
Metabolomic Profiling of Malaysian and New Zealand Honey Using Concatenated NMR and HRMS Datasets


Abstract
:1. Introduction
2. Results
2.1. Biological Activities of Honey Extracts
2.2. Chemical Profiling of Malaysian and New Zealand Honey Based on 1H NMR Experiment
2.3. Chemical Diversity of Malaysian and New Zealand Honey Based on HR-LCMS Data
2.4. Visualising Diversity between Malaysian and New Zealand Honey Using Heatmaps
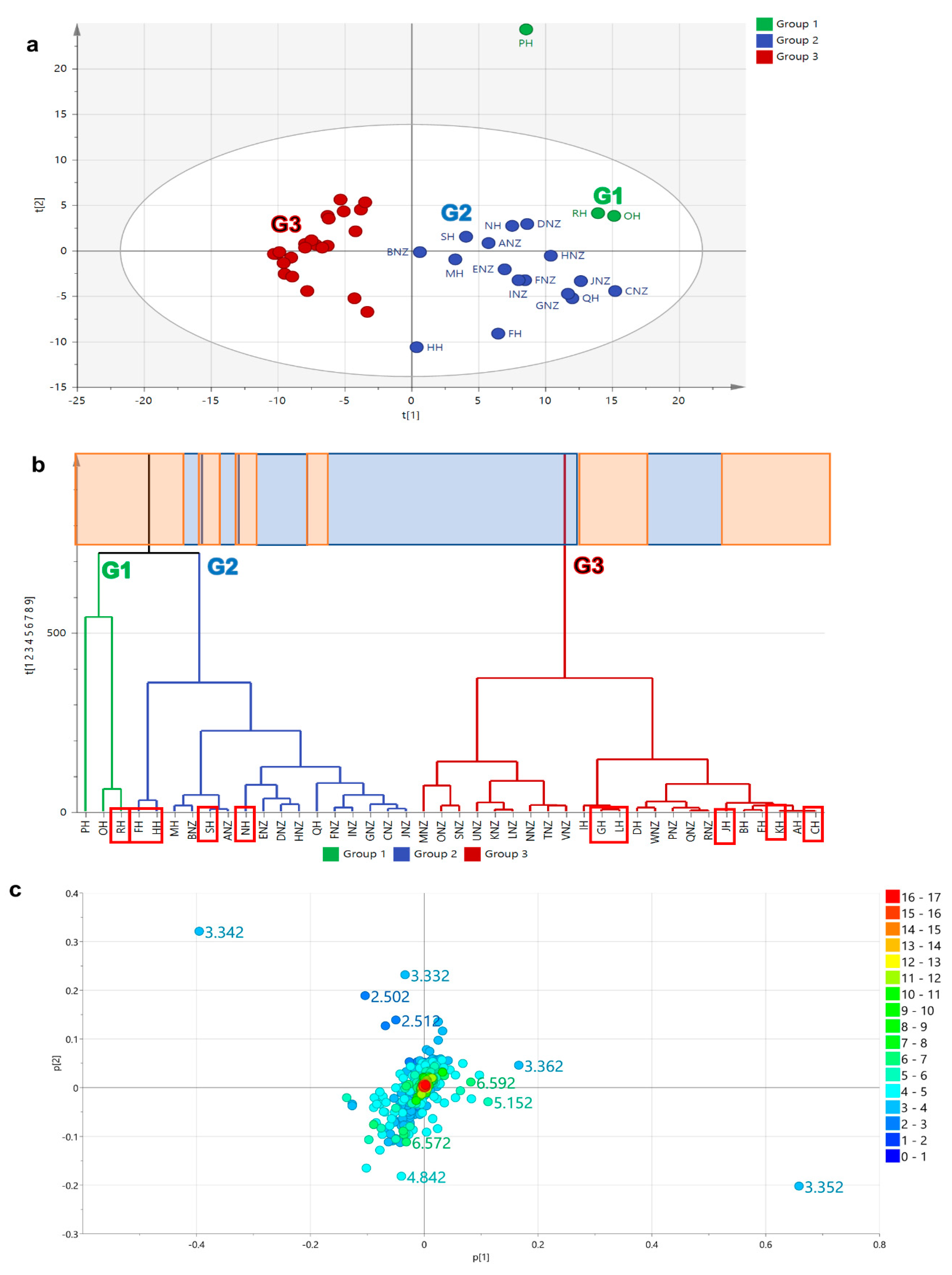
2.5. Multivariate Analysis of Malaysian and New Zealand Honey Samples
2.5.1. Principal Component (PCA) and Hierarchical Clustering Analysis (HCA)
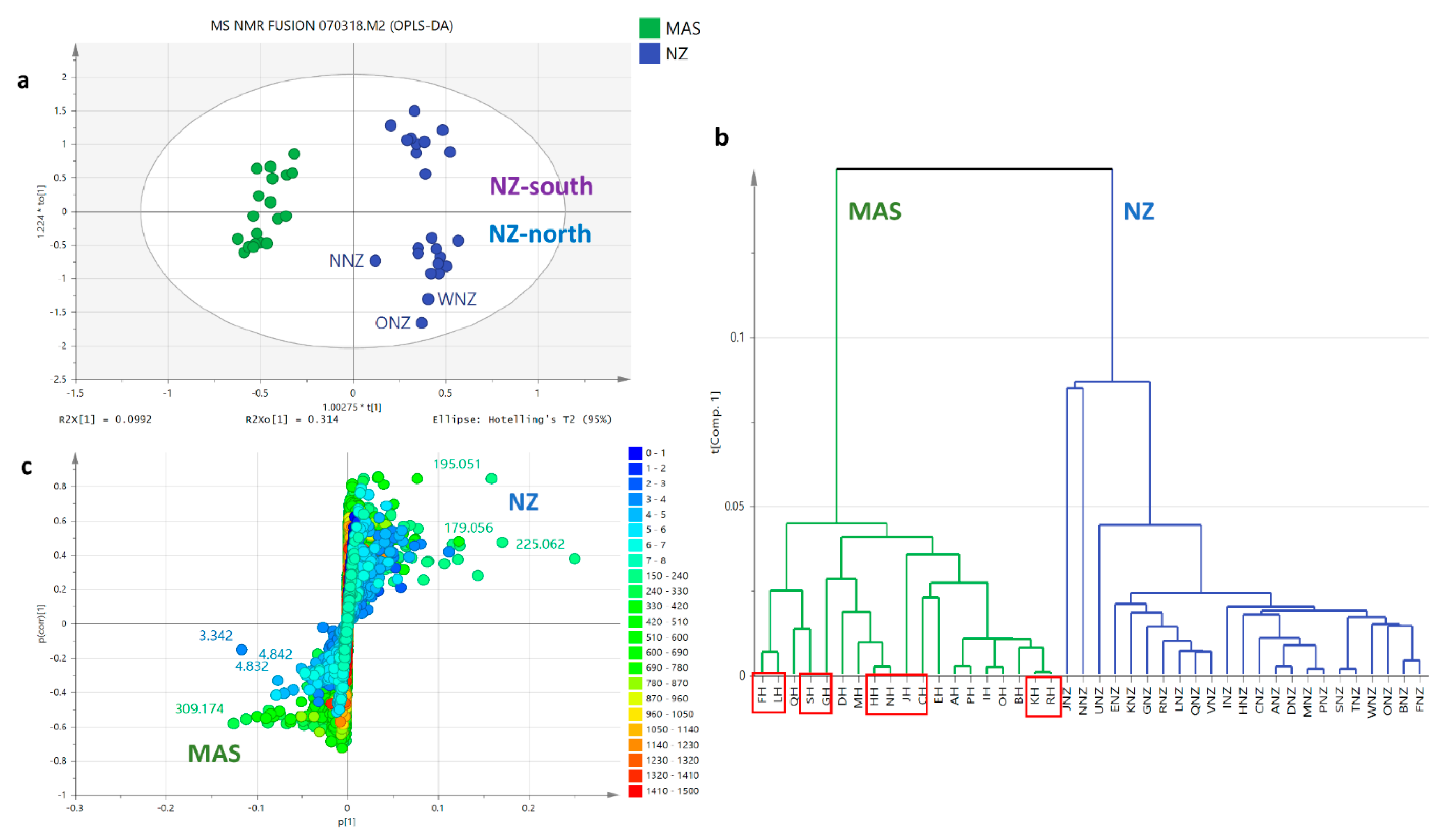
2.5.2. OPLS-DA Based on Geographical Area of Collection
2.6. Dereplication of Bioactive Metabolites
3. Discussion
3.1. Geographical Differences of Collected Honey Samples
3.2. Metabolites from Malaysian Honey
3.3. Metabolites from New Zealand Honey
4. Materials and Methods
4.1. Honey
4.2. Extraction of Fresh Malaysia and New Zealand Honeys
4.3. Liquid Chromatography High Resolution Mass Spectrometry (LC-HRMS) and Dereplication
4.4. Nuclear Magnetic Resonance (NMR) Spectroscopy
4.5. Multivariate Analysis
4.6. Cytotoxicity Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.H.; Gheldof, N.; Engeseth, N.J. Effect of processing and storage on antioxidant capacity of honey. J. Food Sci. 2004, 69, fct96–fct101. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.; Tulipani, S.; Romandini, S.; Bertoli, E.; Battino, M. Contribution of honey in nutrition and human health: A review. Mediterr. J. Nutr. Metab. 2010, 3, 15–23. [Google Scholar] [CrossRef]
- Tsiapara, A.V.; Jaakkola, M.; Chinou, I.; Graikou, K.; Tolonen, T.; Virtanen, V.; Moutsatsou, P. Bioactivity of Greek honey extracts on breast cancer (MCF-7), prostate cancer (PC-3) and endometrial cancer (Ishikawa) cells: Profile analysis of extracts. Food Chem. 2009, 116, 702–708. [Google Scholar] [CrossRef]
- Ghashm, A.; Othman, N.; Khattak, M.; Ismail, N.; Saini, R. Antiproliferative effect of Tualang honey on oral squamous cell carcinoma and osteosarcoma cell lines. BMC Complement. Altern. Med. 2010, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Samarghandian, S.; Afshari, J.T.; Davoodi, S. Honey induces apoptosis in renal cell carcinoma. Pharmacogn. Mag. 2011, 7, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Fauzi, A.N.; Norazmi, M.N.; Yaacob, N.S. Tualang honey induces apoptosis and disrupts the mitochondrial membrane potential of human breast and cervical cancer cell lines. Food Chem. Toxicol. 2011, 49, 871–878. [Google Scholar] [CrossRef]
