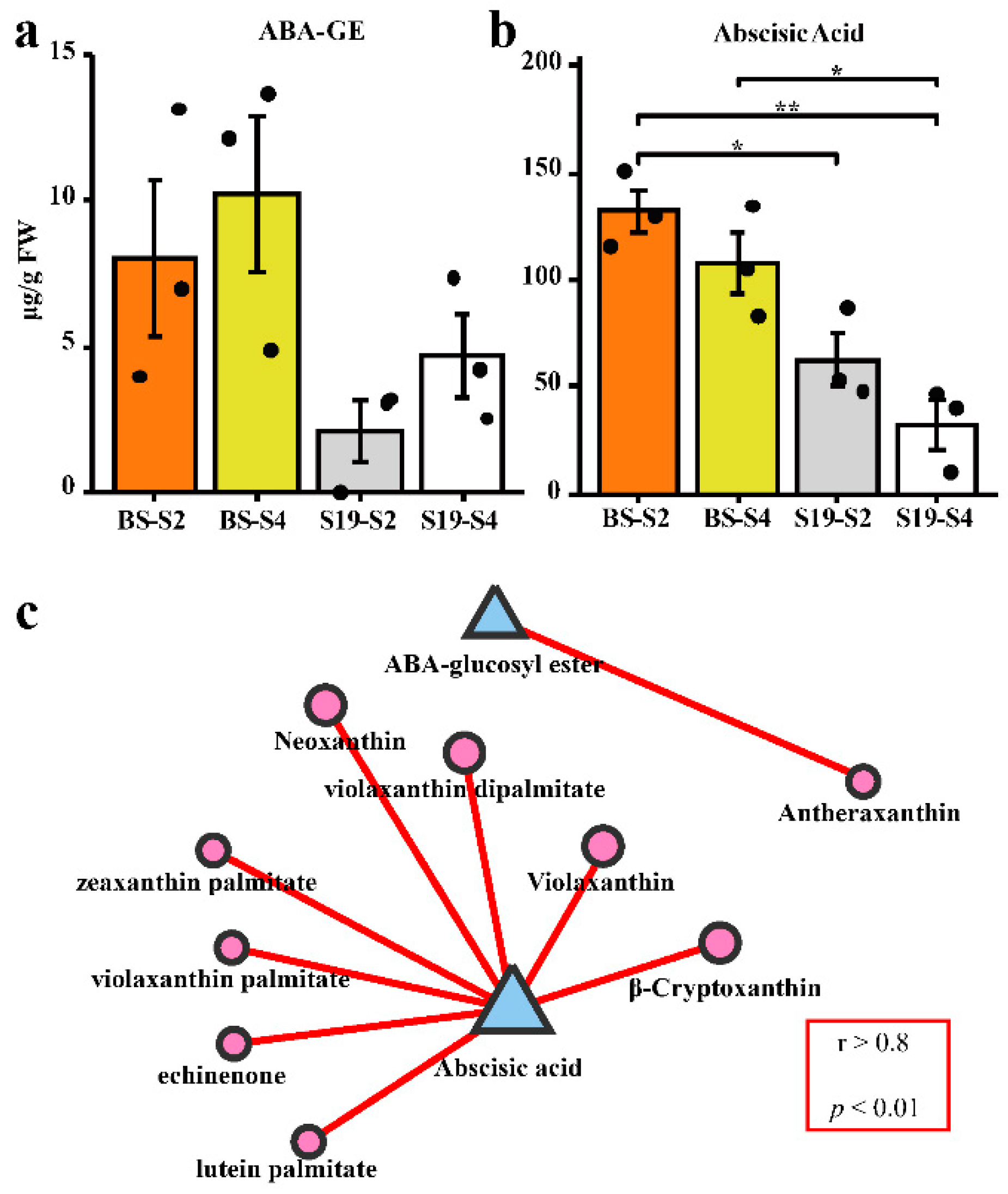
3.2. Changes in ABA Metabolism over the Course of Storage Root Development
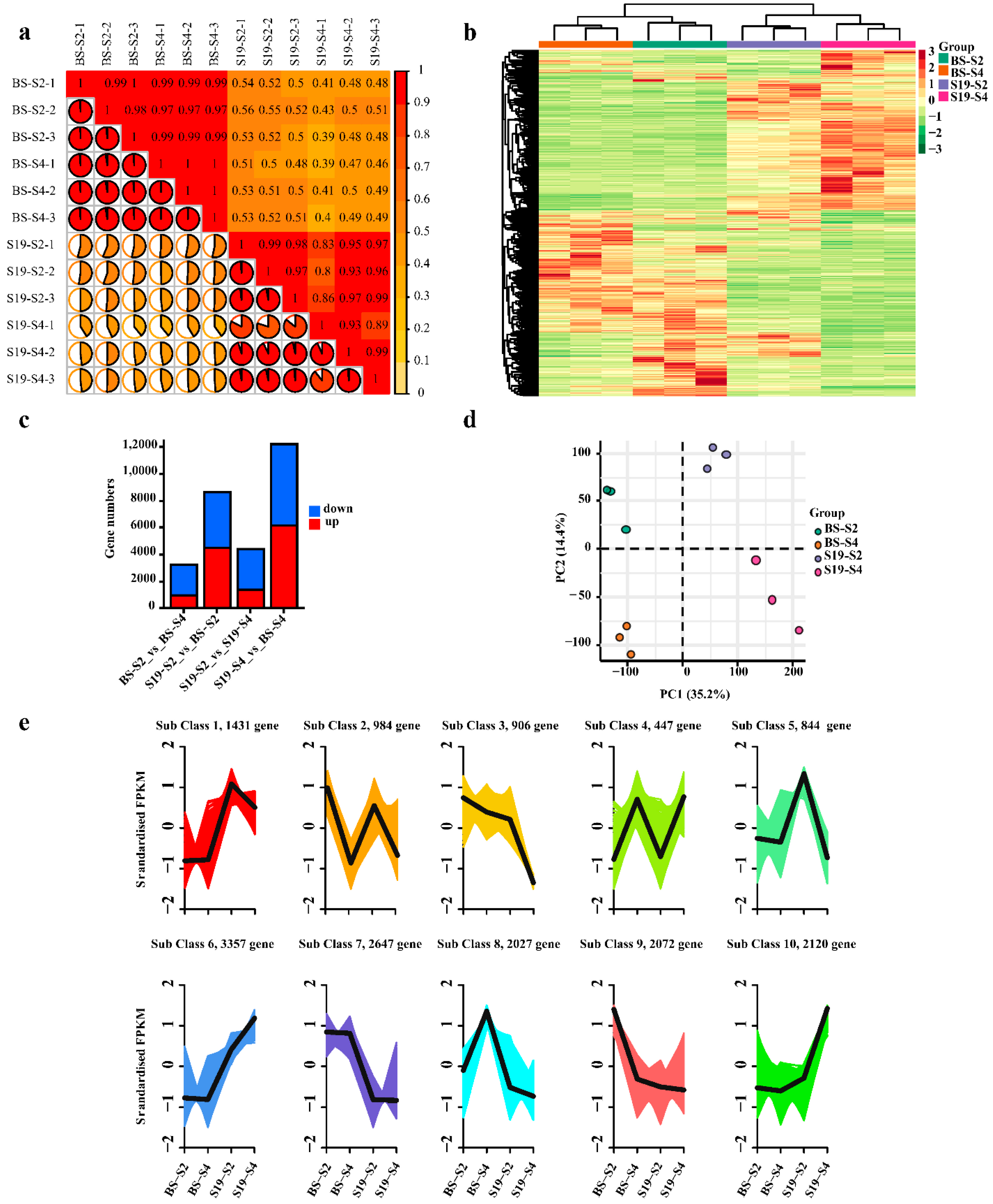
3.3. Transcriptomic Analyses of BS and S19 Sweet Potato Storage Roots
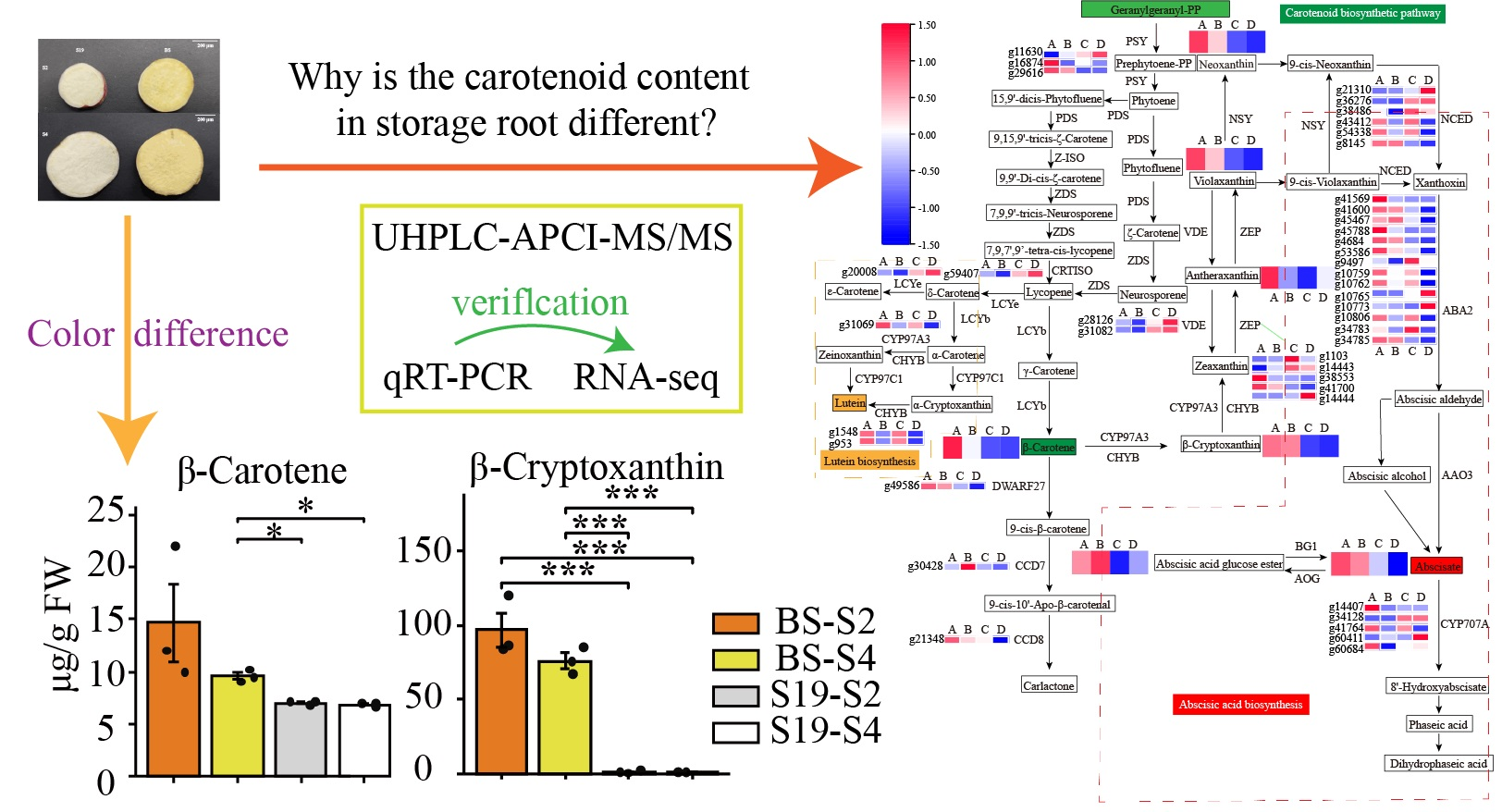
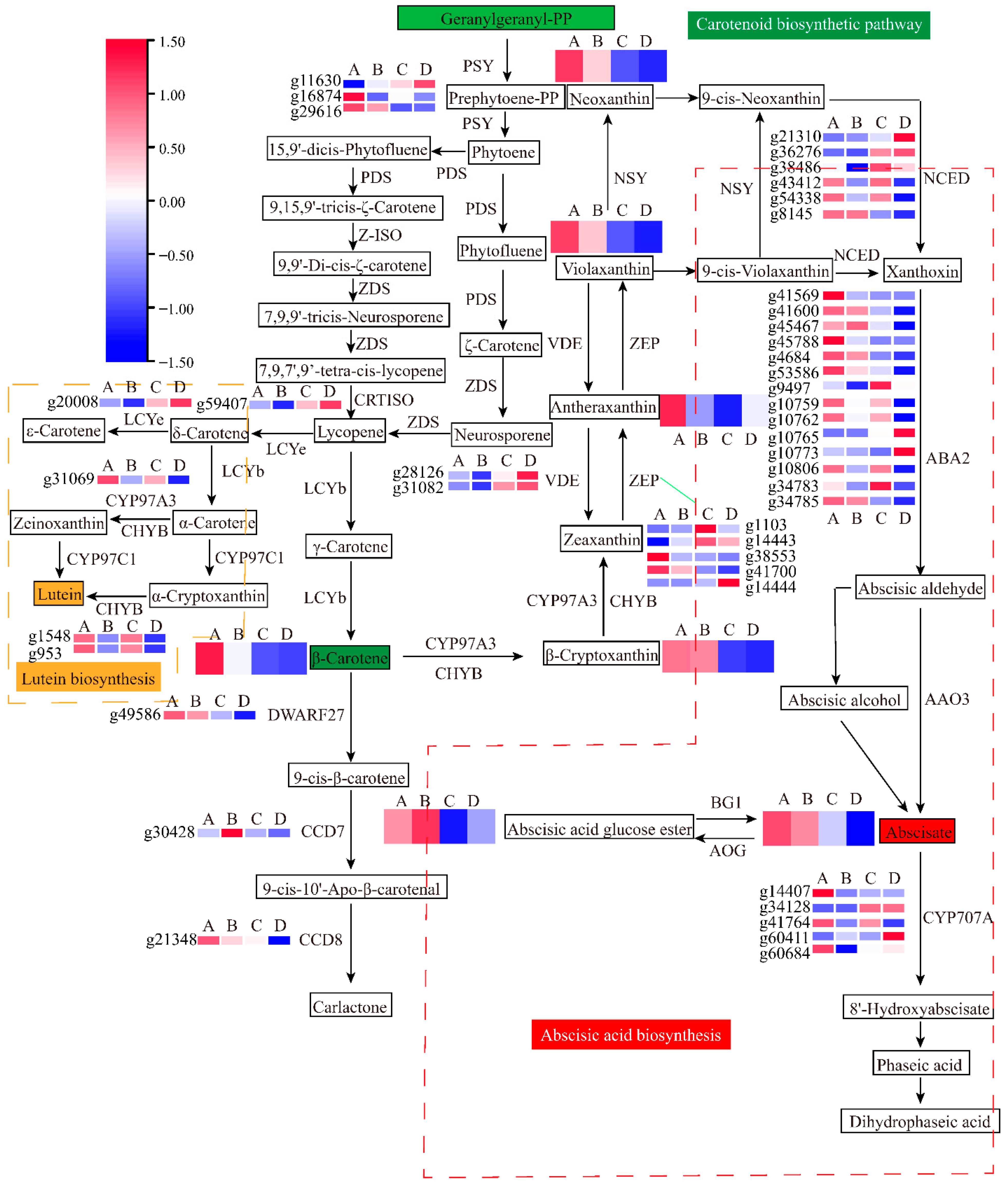
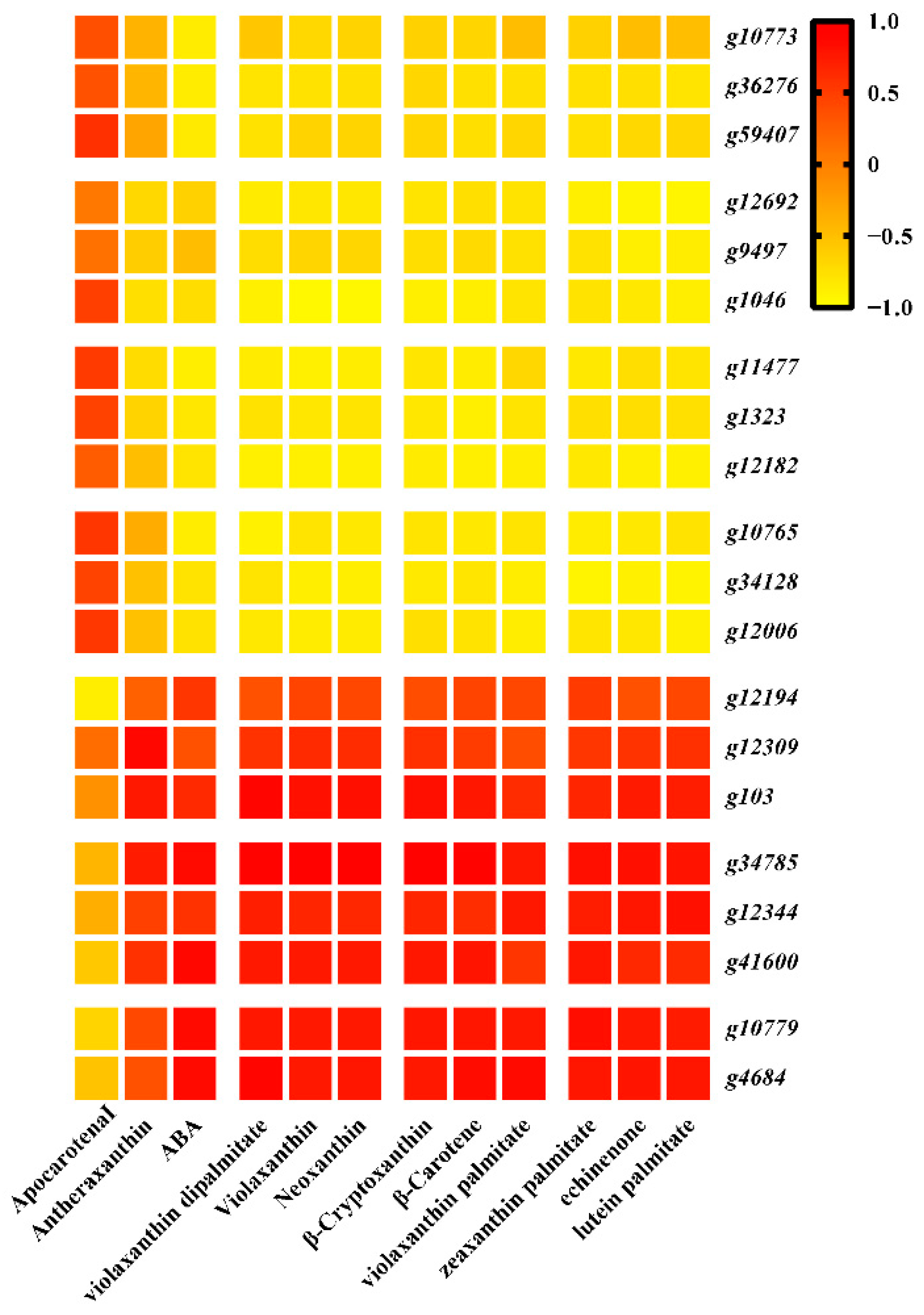
3.4. Combined Analyses of Metabolites and Genes Associated with Carotenoid Biosynthesis in Sweet Potato Storage Roots
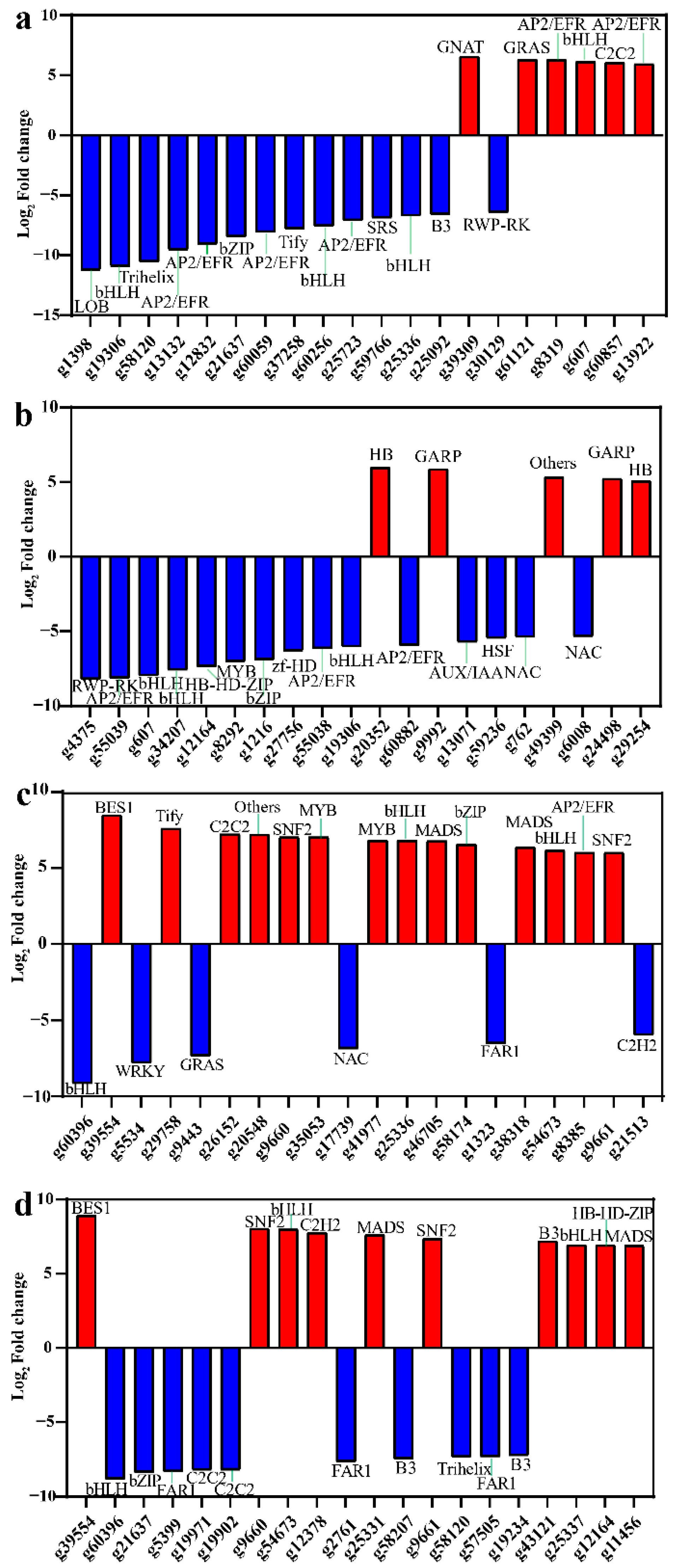
3.5. Identification of Transcription Factors Related to Carotenoid Accumulation
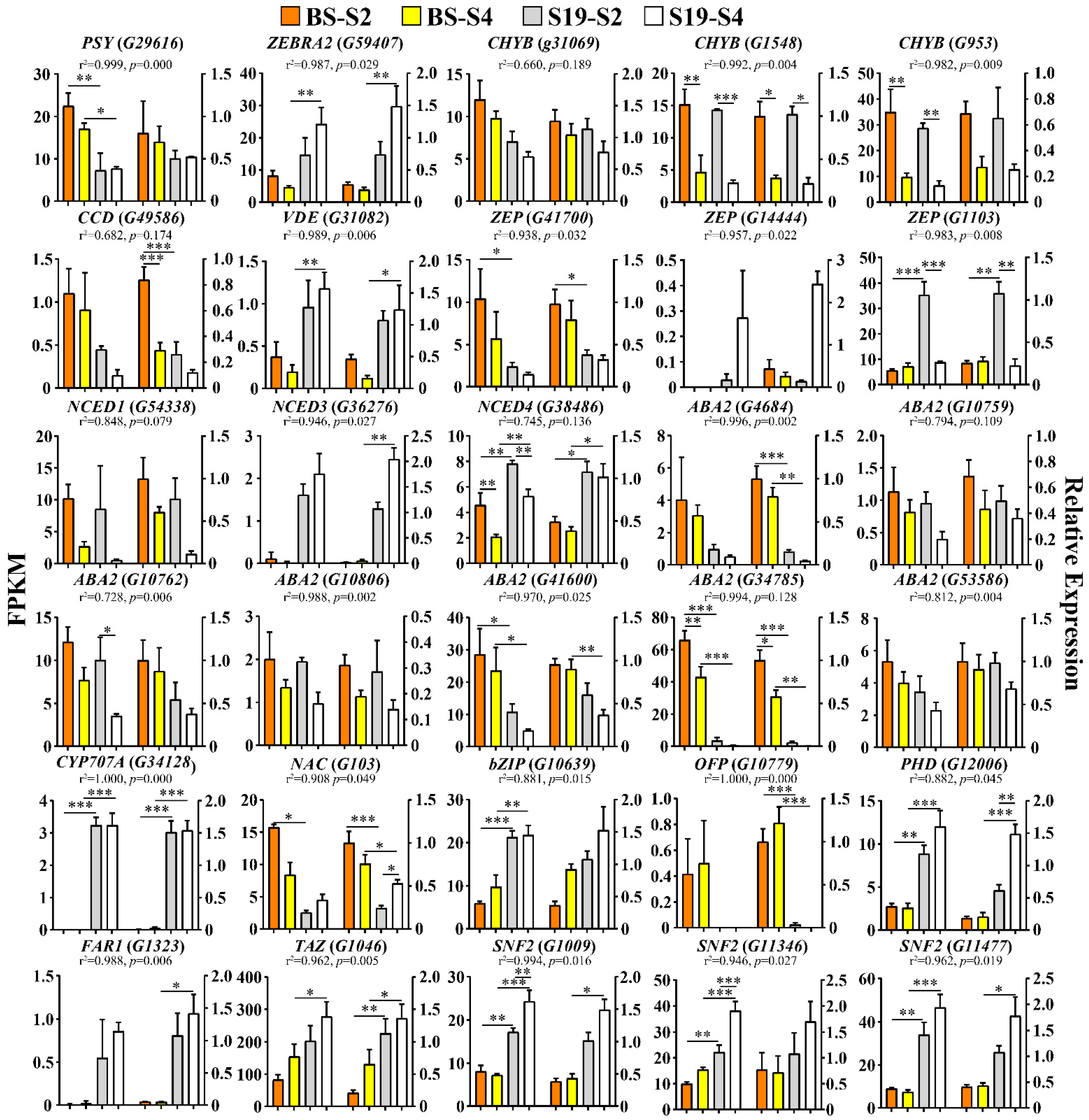
3.6. RNA-Seq Result Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, C.; Kou, M.; Arisha, M.; Tang, W.; Ma, M.; Yan, H.; Wang, X.; Wang, X.; Zhang, Y.; Liu, Y.; et al. Transcriptomic and metabolic profiling of high-temperature treated storage roots reveals the mechanism of saccharification in sweetpotato (Ipomoea batatas (L.) Lam.). Int. J. Mol. Sci. 2021, 22, 6641. [Google Scholar] [CrossRef]
- Banda, L.; Kyallo, M.; Entfellner, J.B.D.; Moyo, M.; Swanckaert, J.; Mwanga, R.O.M.; Onyango, A.; Magiri, E.; Gemenet, D.C.; Yao, N.; et al. Analysis of β-amylase gene (Amyβ) variation reveals allele association with low enzyme activity and increased firmness in cooked sweetpotato (Ipomoea batatas) from East Africa. J. Agric. Food Res. 2021, 4, 100121. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, K.; Zhang, X.; Hayes, M.; Ferruzzi, M.G.; Paton, C.M. Design and nutrient analysis of a carotenoid-rich food product to address vitamin A and protein deficiency. Foods 2021, 10, 1019. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, D.; Mui, L.; Aldridge, E.; McKinney, J.; Hewson, L.; Fisk, I.D. The progression of lipid oxidation, β-carotenes degradation and sensory perception of batch-fried sliced sweet potato crisps during storage. Food Funct. 2021, 12, 4535–4543. [Google Scholar] [CrossRef] [PubMed]
