

Esterified Oxylipins: Do They Matter?


Abstract
:1. Introduction
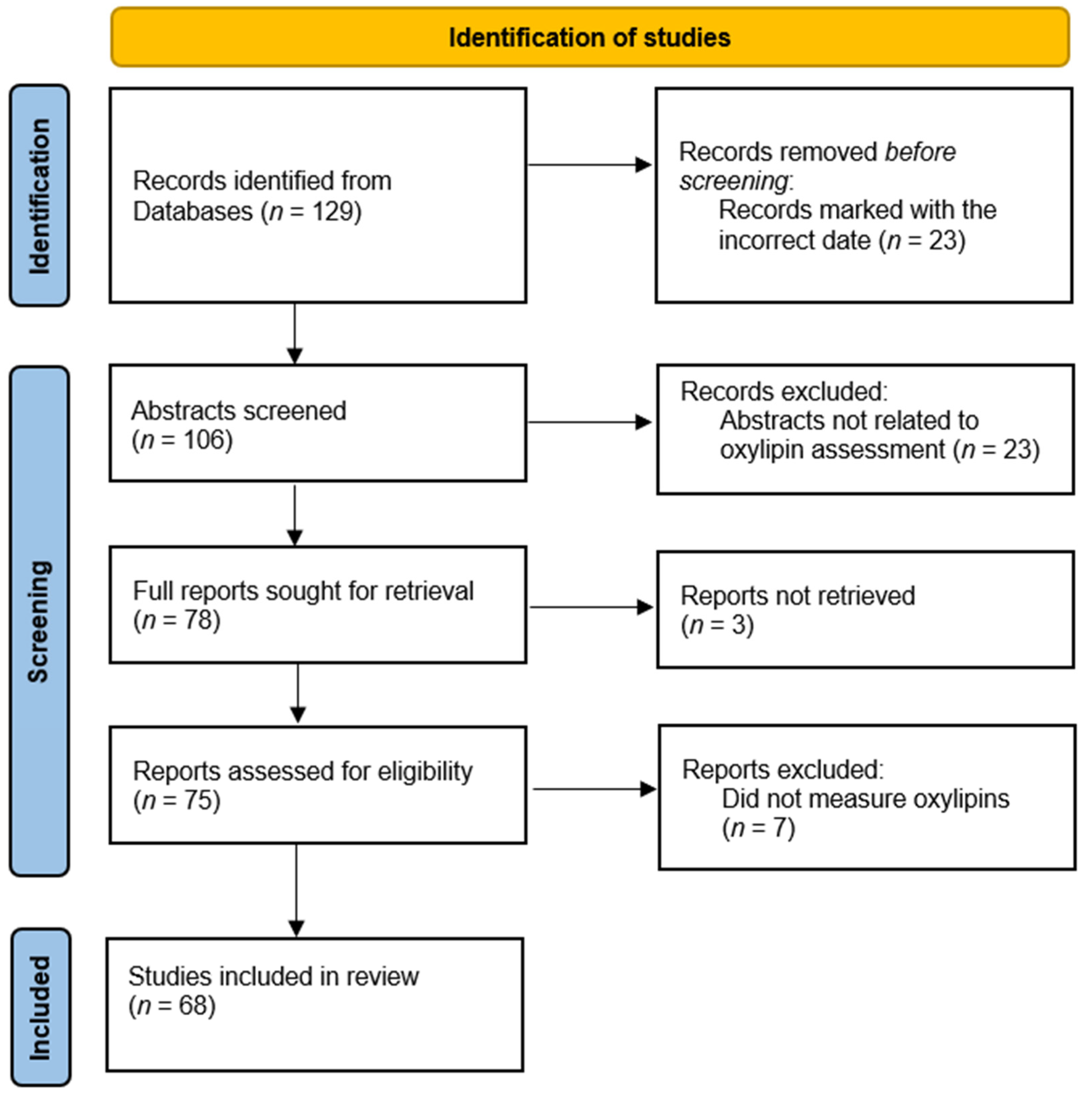
2. Current Practice in Oxylipin Measurements
3. Esterified Oxylipins in Plasma & Implications
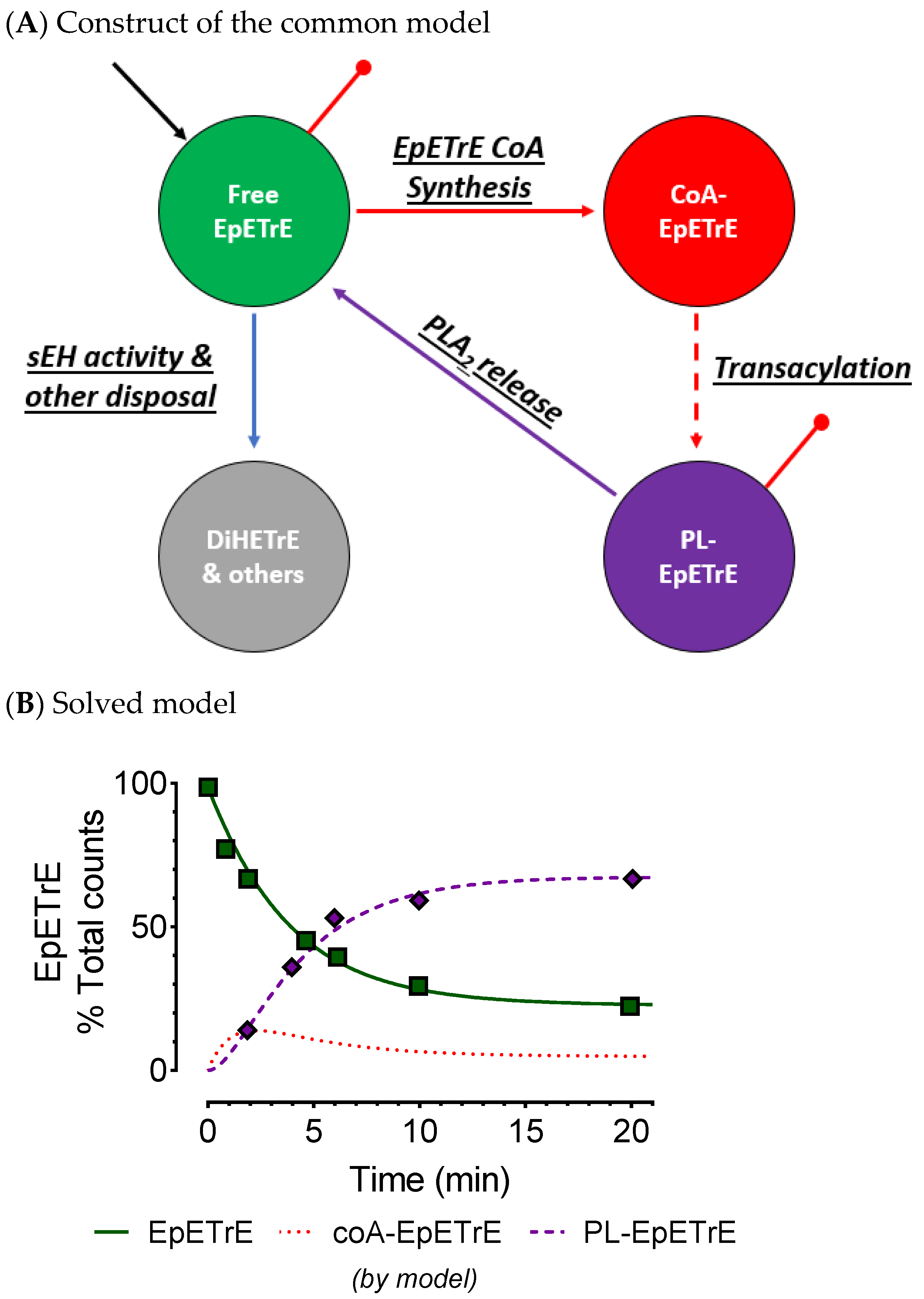
4. Esterified Oxylipins in Cells & Implications
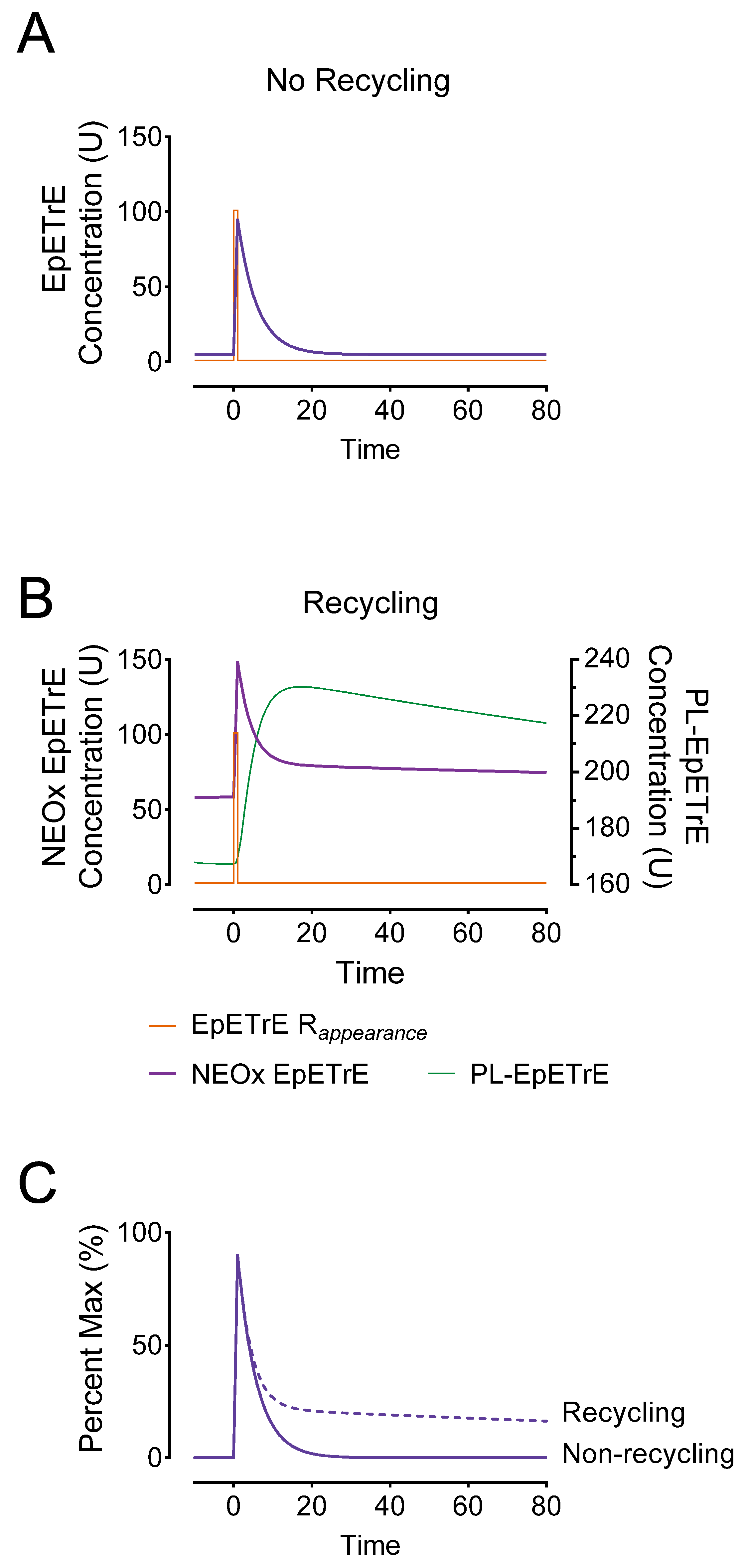
- Steady-state: Compared to a non-recycling system, the basal steady-state concentrations in recycling system producing 1 U/min EpETrE were much greater: 5.8 U vs. 58 U. This drastic difference is largely a function of the permanent disposal, and indicates that in cells undergoing basal oxylipin synthesis, steady state levels are a function of net disposal (e.g., sEH activity), not recycling.
- Rapid (but incomplete) signal termination: Recycling allows for rapid termination of most EpETrE activity in the cytosol. In the non-recycling systems where all disappearance is by cytosolic degradation, the half-life is 3.3 min with the signal nearly 50% terminated by 14 min; however, in the recycling system, the initial rapid half-life is identical and the signal is 50% terminated at nearly the same time.
- Signal extension: Despite rapid initial termination, signal duration in the recycling system is paradoxically extended. This is because most EpETrE disappearance is re-acylation, not sEH-mediated hydrolysis. The half-life for disposal of EpETrE by sEH hydrolysis is 34 min. After 10 min recycling becomes the dominant process, extending the presence of EpETrE in the cytosol so that it takes approximately 235 min until 10% of peak concentration remains, well beyond the modeled timeline.
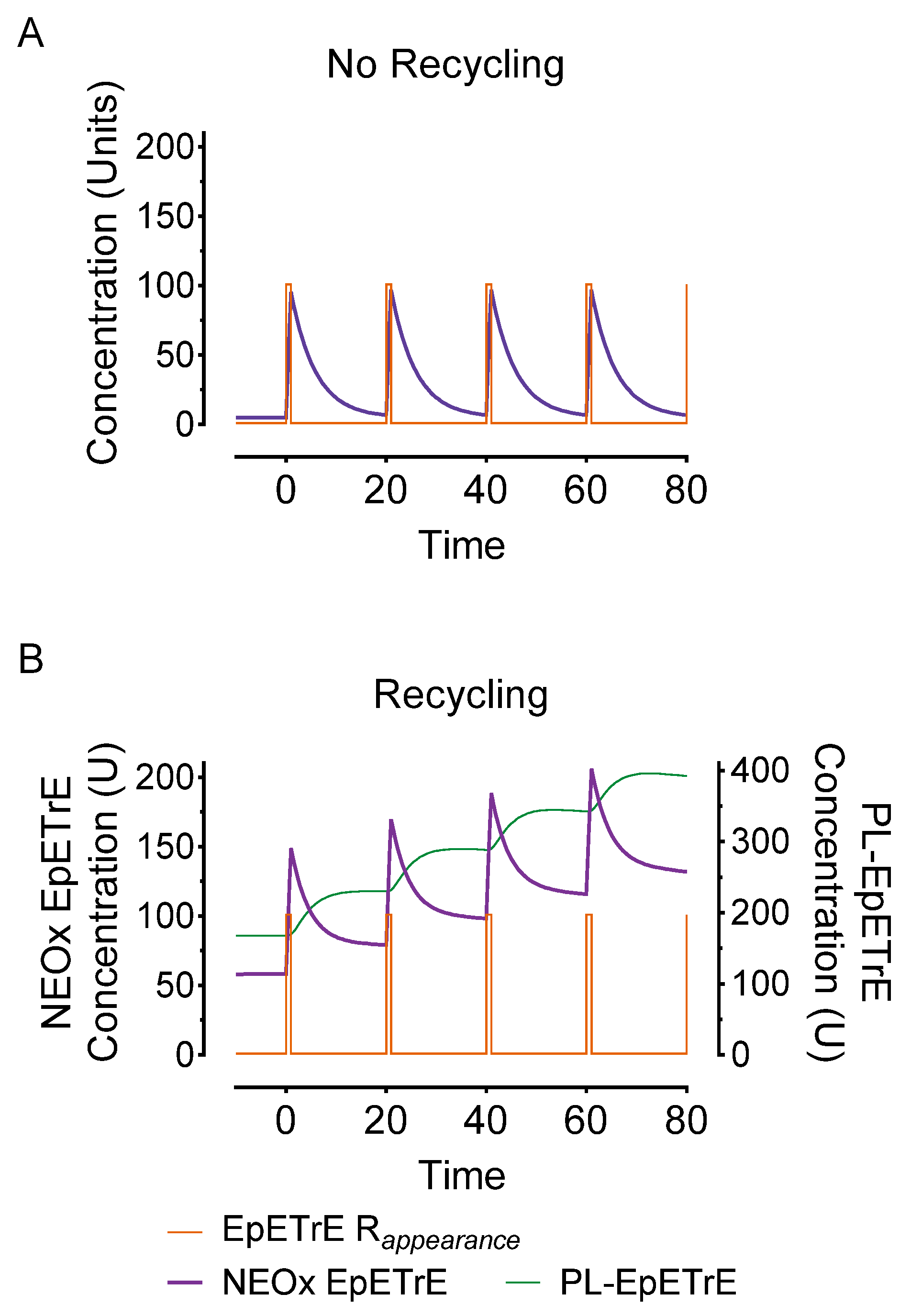
- Stronger signal summation: Finally, recycling would facilitate signal summation. Successive stimuli of EpETrE production would summate more rapidly than systems lacking recycling. Figure 4 depicts the same stimulus as in Figure 3 (100 units over 1 min) but repeated every 20-min. Repeated stimuli do not accumulate in the non-recycling system due to rapid, irreversible disposal. However, in the recycling system, the successive stimuli accumulate in the phospholipid membrane and facilitate a consecutively greater and greater amount of EpETrE in the cytosol with each successive stimulus.