- Jaganathan, S.K.; Mandal, M. Antiproliferative effects of honey and of its polyphenols: A review. BioMed Res. Int. 2009, 2009, 830616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jubri, Z.; Narayanan, N.N.N.; Karim, N.A.; Ngah, W.Z.W. Antiproliferative activity and apoptosis induction by gelam honey on liver cancer cell line. Int. J. Appl. Sci. Technol. 2012, 2, 135–141. [Google Scholar]
- Vanhanen, L.P.; Emmertz, A.; Savage, G.P. Mineral analysis of mono-floral New Zealand honey. Food Chem. 2011, 128, 236–240. [Google Scholar] [CrossRef]
- Nayik, G.A.; Nanda, V. A chemometric approach to evaluate the phenolic compounds, antioxidant activity and mineral content of different unifloral honey types from Kashmir, India. LWT-Food Sci. Technol. 2016, 74, 504–513. [Google Scholar] [CrossRef]
- Leong, A.G.; Herst, P.M.; Harper, J.L. Indigenous New Zealand honeys exhibit multiple anti-inflammatory activities. Innate Immun. 2012, 18, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonks, A.J.; Cooper, R.A.; Jones, K.P.; Blair, S.; Parton, J.; Tonks, A. Honey stimulates inflammatory cytokine production from monocytes. Cytokine 2003, 21, 242–247. [Google Scholar] [CrossRef]
- Macintyre, L.; Zhang, T.; Viegelmann, C.; Juarez Martinez, I.; Cheng, C.; Dowdells, C.; Abdelmohsen, U.R.; Gernert, C.; Hentschel, U.; Edrada-Ebel, R. Metabolomic tools for secondary metabolite discovery from marine microbial symbionts. Mar. Drugs 2014, 12, 3416–3448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boffo, E.F.; Tavares, L.A.; Tobias, A.C.T.; Ferreira, M.M.C.; Ferreira, A.G. Identification of components of Brazilian honey by 1H NMR and classification of its botanical origin by chemometric methods. LWT-Food Sci. Technol. 2012, 49, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Donarski, J.A.; Roberts, D.P.T.; Charlton, A.J. Quantitative NMR spectroscopy for the rapid measurement of methylglyoxal in manuka honey. Anal. Methods 2010, 2, 1479–1483. [Google Scholar] [CrossRef]
- Lee, R.; Harris, J.; Champagne, P.; Jessop, P.G. CO2-Catalysed conversion of carbohydrates to 5-hydroxymethyl furfural. Green Chem. 2016, 18, 6305–6310. [Google Scholar] [CrossRef]