- Quian-Ulloa, R.; Stange, C. Carotenoid biosynthesis and plastid development in plants: The role of light. Int. J. Mol. Sci. 2021, 22, 1184. [Google Scholar] [CrossRef]
- Giannaccare, G.; Pellegrini, M.; Senni, C.; Bernabei, F.; Scorcia, V.; Cicero, A.F.G. Clinical applications of astaxanthin in the treatment of ocular diseases: Emerging insights. Mar. Drugs 2020, 18, 239. [Google Scholar] [CrossRef]
- Ribeiro, D.; Sousa, A.; Nicola, P.; de Oliveira, J.M.P.F.; Rufino, A.T.; Silva, M.; Freitas, M.; Carvalho, F.; Fernandes, E. β-Carotene and its physiological metabolites: Effects on oxidative status regulation and genotoxicity in in vitro models. Food Chem. Toxicol. 2020, 141, 111392. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Liu, X.; Liu, S.; Zhang, J.; Zhang, Y.; Sun, Y.; Tang, R.; Wang, W.; Cui, H.; Li, R.; et al. Transcriptomic and targeted metabolomic analysis identifies genes and metabolites involved in anthocyanin accumulation in tuberous roots of sweetpotato (Ipomoea batatas L.). Plant Physiol. Biochem. 2020, 156, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, J.; Peng, Z.; Shi, M.; Liu, X.; Wen, H.; Jiang, Y.; Cheng, Y.; Xu, J.; Zhang, H. Integrated Transcriptomic and Metabolomic analysis reveals a transcriptional regulation network for the biosynthesis of carotenoids and flavonoids in ‘Cara cara’ navel Orange. BMC Plant Biol. 2021, 21, 29. [Google Scholar] [CrossRef]
- Ishiguro, K.; Yoshinaga, M.; Kai, Y.; Maoka, T.; Yoshimoto, M. Composition, content and antioxidative activity of the carotenoids in yellow-fleshed sweetpotato (Ipomoea batatas L.). Breed. Sci. 2010, 60, 324–329. [Google Scholar] [CrossRef]