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bae, S.S.; Lee, Y.Y.; Shahbazian, A.; Wang, J.; Meriwether, D.; Golub, I.; Oganesian, B.; Dowd, T.; Reddy, S.T.; Charles-Schoeman, C. High- density lipoprotein function is abnormal in idiopathic inflammatory myopathies. Rheumatology 2020, 59, 3515–3525. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-J.; Switzer, C.H.; Kamynina, A.; Charles, R.; Rudyk, O.; Ng, T.; Burgoyne, J.R.; Eaton, P. Complex interrelationships between nitro-alkene-dependent inhibition of soluble epoxide hydrolase, inflammation and tumor growth. Redox Biol. 2019, 29, 101405. [Google Scholar] [CrossRef] [PubMed]
- Cole, R.M.; Puchala, S.; Ke, J.-Y.; Abdel-Rasoul, M.; Harlow, K.; O’Donnell, B.; Bradley, D.; Andridge, R.; Borkowski, K.; Newman, J.W.; et al. Linoleic Acid–Rich Oil Supplementation Increases Total and High-Molecular-Weight Adiponectin and Alters Plasma Oxylipins in Postmenopausal Women with Metabolic Syndrome. Curr. Dev. Nutr. 2020, 4, nzaa136. [Google Scholar] [CrossRef] [PubMed]
- Gollasch, B.; Wu, G.; Dogan, I.; Rothe, M.; Gollasch, M.; Luft, F.C. Effects of hemodialysis on plasma oxylipins. Physiol. Rep. 2020, 8, e14447. [Google Scholar] [CrossRef]
- Gollasch, B.; Wu, G.; Liu, T.; Dogan, I.; Rothe, M.; Gollasch, M.; Luft, F.C. Hemodialysis and erythrocyte epoxy fatty acids. Physiol. Rep. 2020, 8, e14601. [Google Scholar] [CrossRef]
- Hellhake, S.; Meckelmann, S.W.; Empl, M.T.; Rentmeister, K.; Wißdorf, W.; Steinberg, P.; Schmitz, O.J.; Benter, T.; Schebb, N.H. Non-targeted and targeted analysis of oxylipins in combination with charge-switch derivatization by ion mobility high-resolution mass spectrometry. Anal. Bioanal. Chem. 2020, 412, 5743–5757. [Google Scholar] [CrossRef]
- Leung, H.H.; Leung, K.S.; Durand, T.; Galano, J.; Lee, J.C. Measurement of Enzymatic and Nonenzymatic Polyunsaturated Fatty Acid Oxidation Products in Plasma and Urine of Macular Degeneration UsingLC-QTOF-MS/MS. Lipids 2020, 55, 693–706. [Google Scholar] [CrossRef]
- Mainka, M.; Dalle, C.; Pétéra, M.; Dalloux-Chioccioli, J.; Kampschulte, N.; Ostermann, A.I.; Rothe, M.; Bertrand-Michel, J.; Newman, J.W.; Gladine, C.; et al. Harmonized procedures lead to comparable quantification of total oxylipins across laboratories. J. Lipid Res. 2020, 61, 1424–1436. [Google Scholar] [CrossRef]
- Ostermann, A.I.; Koch, E.; Rund, K.M.; Kutzner, L.; Mainka, M.; Schebb, N.H. Targeting esterified oxylipins by LC–MS—Effect of sample preparation on oxylipin pattern. Prostaglandins Other Lipid Mediat. 2019, 146, 106384. [Google Scholar] [CrossRef]