- Ohmenhaeuser, M.; Monakhova, Y.B.; Kuballa, T.; Lachenmeier, D.W. Qualitative and quantitative control of honeys using NMR spectroscopy and chemometrics. ISRN Anal. Chem. 2013, 2013, 825318. [Google Scholar] [CrossRef]
- Spiteri, M.; Jamin, E.; Thomas, F.; Rebours, A.; Lees, M.; Rogers, K.M.; Rutledge, D.N. Fast and global authenticity screening of honey using 1H-NMR profiling. Food Chem. 2015, 189, 60–66. [Google Scholar] [CrossRef]
- Bohlmann, F.; Schuber, J. Polacetylene compounds. CLXXI. Synthesis of a thio ether isolated from Chrysanthemum coronarium. Chem. Ber. 1969, 102, 4209–4211. [Google Scholar]
- Chen, L.; Xu, H.H.; Hu, T.S.; Wu, Y.L. Synthesis of spiroketal enol ethers related to tonghaosu and their insecticidal activities. Pest Manag. Sci. 2005, 61, 477–482. [Google Scholar] [CrossRef]
- Zheng, C.-J.; Lan, X.-P.; Cheng, R.-B.; Huang, B.-K.; Han, T.; Zhang, Q.-Y.; Zhang, H.; Rahman, K.; Qin, L.-P. Furanofuran lignans from Vitex negundo seeds. Phytochem. Lett. 2011, 4, 298–300. [Google Scholar] [CrossRef]
- Chang, M.-S.; Yang, Y.-C.; Kuo, Y.-C.; Kuo, Y.-H.; Chang, C.; Chen, C.-M.; Lee, T.-H. Furocoumarin glycosides from the leaves of Ficus ruficaulis Merr var. antaoensis. J. Nat. Prod. 2005, 68, 11–13. [Google Scholar] [CrossRef]
- Gribel, N.V.; Pashinskiĭ, V.G. The antitumor properties of honey. Vopr. Onkol. 1990, 36, 704–709. [Google Scholar] [PubMed]
- Hammad, L.A.; Saleh, M.M.; Novotny, M.V.; Mechref, Y. Multiple-reaction monitoring liquid chromatography mass spectrometry for monosaccharide compositional analysis of glycoproteins. J. Am. Soc. Mass Spectrom. 2009, 20, 1224–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, B.R.; Leathwick, J.R. Vegetation-environment relationships at Waipoua Forest, Northland, New Zealand. N. Z. J. Bot. 1996, 34, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Dawson, J.; Sneddon, B.V. The New Zealand Rain Forest: A Comparison with Tropical Rain Forest. Pac. Sci. 1969, XXIII, 131–147. [Google Scholar]
- Mustafa, K.; Perry, N.B.; Weavers, R.T. 2-Hydroxyflavanones from Leptospermum polygalifolium subsp. polygalifolium Equilibrating sets of hemiacetal isomers. Phytochemistry 2003, 64, 1285–1293. [Google Scholar] [CrossRef]
- Porter, N.G.; Wilkins, A.L. Chemical, physical and antimicrobial properties of essential oils of Leptospermum scoparium and Kunzea ericoides. Phytochemistry 1999, 50, 407–415. [Google Scholar] [CrossRef]
