
Pinus pinaster Early Hormonal Defence Responses to Pinewood Nematode (Bursaphelenchus xylophilus) Infection

 , , and
, , and
Abstract
:
1. Introduction
2. Results
2.1. LC-QqQ-MS/MS Analytical Method Validation
2.1.1. Linearity
2.1.2. Matrix Effects
2.1.3. Limit of Detection (LOD) and Limit of Quantification (LOQ)
2.1.4. Analytical Recoveries
2.1.5. Method Precision
2.2. P. pinaster Early Hormonal Defence Responses to PWN Infection
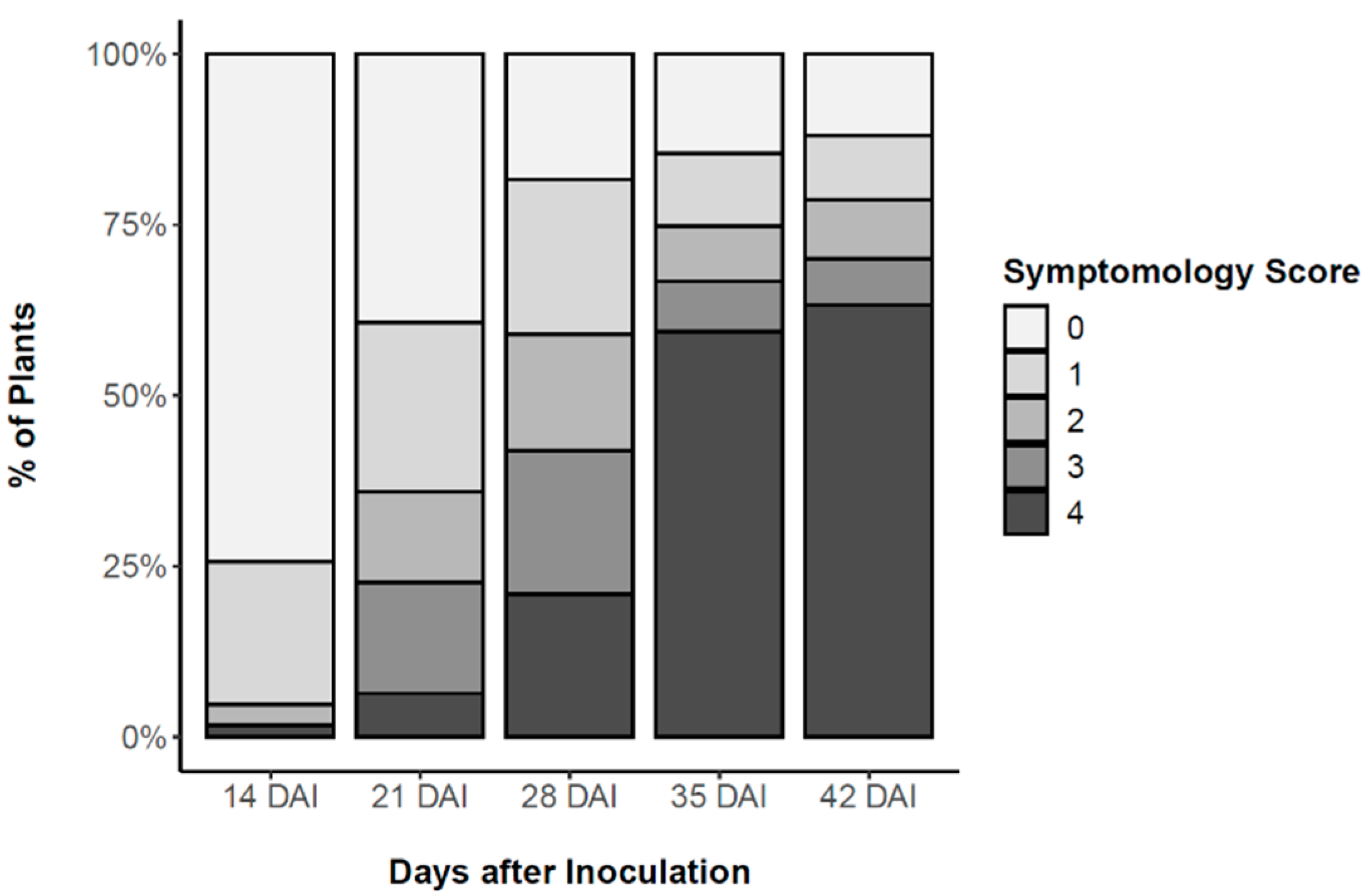
2.2.1. Pine Wilt Disease Progression in PWN-Inoculated P. pinaster Plants
2.2.2. Quantification of Phytohormones in P. pinaster Plants in Response to PWN Infection
3. Discussion
3.1. LC-QqQ-MS/MS Analytical Method Validation
3.2. P. pinaster Early Hormonal Defence Responses to PWN Infection
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material
4.3. LC-QqQ-MS/MS Analytical Method Validation
4.3.1. Standard Stock Solutions and Quality Controls
4.3.2. Extraction of Phytohormones
4.3.3. LC-QqQ-MS/MS Instrument Setup
4.3.4. Calibration Curves and Linearity
4.3.5. Matrix Effects
4.3.6. LOD and LOQ
4.3.7. Analytical Recoveries
4.3.8. Method Precision
4.4. Experimental Design, PWN Inoculation and Sampling Procedure
4.5. Extraction and Quantification of Phytohormones in PWN-Inoculated P. pinaster
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| PH | Treatment | ABA | GA | JA-ME | SA | ZeaR |
|---|---|---|---|---|---|---|
| 24 HAI | H | 162.9 ± 16.3 a | 45.8 ± 7.4 a | 615.7 ± 63.8 a | 573.9 ± 125.5 a | 0.153 ± 0.046 a |
| W | 198.4 ± 47.9 a | 34.1 ± 1.0 a | 577.3 ± 15.5 a | 552.2 ± 151.0 a | 0.086 ± 0.030 a | |
| IN_R | 179.4 ± 73.3 a | 34.1 ± 9.1 a | 526.3 ± 106.6 a | 526.3 ± 185.5 a | 0.206 ± 0.043 a | |
| IN_S | 159.1 ± 30.3 a | 35.2 ± 8.4 a | 703.8 ± 100.6 a | 424.9 ± 96.2 a | 0.120 ± 0.023 a | |
| 48 HAI | H | 190.2 ± 40.7 ab | 47.0 ± 6.7 a | 578.0 ± 92.0 a | 506.2 ± 138.6 b | 0.144 ± 0.030 a |
| W | 207.1 ± 47.9 ab | 42.0 ± 10.4 a | 614.9 ± 86.3 a | 421.0 ± 109.2 b | 0.044 ± 0.008 a | |
| IN_R | 113.9 ± 9.8 b | 47.5 ± 12.6 a | 455.7 ± 15.1 a | 496.3 ± 210.5 b | 0.109 ± 0.030 a | |
| IN_S | 254.6 ± 25.2 a | 47.7 ± 12.5 a | 846.6 ± 91.7 a | 1888.9 ± 657.9 a | 0.136 ± 0.024 a | |
| 72 HAI | H | 408.8 ± 164.7 a | 48.8 ± 11.6 a | 490.1 ± 68.6 b | 681.9 ± 182.7 ab | 0.100 ± 0.034 a |
| W | 134.0 ± 25.6 a | 45.7 ± 9.5 a | 561.5 ± 91.7 b | 609.4 ± 155.9 ab | 0.102 ± 0.019 a | |
| IN_R | 209.3 ± 51.5 a | 55.0 ± 16.4 a | 655.1 ± 65.2 b | 512.3 ± 246.6 b | 0.167 ± 0.052 a | |
| IN_S | 273.2 ± 39.7 a | 36.6 ± 9.6 a | 1131.5 ± 144.4 a | 1583.4 ± 346.4 a | 0.091 ± 0.019 a |
References
- Kuroda, K. Physiological incidences related to symptom development and wilting mechanism. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 204–222. [Google Scholar]
- Mota, M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Rodrigues, J.M. National eradication programme for the pinewood nematode. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Mota, M.M., Vieira, P., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 5–14. [Google Scholar]