- Rund, K.M.; Nolte, F.; Doricic, J.; Greite, R.; Schott, S.; Lichtinghagen, R.; Gueler, F.; Schebb, N.H. Clinical blood sampling for oxylipin analysis—Effect of storage and pneumatic tube transport of blood on free and total oxylipin profile in human plasma and serum. Analyst 2020, 145, 2378–2388. [Google Scholar] [CrossRef]
- Van Name, M.A.; Savoye, M.; Chick, J.M.; Galuppo, B.T.; Feldstein, A.E.; Pierpont, B.; Johnson, C.; Shabanova, V.; Ekong, U.; Valentino, P.L.; et al. A Low omega-6 to omega-3 PUFA Ratio (n-6:n-3 PUFA) Diet to Treat Fatty Liver Disease in Obese Youth. J. Nutr. 2020, 150, 2314–2321. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Lin, Y.; Wang, X.; Liu, X.; Wang, Y.; Qin, J.; Wang, X.; Day, D.; Xiang, J.; Mo, J.; et al. Associations of ozone exposure with urinary metabolites of arachidonic acid. Environ. Int. 2020, 145, 106154. [Google Scholar] [CrossRef]
- Dróżdż, M.; Gdula-Argasińska, J.; Biesiada, G.; Jurczyszyn, A.; Perucki, W.; Wolkow, P.; Borys, A.; Sroczyńska, K.; Zając, A.; Librowski, T.; et al. Clostridium difficile caused changes in fatty acids profile and resolvin D1 content in plasma of infected patients. Eur. J. Gastroenterol. Hepatol. 2020, 32, 318–324. [Google Scholar] [CrossRef]
- Jung, J.-S.; Kho, A.R.; Lee, S.H.; Choi, B.Y.; Kang, S.-H.; Koh, J.-Y.; Suh, S.W.; Song, D.-K. Changes in plasma lipoxin A4, resolvins and CD59 levels after ischemic and traumatic brain injuries in rats. Korean J. Physiol. Pharmacol. 2020, 24, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.A.; Lawyer, G.; McDonough, S.; Wang, Q.; Kassem, N.O.; Kas-Petrus, F.; Ye, D.; Singh, K.P.; Kassem, N.O.; Rahman, I. Systemic biomarkers of inflammation, oxidative stress and tissue injury and repair among waterpipe, cigarette and dual tobacco smokers. Tob. Control. 2019, 29, s102–s109. [Google Scholar] [CrossRef] [PubMed]
- Miao, T.; Huang, B.; He, N.; Sun, L.; Du, G.; Gong, X.; Xu, Y.; Zheng, Y.; Zheng, H.; Qu, H. Decreased Plasma Maresin 1 Concentration Is Associated with Diabetic Foot Ulcer. Mediat. Inflamm. 2020, 2020, 4539035. [Google Scholar] [CrossRef] [PubMed]
- Monserrat-Mesquida, M.; Quetglas-Llabrés, M.; Abbate, M.; Montemayor, S.; Mascaró, C.; Casares, M.; Tejada, S.; Abete, I.; Zulet, M.; Tur, J.; et al. Oxidative Stress and Pro-Inflammatory Status in Patients with Non-Alcoholic Fatty Liver Disease. Antioxidants 2020, 9, 759. [Google Scholar] [CrossRef]