- Kikuzaki, H.; Tsai, S.-M.; Nakatani, N. Gingerdiol related compounds from the rhizomes of Zingiber officinale. Phytochemistry 1992, 31, 1783–1786. [Google Scholar] [CrossRef]
- Biplab, D.; Avijit, C.; Tandrima, M.; Shilpi, C.; Goswami, B.B. Evaluation of spermicidal and antimicrobial activities of methanolic extract of Leucas aspera and structural elucidation of separated active component. Asian J. Chem. 2011, 23, 826–828. [Google Scholar]
- Hundal, T.; Forsmark-Andree, P.; Ernster, L.; Andersson, B. Antioxidant activity of reduced plastoquinone in chloroplast thylakoid membranes. Arch. Biochem. Biophys. 1995, 324, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.J.G.; DeOliveira, C.J.C.; Fernandes, J.; Kijjoa, A.; Herz, W. Further alkyl and alkenylphenols of Knema laurina and Knema austrosiamensis: Location of the double bond in the alkenyl side chains. Phytochemistry 1996, 43, 1333–1337. [Google Scholar] [CrossRef]
- Hake, I.; Schonenberger, S.; Neumann, J.; Franke, K.; Paulsen-Merker, K.; Reymann, K.; Ismail, G.; Bin Din, L.; Said, I.M.; Latiff, A.; et al. Neuroprotection and enhanced neurogenesis by extract from the tropical plant Knema laurina after inflammatory damage in living brain tissue. J. Neuroimmunol. 2009, 206, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Nebula, M.; Harisankar, H.S.; Chandramohanakumar, N. Metabolites and bioactivities of Rhizophoraceae mangroves. Nat. Prod. Bioprospect 2013, 3, 207–232. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, M.; Kumar, V.; Singh, S. Phytochemical and pharmacological activity of Genus Plumeria: An updated review. Int. J. Biomed. Adv. Res. 2014, 5, 266–271. [Google Scholar] [CrossRef]
- Malik, A.; Najam-ul-Hussain Kazmi, S.; Ahmed, Z.; Ahmed, W. Plumerinine—A novel lupin alkaloid from Plumeria rubra. Heterocycles 1989, 29, 1901–1906. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Database. Sodium Cetyl Sulfate, CID = 23616669. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Sodium-cetyl-sulfate#section=Canonical-SMILES (accessed on 17 July 2020).
- Costea, T.; Vlad, O.C.; Miclea, L.C.; Ganea, C.; Szollosi, J.; Mocanu, M.M. Alleviation of multidrug resistance by flavonoid and non-flavonoid compounds in breast, lung, colorectal and prostate cancer. Int. J. Mol. Sci. 2020, 21, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, M.; Sirajudeen, K.N.S.; Swamy, M.; Yaacob, M.; Sulaiman, S. Studies on the antioxidant properties of Tualang honey of Malaysia. Afr. J. Tradit. Complement. Altern. Med. 2010, 7, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Sulaiman, S.A.; Othman, N.H. Oral administration of Tualang and Manuka Honeys modulates breast cancer progression in Sprague-Dawley rats model. Evid.-Based Complement. Altern. Med. 2017, 2017, 5904361. [Google Scholar] [CrossRef]