- Sousa, E.; Rodrigues, J.M.; Bonifácio, L.F.; Naves, P.M.; Rodrigues, A. Management and control of the pine wood nematode, Bursaphelenchus xylophilus, in Portugal. In Nematodes: Morphology, Functions and Management Strategies; Boeri, F., Chung, J.A., Eds.; Nova Science Publishers Inc.: Boston, MA, USA, 2011; pp. 157–178. [Google Scholar]
- EPPO. PM 9/1 (6) Bursaphelenchus xylophilus and its vectors: Procedures for official control. EPPO Bull. 2018, 48, 503–505. [Google Scholar] [CrossRef] [Green Version]
- Nunes da Silva, M.N.; Solla, A.; Sampedro, L.; Zas, R.; Vasconcelos, M.W. Susceptibility to the pinewood nematode (PWN) of four pine species involved in potential range expansion across Europe. Tree Physiol. 2015, 35, 987–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zas, R.; Moreira, X.; Ramos, M.; Lima, M.R.M.; Nunes da Silva, M.; Solla, A.; Vasconcelos, M.W.; Sampedro, L. Intraspecific variation of anatomical and chemical defensive traits in Maritime pine (Pinus pinaster) as factors in susceptibility to the pinewood nematode (Bursaphelenchus xylophilus). Trees 2015, 29, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.M.; Mendes, M.D.; Lima, A.S.; Barbosa, P.M.; Ascensão, L.; Barroso, J.G.; Pedro, L.G.; Mota, M.M.; Figueiredo, A.C. Pinus halepensis, P. pinaster, P. pinea and P. sylvestris essential oils chemotypes and monoterpene hydrocarbon enantiomers, before and after inoculation with the pinewood nematode Bursaphelenchus xylophilus. Chem. Biodiv. 2017, 14, e1600153. [Google Scholar] [CrossRef] [PubMed]
- Carrasquinho, I.; Lisboa, A.; Inácio, M.L.; Gonçalves, E. Genetic variation in susceptibility to pine wilt disease of maritime pine (Pinus pinaster Aiton) half-sib families. Ann. For. Sci. 2018, 75, 85. [Google Scholar] [CrossRef] [Green Version]
- Menéndez-Gutiérrez, M.; Alonso, M.; Toval, G.; Díaz, R. Testing of selected Pinus pinaster half-sib families for tolerance to pinewood nematode (Bursaphelenchus xylophilus). Forestry 2018, 91, 38–48. [Google Scholar] [CrossRef]
- Erb, M.; Kliebensteinb, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Sena, I.; Vieira da Silva, I.; Ribeiro, B.; Barbosa, P.; Ascensão, L.; Bennett, R.N.; Mota, M.; Figueiredo, A.C. In vitro co-cultures of Pinus pinaster with Bursaphelenchus xylophilus: A biotechnological approach to study pine wilt disease. Planta 2015, 241, 1325–1326. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, H.; Goto, S.; Kazumi, E.; Kuroda, K. The expressed genes of Japanese red pine (Pinus densiflora) involved in the pine wilt disease severity. BMC Proc. 2011, 5, P92. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Kajiwara, J.; Mori, Y.; Ohira, M.; Tsutsumi, Y.; Kondo, R. Metabolites from resistant and susceptible Pinus thunbergii after inoculation with pine wood nematode. Am. J. Plant Sci. 2013, 4, 28699. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, C.S.; Gonçalves, E.V.; Firmino, P.N.; Calvão, T.; Fonseca, L.; Abrantes, I.; Correia, O.; Máguas, C. Differences in pine species constitutive and inducible defences determining the susceptibility to the pinewood nematode. Plant Pathol. 