- Mostafa, D.G.; Satti, H.H. Resolvin D1 Prevents the Impairment in the Retention Memory and Hippocampal Damage in Rats Fed a Corn Oil-Based High Fat Diet by Upregulation of Nrf2 and Downregulation and Inactivation of p66Shc. Neurochem. Res. 2020, 45, 1576–1591. [Google Scholar] [CrossRef] [PubMed]
- Mottola, G.; Werlin, E.C.; Wu, B.; Chen, M.; Chatterjee, A.; Schaller, M.S.; Conte, M.S. Oral Resolvin D1 attenuates early inflammation but not intimal hyperplasia in a rat carotid angioplasty model. Prostaglandins Other Lipid Mediat. 2019, 146, 106401. [Google Scholar] [CrossRef]
- Parashar, K.; Schulte, F.; Hardt, M.; Baker, O.J. Sex-mediated elevation of the specialized pro-resolving lipid mediator levels in a Sjogren’s syndrome mouse model. FASEB J. 2020, 34, 7733–7744. [Google Scholar] [CrossRef] [PubMed]
- Tejera, P.; Abdulnour, R.-E.E.; Zhu, Z.; Su, L.; Levy, B.D.; Christiani, D.C. Plasma Levels of Proresolving and Prophlogistic Lipid Mediators: Association With Severity of Respiratory Failure and Mortality in Acute Respiratory Distress Syndrome. Crit. Care Explor. 2020, 2, e0241. [Google Scholar] [CrossRef]
- Tian, L.X.; Tang, X.; Zhu, J.Y.; Luo, L.; Ma, X.Y.; Cheng, S.W.; Tang, W.-Q.; Ma, W.; Yang, X.; Lv, C.-Z.; et al. Cytochrome P450 1A1 enhances inflammatory responses and impedes phagocytosis of bacteria in macrophages during sepsis. Cell Commun. Signal. 2020, 18, 70. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.; Manke, J.; Nkrumah-Elie, Y.; Shaikh, S.R.; Reisdorph, N. Improved quantification of lipid mediators in plasma and tissues by liquid chromatography tandem mass spectrometry demonstrates mouse strain specific differences. Prostaglandins Other Lipid Mediat. 2020, 151, 106483. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lopez, A.; Godzien, J.; Soldevilla, B.; Gradillas, A.; Lopez-Gonzalvez, A.; Lens-Pardo, A.; La Salvia, A.; Del Carmen Riesco-Martinez, M.; Garcia-Carbonero, R.; Barbas, C. Oxidized lipids in the metabolic profiling of neuroendocrine tumors—Analytical challenges and biological implications. J. Chromatogr. A 2020, 1625, 461233. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. and Chou, R. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Systematic reviews 2021, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hinz, C.; Liggi, S.; Mocciaro, G.; Jung, S.; Induruwa, I.; Pereira, M.; Bryant, C.E.; Meckelmann, S.W.; O’Donnell, V.B.; Farndale, R.W.; et al. A Comprehensive UHPLC Ion Mobility Quadrupole Time-of-Flight Method for Profiling and Quantification of Eicosanoids, Other Oxylipins, and Fatty Acids. Anal. Chem. 2019, 91, 8025–8035. [Google Scholar] [CrossRef] [PubMed]