- Yaacob, N.S.; Ismail, N.F. Comparison of cytotoxicity and genotoxicity of 4-hydroxytamoxifen in combination with Tualang honey in MCF-7 and MCF-10A cells. BMC Complement. Altern. Med. 2014, 14, 106. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Othman, N.H. The anti-cancer effects of Tualang honey in modulating breast carcinogenesis: An experimental animal study. BMC Complement. Altern. Med. 2017, 17, 208. [Google Scholar] [CrossRef] [Green Version]
- Kadir, E.A.; Sulaiman, S.A.; Yahya, N.K.; Othman, N.H. Inhibitory effects of Tualang Honey on experimental breast cancer in rats: A preliminary study. Asian Pac. J. Cancer Prev. 2013, 14, 2249–2254. [Google Scholar] [CrossRef] [Green Version]
- Welsch, C.W. Interrelationship between dietary lipids and calories and experimental mammary gland tumorigenesis. Cancer 1994, 74, 1055–1062. [Google Scholar] [CrossRef]
- Fernandez-Cabezudo, M.J.; El-Kharrag, R.; Torab, F.; Bashir, G.; George, J.A.; El-Taji, H.; Al-Ramadi, B.K. Intravenous administration of manuka honey inhibits tumor growth and improves host survival when used in combination with chemotherapy in a melanoma mouse model. PLoS ONE 2013, 8, e55993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hixon, K.R.; Lu, T.; McBride-Gagyi, S.H.; Janowiak, B.E.; Sell, S.A. A comparison of tissue engineering scaffolds incorporated with Manuka honey of varying UMF. BioMed Res. Int. 2017, 2017, 4843065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speer, S.L.; Schreyack, G.E.; Bowlin, G.L. Manuka honey: A tissue engineering essential ingredient. J. Tissue Sci. Eng. 2015, 6, e130. [Google Scholar] [CrossRef] [Green Version]
- Perry, N.B.; Brennan, N.J.; Van Klink, J.W.; Harris, W.; Douglas, M.H.; McGimpsey, J.A.; Smallfield, B.M.; Anderson, R.E. Essential oils from New Zealand manuka and kanuka: Chemotaxonomy of Leptospermum. Phytochemistry 1997, 44, 1485–1494. [Google Scholar] [CrossRef]