- Drapal, M.; Fraser, P.D. Determination of carotenoids in sweet potato (Ipomoea batatas L. Lam) tubers: Implications for accurate provitamin A determination in staple sturdy tuber crops. Phytochemistry 2019, 167, 112102. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Shi, D.; Wang, Y.; Zhang, L.; Chen, X.; Yang, X.; Xiong, H.; Bhattarai, G.; Ravelombola, W.; Olaoye, D.; et al. Transcript profiling for regulation of sweet potato skin color in Sushu8 and its mutant Zhengshu20. Plant Physiol. Biochem. 2020, 148, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Pawlowski, E.G.; Yang, Y.; Sun, T.; Thannhauser, T.W.; Mazourek, M.; Schnell, D.; Li, L. Arabidopsis ORANGE protein regulates plastid pre-protein import through interacting with Tic proteins. J. Exp. Bot. 2021, 72, 1059–1072. [Google Scholar] [CrossRef] [PubMed]
- Barja, M.V.; Ezquerro, M.; Beretta, S.; Diretto, G.; Florez-Sarasa, I.; Feixes, E.; Fiore, A.; Karlova, R.; Fernie, A.R.; Beekwilder, J.; et al. Several geranylgeranyl diphosphate synthase isoforms supply metabolic substrates for carotenoid biosynthesis in tomato. New Phytol. 2021, 231, 255–272. [Google Scholar] [CrossRef]
- Wei, X.; Meng, C.; Yuan, Y.; Nath, U.K.; Zhao, Y.; Wang, Z.; Yang, S.; Li, L.; Niu, L.; Yao, Q.; et al. CaPSY1 gene plays likely the key role in carotenoid metabolism of pepper (Capsicum annuum) at ripening. Funct. Plant. Biol. 2021, 48, 141–155. [Google Scholar] [CrossRef]
- Mallikarjuna Swamy, B.P.; Jr Marundan, S.; Samia, M.; Ordonio, R.L.; Rebong, D.B.; Miranda, R.; Alibuyog, A.; Rebong, A.T.; Tabil, M.A.; Suralta, R.R.; et al. Development and characterization of GR2E Golden rice introgression lines. Sci. Rep. 2021, 11, 2496. [Google Scholar] [CrossRef] [PubMed]
- Diepenbrock, C.H.; Ilut, D.C.; Magallanes-Lundback, M.; Kandianis, C.B.; Lipka, A.E.; Bradbury, P.J.; Holland, J.B.; Hamilton, J.P.; Wooldridge, E.; Vaillancourt, B.; et al. Eleven biosynthetic genes explain the majority of natural variation in carotenoid levels in maize grain. Plant Cell 2021, 33, 882–900. [Google Scholar] [CrossRef] [PubMed]