2017, 66, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Canas, S.; Trindade, C.S.; Sun, B.; Naves, P. Phenolic compounds involved in pine wilt disease: HPLC-based method development and validation for their quantification. J. Plant Biochem. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Gaspar, D.; Trindade, C.; Usié, A.; Meireles, B.; Barbosa, P.; Fortes, A.M.; Pesquita, C.; Costa, R.L.; Ramos, A.M. Expression profiling in Pinus pinaster in response to infection with the pine wood nematode Bursaphelenchus xylophilus. Forests 2017, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wei, Y.; Xu, L.; Hao, Y.; Chen, X.; Zhou, Z. Transcriptomic profiling reveals differentially expressed genes associated with pine wood nematode resistance in Masson Pine (Pinus massoniana Lamb.). Sci. Rep. 2017, 7, 4693. [Google Scholar] [CrossRef]
- Lee, I.H.; Kim, J.; Woo, K.S.; Jang, K.-H.; Kim, Y.-H.; Shim, D. De novo assembly and transcriptome analysis of the Pinus densiflora response to pine wilt disease in nature. Plant Biotechnol. Rep. 2018, 12, 229–236. [Google Scholar] [CrossRef]
- Lee, I.H.; Han, H.; Koh, Y.H.; Kim, I.S.; Lee, S.-W.; Shim, D. Comparative transcriptome analysis of Pinus densiflora following inoculation with pathogenic (Bursaphelenchus xylophilus) or non-pathogenic nematodes (B. thailandae). Sci. Rep. 2019, 9, 12180. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Wina, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 2016, 4, 162–176. [Google Scholar]
- Dubois, M.; Van den Broeck, L.; Inzé, D. The pivotal role of ethylene in plant growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone crosstalk in plant disease and defense: More than just jasmonates-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 17–43. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.; van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; van Wees, S.C. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Thaler, J.S.; Humphrey, P.T.; Whiteman, N.K. Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 2012, 17, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Davies, P.J. The plant hormones: Their nature, occurrence, and functions. In Plant Hormones; Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 1–15. [Google Scholar]
- Fu, J.; Sun, X.; Wang, J.; Chu, J.; Yan, C. Progress in quantitative analysis of plant hormones. Sci. Bull. 2011, 56, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Rawlinson, C.; Kamphuis, L.G.; Gummer, J.P.A.; Singh, K.B.; Trengove, R.D. A rapid method for profiling of volatile and semi-volatile phytohormones using methyl chloroformate derivatization and GC-MS. Metabolomics 2015, 11, 1922–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.; Kriechbaumer, V.; Park, W.J. Modern quantitative analytical tools and biosensors for functional studies of auxin. J. Plant Biol. 2016, 59, 93–104. [Google Scholar] [CrossRef]