- Douglas, M.H.; van Klink, J.W.; Smallfield, B.M.; Perry, N.B.; Anderson, R.E.; Johnstone, P.; Weavers, R.T. Essential oils from New Zealand manuka: Triketone and other chemotypes of Leptospermum scoparium. Phytochemistry 2004, 65, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Afrin, S.; Giampieri, F.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Amici, A.; Cianciosi, D.; Quiles, J.L.; Battino, M. Manuka honey synergistically enhances the chemopreventive effect of 5-fluorouracil on human colon cancer cells by inducing oxidative stress and apoptosis, altering metabolic phenotypes and suppressing metastasis ability. Free Radic. Biol. Med. 2018, 126, 41–54. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | ID | A549 | A2780 | ZR75 | PANC-1 | HFL-1 | No. | ID | A549 | A2780 | ZR75 | PANC-1 | HFL-1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Malaysian Samples | New Zealand Samples | ||||||||||||
| 1 | AH | 86 | 92 | 92 | 85 | 77 | 20 | ANZ | 90 | 79 | 68 | 89 | 87 |
| 2 | BH | 88 | 92 | 94 | 84 | 79 | 21 | BNZ | 89 | 93 | 66 | 88 | 86 |
| 3 | CH | 88 | 92 | 38 | 87 | 84 | 22 | CNZ | 83 | 90 | 93 | 86 | 78 |
| 4 | DH | 88 | 92 | 90 | 86 | 79 | 23 | DNZ | 85 | 91 | 92 | 83 | 71 |
| 5 | EH | 88 | 77 | 71 | 82 | 81 | 24 | ENZ | 86 | 86 | 92 | 88 | 82 |
| 6 | FH | 83 | 92 | 41 | 85 | 78 | 25 | FNZ | 90 | 92 | 69 | 87 | 81 |
| 7 | GH | 88 | 93 | 41 | 86 | 83 | 26 | GNZ | 84 | 92 | 92 | 88 | 82 |
| 8 | HH | 83 | 91 | 39 | 90 | 80 | 27 | HNZ | 88 | 93 | 90 | 88 | 81 |
| 9 | IH | 89 | 88 | 73 | 83 | 82 | 28 | INZ | 90 | 78 | 82 | 88 | 83 |
| 10 | JH | 87 | 93 | 40 | 85 | 81 | 29 | JNZ | 91 | 92 | 68 | 88 | 83 |
| 11 | KH | 84 | 92 | 41 | 83 | 78 | 30 | KNZ | 90 | 86 | 76 | 87 | 84 |
| 12 | LH | 85 | 92 | 42 | 85 | 80 | 31 | LNZ | 88 | 92 | 71 | 88 | 82 |
| 13 | MH | 87 | 92 | 70 | 84 | 74 | 32 | MNZ | 89 | 92 | 67 | 90 | 83 |
| 14 | NH | 87 | 93 | 39 | 89 | 84 | 33 | NNZ | 89 | 92 | 72 | 88 | 78 |
| 15 | OH | 85 | 92 | 73 | 86 | 79 | 34 | ONZ | 89 | 93 | 68 | 88 | 83 |
| 16 | PH | 90 | 91 | 72 | 86 | 81 | 35 | PNZ | 87 | 92 | 72 | 84 | 73 |
| 17 | QH | 87 | 93 | 77 | 80 | 80 | 36 | QNZ | 88 | 94 | 70 | 90 | 82 |
| 18 | RH | 89 | 83 | 43 | 89 | 83 | 37 | RNZ | 89 | 90 | 91 | 89 | 81 |
| 19 | SH | 85 | 75 | 47 | 88 | 83 | 38 | SNZ | 89 | 93 | 73 | 89 | 83 |
| 39 | TNZ | 88 | 91 | 71 | 88 | 77 | |||||||
| 40 | UNZ | 88 | 91 | 73 | 87 | 77 | |||||||
| 41 | VNZ | 88 | 92 | 73 | 89 | 84 | |||||||
| 42 | WNZ | 86 | 92 | 70 | 87 | 82 | |||||||