- Morgan, A.H.; Hammond, V.J.; Morgan, L.; Thomas, C.P.; Tallman, K.A.; Garcia-Diaz, Y.R.; McGuigan, C.; Serpi, M.; Porter, N.A.; Murphy, R.C.; et al. Quantitative assays for esterified oxylipins generated by immune cells. Nat. Protoc. 2010, 5, 1919–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illingworth, D.R. Lipoprotein metabolism. Am. J. Kidney Dis. 1993, 22, 90–97. [Google Scholar] [CrossRef]
- Cho, K.-H. The Current Status of Research on High-Density Lipoproteins (HDL): A Paradigm Shift from HDL Quantity to HDL Quality and HDL Functionality. Int. J. Mol. Sci. 2022, 23, 3967. [Google Scholar] [CrossRef]
- Goldstein, J.L.; Brown, M.S. The LDL receptor. Arter. Thromb. Vasc. Biol. 2009, 29, 431–438. [Google Scholar] [CrossRef]
- Huang, J.-K.; Lee, H.-C. Emerging Evidence of Pathological Roles of Very-Low-Density Lipoprotein (VLDL). Int. J. Mol. Sci. 2022, 23, 4300. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, N.; Ontko, J.A. Interactions between fatty acid synthesis, oxidation, and esterification in the production of triglyceride-rich lipoproteins by the liver. J. Lipid Res. 1984, 25, 831–842. [Google Scholar] [CrossRef]
- Curtiss, L.K.; Valenta, D.T.; Hime, N.J.; Rye, K.A. What is so special about apolipoprotein AI in reverse cholesterol transport? Arter. Thromb. Vasc. Biol. 2006, 26, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Go, G.-W.; Mani, A. Low-density lipoprotein receptor (LDLR) family orchestrates cholesterol homeostasis. Yale J. Biol. Med. 2012, 85, 19–28. [Google Scholar] [PubMed]
- Spector, A.A. Fatty acid binding to plasma albumin. J. Lipid Res. 1975, 16, 165–179. [Google Scholar] [CrossRef]
- Walker, R.E.; Ford, J.L.; Boston, R.C.; Savinova, O.V.; Harris, W.S.; Green, M.H.; Shearer, G.C. Trafficking of nonesterified fatty acids in insulin resistance and relationship to dysglycemia. Am. J. Physiol. Metab. 2020, 318, E392–E404. [Google Scholar] [CrossRef]
- Hue, L.; Taegtmeyer, H. The Randle cycle revisited: A new head for an old hat. Am. J. Physiol. Metab. 2009, 297, E578–E591. [Google Scholar] [CrossRef] [Green Version]
- Gouveia-Figueira, S.; Späth, J.; Zivkovic, A.M.; Nording, M.L. Profiling the Oxylipin and Endocannabinoid Metabolome by UPLC-ESI-MS/MS in Human Plasma to Monitor Postprandial Inflammation. PLoS ONE 2015, 10, e0132042. [Google Scholar] [CrossRef] [Green Version]