- Naing, A.H.; Kyu, S.Y.; Win Pe, P.P.; Il Park, K.; Lee, J.M.; Lim, K.B.; Kim, C.K. Silencing of the phytoene desaturase (PDS) gene affects the expression of fruit-ripening genes in tomatoes. Plant Methods 2019, 15, 110. [Google Scholar] [CrossRef]
- Kössler, S.; Armarego-Marriott, T.; Tarkowská, D.; Turečková, V.; Agrawal, S.; Mi, J.; de Souza, L.P.; Schöttler, M.A.; Schadach, A.; Fröhlich, A.; et al. Lycopene β-cyclase expression influences plant physiology, development, and metabolism in tobacco plants. J. Exp. Bot. 2021, 72, 2544–2569. [Google Scholar] [CrossRef]
- Ding, X.; Jia, L.L.; Xing, G.M.; Tao, J.P.; Sun, S.; Tan, G.F.; Li, S.; Liu, J.X.; Duan, A.Q.; Wang, H.; et al. The accumulation of lutein and β-carotene and transcript profiling of genes related to carotenoids biosynthesis in yellow celery. Mol. Biotechnol. 2021, 63, 638–649. [Google Scholar] [CrossRef]
- Lana, G.; Zacarias-Garcia, J.; Distefano, G.; Gentile, A.; Rodrigo, M.J.; Zacarias, L. Transcriptional analysis of carotenoids accumulation and metabolism in a pink-fleshed lemon mutant. Genes 2020, 11, 1294. [Google Scholar] [CrossRef]
- Chayut, N.; Yuan, H.; Saar, Y.; Zheng, Y.; Sun, T.; Zhou, X.; Hermanns, A.; Elad, O.; Faigenboim, A.; Hui, M.; et al. Comparative transcriptome analyses shed light on carotenoid production and plastid development in melon fruit. Hortic. Res. 2021, 8, 112. [Google Scholar] [CrossRef]
- Felemban, A.; Braguy, J.; Zurbriggen, M.D.; Al-Babili, S. Apocarotenoids involved in plant development and stress response. Front. Plant Sci. 2019, 10, 1168. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.C.; Mi, J.; Alagoz, Y.; Al-Babili, S. Plant apocarotenoids: From retrograde signaling to interspecific communication. Plant J. 2021, 105, 351–375. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, L.; Wang, J.; Wang, R.; Duan, M.; Qiao, C.; Chen, X.; Ma, G.; Zhou, X.; Zhu, M.; Jing, F.; et al. Comparative transcriptomic and metabolomic analyses of carotenoid biosynthesis reveal the basis of white petal color in Brassica napus. Planta 2021, 253, 8. [Google Scholar] [CrossRef] [PubMed]
- Tomlekova, N.; Spasova-Apostolova, V.; Pantchev, I.; Sarsu, F. Mutation associated with orange fruit color increases concentrations of β-carotene in a sweet pepper variety (Capsicum annuum L.). Foods 2021, 10, 1225. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Chen, H.; Wang, J.; Zheng, Y.; Li, Y.; Jin, Z.; Li, J. Joint transcriptomic and metabolic analysis of flavonoids in cyclocarya paliurus leaves. ACS Omega 2021, 6, 9028–9038. [Google Scholar] [CrossRef]
- Zhao, D.; Zhao, L.; Liu, Y.; Zhang, A.; Xiao, S.; Dai, X.; Yuan, R.; Zhou, Z.; Cao, Q. Metabolomic and transcriptomic analyses of the flavonoid biosynthetic pathway for the accumulation of anthocyanins and other flavonoids in sweetpotato root skin and leaf vein base. J. Agric. Food Chem. 2022, 70, 2574–2588. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, A.; Nakamura, K.; Sakata, K.; Sato-Fukuda, N.; Ishigaki, T.; Mano, J.; Takabatake, R.; Kitta, K.; Teshima, R.; Kondo, K.; et al. Development and interlaboratory validation of a simple screening method for genetically modified maize using a ΔΔC(q)-based multiplex real-time PCR assay. Anal. Chem. 2016, 88, 4285–4293. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.N.; Nusrat, T.; Begum, P.; Ahsan, M. Carotenoids and β-carotene in orange fleshed sweet potato: A possible solution to vitamin A deficiency. Food Chem. 2016, 199, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Park, S.C.; Ji, C.Y.; Kim, H.S.; Lee, H.S.; Kwak, S.S. Metabolic engineering of carotenoids in transgenic sweetpotato. Breed. Sci. 2017, 67, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, D.; Arenas, M.A.; Flores-Ortiz, C.; Peirano, C.; Handford, M.; Stange, C. Daucus carota DcPSY2 and DcLCYB1 as tools for carotenoid metabolic engineering to improve the nutritional value of fruits. Front. Plant Sci. 2021, 12, 677553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Tao, N.; Xu, Q.; Zhou, W.; Cao, H.; Xu, J.; Deng, X. Functional characterization of Citrus PSY gene in Hongkong kumquat (Fortunella hindsii Swingle). Plant Cell Rep. 2009, 28, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Villalón, A.; Gas, E.; Rodríguez-Concepción, M. Phytoene synthase activity controls the biosynthesis of carotenoids and the supply of their metabolic precursors in dark-grown Arabidopsis seedlings. Plant J. 2009, 60, 424–435. [Google Scholar] [CrossRef]
- Yamamizo, C.; Kishimoto, S.; Ohmiya, A. Carotenoid composition and carotenogenic gene expression during Ipomoea petal development. J. Exp. Bot. 2009, 61, 709–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suematsu, K.; Tanaka, M.; Kurata, R.; Kai, Y. Comparative transcriptome analysis implied a ZEP paralog was a key gene involved in carotenoid accumulation in yellow-fleshed sweetpotato. Sci. Rep. 2020, 10, 20607. [Google Scholar] [CrossRef] [PubMed]
- Chauffour, F.; Bailly, M.; Perreau, F.; Cueff, G.; Suzuki, H.; Collet, B.; Frey, A.; Clément, G.; Soubigou-Taconnat, L.; Balliau, T.; et al. Multi-omics analysis reveals sequential roles for ABA during seed maturation. Plant Physiol. 2019, 180, 1198–1218. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Zhang, J.; Wang, S.; Yang, L.; Luo, Y.; Gao, S.; Zhang, M.; Wu, S.; Hu, S.; Sun, H.; et al. The apricot (Prunus armeniaca L.) genome elucidates Rosaceae evolution and beta-carotenoid synthesis. Hortic. Res. 2019, 6, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Zhao, S.; Xu, M.; Niu, Y.; Nasier, M.; Fan, G.; Quan, S.; Zhang, S.; Wang, Y.; Liao, K. Identification of key genes controlling carotenoid metabolism during apricot fruit development by integrating metabolic phenotypes and gene expression profiles. J. Agric. Food Chem. 2021, 69, 9472–9483. [Google Scholar] [CrossRef]
- González-Guzmán, M.; Apostolova, N.; Bellés, J.M.; Piqueras, P.; Ponce, M.R.; Micol, J.L.; Serrano, R.; Rodríguez, P.L. The short-chain alcohol dehydrogenase ABA2 catalyzes the conversion of xanthoxin to abscisic aldehyde. Plant Cell 2002, 14, 1833–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.M.; Park, S.H.; Ma, S.H.; Park, S.Y.; Yun, C.H.; Jang, G.; Joung, Y.H. Promoted ABA hydroxylation by capsicum annuum CYP707As overexpression suppresses pollen maturation in nicotiana tabacum. Front. Plant Sci. 2020, 11, 583767. [Google Scholar] [CrossRef]
- Dhar, M.K.; Sharma, R.; Koul, A.; Kaul, S. Development of fruit color in Solanaceae: A story of two biosynthetic pathways. Brief. Funct. Genom. 2014, 14, 199–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, F.; Cai, J.H.; Kong, X.M.; Zhou, Q.; Zhou, X.; Zhao, Y.B.; Ji, S.J. Transcriptome profiling reveals the roles of pigment mechanisms in postharvest broccoli yellowing. Hortic. Res. 2019, 6, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Roof, S.; Ye, Z.; Barry, C.; van Tuinen, A.; Vrebalov, J.; Bowler, C.; Giovannoni, J. Manipulation of light signal transduction as a means of modifying fruit nutritional quality in tomato. Proc. Nati. Acad. Sci. USA 2004, 101, 9897–9902. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Guo, C.; Shi, H.; Zhao, J.; Ma, F.; An, W.; He, X.; Luo, Q.; Cao, Y.; Zhan, X. Genome-wide comparative analysis of the R2R3-MYB gene family in five solanaceae species and identification of members regulating carotenoid biosynthesis in wolfberry. Int. J. Mol. Sci. 2022, 23, 2259. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Ji, Q.; Huang, Y.; Jiang, Z.; Bao, M.; Wang, H.; Lin, R. FAR-RED ELONGATED HYPOCOTYL3 and FAR-RED IMPAIRED RESPONSE1 transcription factors integrate light and abscisic acid signaling in Arabidopsis. Plant Physiol. 2013, 163, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, D.; Zhou, X.; Zhou, G.; Zong, W.; Chen, L.; Chang, Y.; Wu, X. Transcription factor AtOFP1 involved in ABA-mediated seed germination and root growth through modulation of ROS homeostasis in arabidopsis. Int. J. Mol. Sci. 2022, 23, 7427. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chang, Y.; Guo, J.; Zeng, Q.; Ellis, B.E.; Chen, J.G. Arabidopsis ovate family proteins, a novel transcriptional repressor family, control multiple aspects of plant growth and development. PLoS ONE 2011, 6, e23896. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Q.; Zhen, X.; Gao, H.; Liang, Y.; Li, H.; Zhao, J.; Yin, M.; Han, Y.; Zhang, B. Integrated Metabolomic and Transcriptomic Analyses Reveal the Basis for Carotenoid Biosynthesis in Sweet Potato (Ipomoea batatas (L.) Lam.) Storage Roots. Metabolites 2022, 12, 1010. https://doi.org/10.3390/metabo12111010
Ren Q, Zhen X, Gao H, Liang Y, Li H, Zhao J, Yin M, Han Y, Zhang B. Integrated Metabolomic and Transcriptomic Analyses Reveal the Basis for Carotenoid Biosynthesis in Sweet Potato (Ipomoea batatas (L.) Lam.) Storage Roots. Metabolites. 2022; 12(11):1010. https://doi.org/10.3390/metabo12111010
Chicago/Turabian StyleRen, Qingming, Xiaoxi Zhen, Huiyu Gao, Yinpei Liang, Hongying Li, Juan Zhao, Meiqiang Yin, Yuanhuai Han, and Bin Zhang. 2022. "Integrated Metabolomic and Transcriptomic Analyses Reveal the Basis for Carotenoid Biosynthesis in Sweet Potato (Ipomoea batatas (L.) Lam.) Storage Roots" Metabolites 12, no. 11: 1010. https://doi.org/10.3390/metabo12111010