| Mzmine ID a | m/z | p Values ≤ 0.05 | RT (min) | Predicted Molecular Formula (DBE) b | Accurate Mass (∆ ppm) | Dereplicated Hits | Reported Source c |
|---|---|---|---|---|---|---|---|
| |||||||
| N_1923 * | 309.17 * | 1.08 × 10−4 | 16.15 | C17H26O5 (DBE = 5) | 310.1776 (−1.370181) | gingerdiol 2-hexylphenol-O-β-D-xylopyranoside blumeaene L 9-acetyl-6,7-dihydroxy-3(15)-caryophyllen-8-one | Zingiber officinale Leucas aspera Blumea balsamifera Buddleja davidii |
| N_2025 * | 321.21 | 1.41 × 10−4 | 29.19 | C16H34O4S (DBE = 0) | 322.2178 (0.055863) | cetyl sulphate | Cocos nucifera |
| N_2028 * | 381.23 | 1.60 × 10−4 | 27.52 | C27H30N2 (DBE = 14) | 382.2392 (−4.442225) | no hits | |
| N_4469 * | 311.17 | 1.77 × 10−4 | 20.46 | C10H20N10O2 (DBE = 6) | 312.1761 (−3.107211) | no hits | |
| N_4445 * | 325.184 | 2.14 × 10−4 | 21.55 | no prediction | 326.1916 | no hits | |
| P_3386 * | 295.13 | 3.11 × 10−4 | 19.25 | C19H18O3 (DBE = 11) | 294.1255 (−0.322991) | Please see under 2c and structures shown in Figure 9e | |
| P_4751 * | 413.27 | 3.86 × 10−4 | 32.33 | C22H32N6O2 (DBE = 10) | 412.2590 (0.790766) | no hits | |
| N_4473 * | 339.2 | 3.91 × 10−4 | 26.88 | C23H32O2 (DBE = 8) | 340.2401 (−0.382083) | plastoquinone 3 | Spinacia oleracea |
| N_1976 * | 337.21 | 6.68 × 10−4 | 37.35 | no prediction | 338.2128 | no hits | |
| N_1925 * | 353.20 | 9.43 × 10−4 | 21.38 | C24H32O2 (DBE = 9) | 352.2401 (−0.369066) | 5-(12-phenyl-8-dodecenyl)resorcinol | Knema laurina |
| |||||||
| N_1928 * | 397.227 | 1.65 × 10−16 | 22.19 | C27H30N2O (DBE = 14) | 398.2340 (−4.552579) | no hits | |
| N_7 | 195.05 | 2.39 × 10−11 | 1.33 | C6H12O7 (DBE = 1) | 196.0582 (−0.535555) | gluconic acid | Aureobasidium pullulans |
| N_720 | 215.03 | 3.76 × 10−7 | 1.28 | C16H6O (DBE = 14) | 214.0424 (2.499511) | no hits | |
| N_536 | 387.115 | 1.36 × 10−7 | 1.09 | C13H24O13 (DBE = 2) | 388.1218 (0.270534) | disaccharide (no hits in DNP) | |
| N_1011 | 225.06 | 1.55 × 10−3 | 1.34 | C7H14O8 (DBE = 1) | 226.0688 (−0.309640) | no hits | |
| N_723 | 161.05 | 5.40 × 10−3 | 1.34 | C6H10O5 (DBE = 2) | 162.0528 (−0.148100) | 1,5-anhydrofructose | various plant sources |
| N_30 | 207.05 | 2.85 × 10−2 | 1.58 | C14H6O2 (DBE = 12) | 206.0370 (1.067771) | no hits | |
| N_4692 | 179.06 | 5.49 × 10−2 | 1.33 | C6H12O6 (DBE = 1) | 180.0634 (0.055536) | various hexoses e.g., fructose, allose | various plant sources Protea rubropilosa |
| |||||||