- Schebb, N.H.; Ostermann, A.I.; Yang, J.; Hammock, B.D.; Hahn, A.; Schuchardt, J.P. Comparison of the effects of long-chain omega-3 fatty acid supplementation on plasma levels of free and esterified oxylipins. Prostaglandins Other Lipid Mediat. 2014, 113–115, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Shearer, G.C.; Newman, J.W. Lipoprotein lipase releases esterified oxylipins from very low-density lipoproteins. Prostaglandins Leukot. Essent. Fat. Acids 2008, 79, 215–222. [Google Scholar] [CrossRef]
- Newman, J.W.; Kaysen, G.A.; Hammock, B.D.; Shearer, G.C. Proteinuria increases oxylipid concentrations in VLDL and HDL but not LDL particles in the rat. J. Lipid Res. 2007, 48, 1792–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, G.C.; Borkowski, K.; Puumala, S.L.; Harris, W.S.; Pedersen, T.L.; Newman, J.W. Abnormal lipoprotein oxylipins in metabolic syndrome and partial correction by omega-3 fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 2017, 128, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.E.; Savinova, O.V.; Pedersen, T.L.; Newman, J.W.; Shearer, G.C. Effects of inflammation and soluble epoxide hydrolase inhibition on oxylipin composition of very low-density lipoproteins in isolated perfused rat livers. Physiol. Rep. 2021, 9, e14480. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.A.; Harsch, B.A.; Healy, C.L.; Joshi, S.S.; Huang, S.; Walker, R.E.; Wagner, B.M.; Ernste, K.M.; Huang, W.; Block, R.C.; et al. Free fatty acid receptor 4 responds to endogenous fatty acids to protect the heart from pressure overload. Cardiovasc. Res. 2021, 118, 1061–1073. [Google Scholar] [CrossRef]
- Borkowski, K.; Yim, S.J.; Holt, R.R.; Hackman, R.M.; Keen, C.L.; Newman, J.W.; Shearer, G.C. Walnuts change lipoprotein composition suppressing TNFalpha-stimulated cytokine production by diabetic adipocyte. J. Nutr. Biochem. 2019, 68, 51–58. [Google Scholar] [CrossRef]
- Holt, R.R.; Yim, S.J.; Shearer, G.C.; Hackman, R.M.; Djurica, D.; Newman, J.; Shindel, A.W.; Keen, C.L. Effects of short-term walnut consumption on human microvascular function and its relationship to plasma epoxide content. J. Nutr. Biochem. 2015, 26, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.W.; Pedersen, T.L.; Brandenburg, V.R.; Harris, W.S.; Shearer, G.C. Effect of Omega-3 Fatty Acid Ethyl Esters on the Oxylipin Composition of Lipoproteins in Hypertriglyceridemic, Statin-Treated Subjects. PLoS ONE 2014, 9, e111471. [Google Scholar] [CrossRef] [Green Version]