| N_539 | 308.099 | 1.11 × 10−4 | 1.46 | C11H19NO9 (DBE = 3) | 309.1058 (−0.595265) | O-sialic acid (1) | enzymatic hydrolysis of sialoproteins and oligosaccharides |
| N_5017 | 321.210 | 8.28 × 10−3 | 37.32 | C16H34O4S (DBE = 0) | 322.2177 (−0.254486) | cetyl sulphate (2) | Cocos nucifera |
| N_1030 | 470.151 | 1.72 × 10−2 | 1.15 | C30H21N3O3 (DBE = 22) | 471.1587 (0.868073) | no hits | |
| P_5294 | 429.261 | 1.80 × 10−2 | 27.11 | C22H32N6O3 (DBE = 10) | 428.2539 (0.726205) | no hits | |
| N_5083 | 441.253 | 1.80 × 10−2 | 37.23 | C20H42O8S (DBE = 0) | 442.2602 (0.357256) | no hits | |
| P_3154 | 226.180 | 2.76 × 10−2 | 26.94 | C13H23NO2 (DBE = 3) | 225.1728 (−0.350842) | tropine isovalerate(3) | Bruguieras exangula |
| P_6857 | 355.282 | 3.10 × 10−2 | 26.54 | C17H34N6O2 (DBE = 4) | 354.2745 (0.496790) | no hits | |
| P_151 | 226.216 | 3.17×10−2 | 26.39 | C14H27NO (DBE = 2) | 225.2092 (−0.284180) | plumerinine (4) | Plumeria rubra |
| N_41 | 369.103 | 3.53 × 10−2 | 1.11 | C26H14N2O (DBE = 21) | 370.1108 (0.505254) | no hits | |
| P_1365 | 347.096 | 3.54 × 10−2 | 1.09 | C14H18O10 (DBE = 10) | 346.0883 (−4.912017) | tetra-acyl- mannolactone(5) tetra-acyl- ribonic lactone (6) | various plant sources |
| P_3386 | 295.133* | 3.75× 10−2 | 19.25 | C19H18O3 (DBE = 11) | 294.1255 (−0.322991) | dianisylidene acetone (7) 5-dehydroxy artocarbene (8) eupomatenoid 13 (9) methyloroxyl-opterocarpan(10) sterequinone H(11) | Curcuma longa Artocarpus incises Caryodaphnopsis tonkinensis Oroxylum indicum Stereospermum personatum |
| P_6818 | 359.240 | 4.96 × 10−2 | 26.78 | C15H30N6O4 (DBE = 4) | 358.233 (0.407556) | no hits | |
| N_2122 | 279.164 | 5.06 × 10−2 | 20.35 | C13H28O4S (DBE = 0) | 280.1709 (0.242709) | no hits | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yusoff, Y.M.; Abbott, G.; Young, L.; Edrada-Ebel, R. Metabolomic Profiling of Malaysian and New Zealand Honey Using Concatenated NMR and HRMS Datasets. Metabolites 2022, 12, 85. https://doi.org/10.3390/metabo12010085
Yusoff YM, Abbott G, Young L, Edrada-Ebel R. Metabolomic Profiling of Malaysian and New Zealand Honey Using Concatenated NMR and HRMS Datasets. Metabolites. 2022; 12(1):85. https://doi.org/10.3390/metabo12010085
Chicago/Turabian StyleYusoff, Yusnaini M., Grainne Abbott, Louise Young, and RuAngelie Edrada-Ebel. 2022. "Metabolomic Profiling of Malaysian and New Zealand Honey Using Concatenated NMR and HRMS Datasets" Metabolites 12, no. 1: 85. https://doi.org/10.3390/metabo12010085