- Sala, A.; Folco, G.; Murphy, R.C. Transcellular biosynthesis of eicosanoids. Pharmacol. Rep. 2010, 62, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Sala, A.; Rossoni, G.; Buccellati, C.; Berti, F.; Folco, G.; Maclouf, J. Formation of sulphidopeptide-leukotrienes by cell-cell interaction causes coronary vasoconstriction in isolated, cell-perfused heart of rabbit. J. Cereb. Blood Flow Metab. 1993, 110, 1206–1212. [Google Scholar] [CrossRef] [Green Version]
- Sala, A.; Aliev, G.M.; Rossoni, G.; Berti, F.; Buccellati, C.; Burnstock, G.; Folco, G.; Maclouf, J. Morphological and functional changes of coronary vasculature caused by transcellular biosynthesis of sulfidopeptide leukotrienes in isolated heart of rabbit. Blood 1996, 87, 1824–1832. [Google Scholar] [CrossRef]
- Ek-Von Mentzer, B.A.; Zhang, F.; Hamilton, J.A. Binding of 13-HODE and 15-HETE to phospholipid bilayers, albumin, and intracellular fatty acid binding proteins. implications for transmembrane and intracellular transport and for protection from lipid peroxidation. J. Biol. Chem. 2001, 276, 15575–15580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulyassy, P.F.; Depner, T.A.; Shearer, G.C. Comparison of binding by concentrated peritoneal dialysate and serum. ASAIO J. 1993, 39, M569–M572. [Google Scholar] [PubMed]
- Karara, A.; Dishman, E.; Jacobson, H.; Falck, J.R.; Capdevila, J.H. Arachidonic acid epoxygenase. Stereochemical analysis of the endogenous epoxyeicosatrienoic acids of human kidney cortex. FEBS Lett. 1990, 268, 227–230. [Google Scholar] [CrossRef] [Green Version]
- Capdevila, J.; Pramanik, B.; Napoli, J.L.; Manna, S.; Falck, J. Arachidonic acid epoxidation: Epoxyeicosatrienoic acids are endogenous constituents of rat liver. Arch. Biochem. Biophys. 1984, 231, 511–517. [Google Scholar] [CrossRef]
- Wu, S.; Moomaw, C.R.; Tomer, K.B.; Falck, J.R.; Zeldin, D.C. Molecular Cloning and Expression of CYP2J2, a Human Cytochrome P450 Arachidonic Acid Epoxygenase Highly Expressed in Heart. J. Biol. Chem. 1996, 271, 3460–3468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Henson, P.M.; Murphy, R.C. Occurrence of Oxidized Metabolites of Arachidonic Acid Esterified to Phospholipids in Murine Lung Tissue. Anal. Biochem. 1998, 262, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Hammond, V.J.; O’Donnell, V.B. Esterified eicosanoids: Generation, characterization and function. Biochim. Biophys. Acta (BBA)-Biomembr. 2011, 1818, 2403–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenson, W.F.; Parker, C.W. 12-L-hydroxy-5,8,10,14-eicosatetraenoic acid, a chemotactic fatty acid, is incorporated into neutrophil phospholipids and triglyceride. Prostaglandins 1979, 18, 285–292. [Google Scholar] [CrossRef]
- O’Donnell, V.B. New appreciation for an old pathway: The Lands Cycle moves into new arenas in health and disease. Biochem. Soc. Trans. 2022, 50, 1–11. [Google Scholar] [CrossRef]
- Karara, A.; Dishman, E.; Falck, J.R.; Capdevila, J.H. Endogenous epoxyeicosatrienoyl-phospholipids. A novel class of cellular glycerolipids containing epoxidized arachidonate moieties. J. Biol. Chem. 1991, 266, 7561–7569. [Google Scholar] [CrossRef]
- Weintraub, N.L.; Fang, X.; Kaduce, T.L.; VanRollins, M.; Chatterjee, P.; Spector, A.A. Potentiation of Endothelium-Dependent Relaxation by Epoxyeicosatrienoic Acids. Circ. Res. 1997, 81, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Brezinski, M.E.; Serhan, C.N. Selective incorporation of (15S)-hydroxyeicosatetraenoic acid in phosphatidylinositol of human neutrophils: Agonist-induced deacylation and transformation of stored hydroxyeicosanoids. Proc. Natl. Acad. Sci. USA 1990, 87, 6248–6252. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hydrolysis | No Hydrolysis | ||
|---|---|---|---|
| Location | ELISA | ||
| Plasma | 10 | 38 | 8 |
| Tissue | 0 | 9 | 1 |
| Cell | 2 | 5 | 1 |
| Urine | 2 | 0 | 0 |
| Total * | 12 | 46 | 10 |
| Enzyme Activity | Phospholipid Data Mean (95%CI) |
|---|---|
| Acyl-CoA Synthesis (pools/min) | 0.19 (0.16, 0.21) |
| Esterification (pools/min) | 0.82 (0.58, 1.06) |
| PLA2 Release(pools/min) | 0.065 (0.048, 0.082) |
| sEH (pools/min) | 0.0064 (−0.0046, 0.0173) |
| Residence Time in Free (min) | 157 |
| Residence Time in coA (min) | 36 |
| Residence Time in PL (min) | 450 |
| Total Time in system (min) | 643 |
| Recycling Number | 29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Annevelink, C.E.; Walker, R.E.; Shearer, G.C. Esterified Oxylipins: Do They Matter? Metabolites 2022, 12, 1007. https://doi.org/10.3390/metabo12111007
Annevelink CE, Walker RE, Shearer GC. Esterified Oxylipins: Do They Matter? Metabolites. 2022; 12(11):1007. https://doi.org/10.3390/metabo12111007
Chicago/Turabian StyleAnnevelink, Carmen E., Rachel E. Walker, and Gregory C. Shearer. 2022. "Esterified Oxylipins: Do They Matter?" Metabolites 12, no. 11: 1007. https://doi.org/10.3390/metabo12111007