- Cao, D.; Lutz, A.; Hill, C.B.; Callahan, D.; Roessner, U. A quantitative profiling method of phytohormones and other metabolites applied to barley roots subjected to salinity stress. Front. Plant Sci. 2017, 7, 2070. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Welti, R.; Wang, X. Simultaneous quantification of major phytohormones and related compounds in crude plant extracts by liquid chromatography–electrospray tandem mass spectrometry. Phytochemistry 2008, 69, 1773–1781. [Google Scholar] [CrossRef]
- Pan, X.; Wang, X. Profiling of plant hormones by mass spectrometry. J. Chromatogr. B 2009, 877, 280–2813. [Google Scholar] [CrossRef]
- Trapp, M.A.; de Souza, G.D.; Rodrigues-Filho, E.; Boland, W.; Mithӧfer, A. Validated method for phytohormone quantification in plants. Front. Plant Sci. 2014, 5, 417. [Google Scholar]
- Ross, A.R.S.; Ambrose, S.J.; Cutler, A.J.; Feurtado, A.; Kermose, A.R.; Nelson, K.; Zhou, R.; Abrams, S.R. Determination of endogenous and supplied deuterated abscisic acid in plant tissues by high-performance liquid chromatography-electrospray ionization tandem mass spectrometry with multiple reaction monitoring. Anal. Biochem. 2004, 329, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, L.; Fan, C.Q.; Wang, P. Determination of abscisic acid, gibberellic acid, indole-3-acetic acid, and zeatin riboside in masson pine [Pinus massoniana L.] by accelerated solvent extraction and high-performance liquid chromatography—Tandem mass spectrometry. Anal. Lett. 2016, 49, 1986–1996. [Google Scholar] [CrossRef]
- Delatorre, C.; Rodríguez, A.; Rodríguez, L.; Majada, J.P.; Ordás, R.J.; Feito, I. Hormonal profiling: Development of a simple method to extract and quantify phytohormones in complex matrices by UHPLC–MS/MS. J. Chromatogr. B 2017, 1040, 239–249. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; António, C. Standard key steps in mass spectrometry-based plant metabolomics experiments: Instrument performance and analytical method validation. In Plant Metabolomics. Methods in Molecular Biology; António, C., Ed.; Humana Press: New York, NY, USA, 2018; pp. 19–31. [Google Scholar]
- Moosavi, S.M.; Ghassabian, S. Linearity of calibration curves for analytical methods: A review of criteria for assessment of method reliability. In Calibration and Validation of Analytical Methods—A Sampling of Current Approaches; IntechOpen Ltd.: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Welti, R.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef]
- Balcke, G.U.; Handrick, V.; Bergau, N.; Fichtner, M.; Henning, A.; Stellmach, H.; Tissier, A.; Hause, B.; Frolov, A. An UPLC-MS/MS method for highly sensitive high-throughput analysis of phytohormones in plant tissues. Plant Methods 2012, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Zhang, H.; Xu, W.; He, X.; Yang, L.; Luo, Y.; Huang, K. Simultaneous determination of 15 plant growth regulators in bean sprout and tomato with liquid chromatography-triple quadrupole tandem mass spectrometry. Food Anal. Methods 2013, 6, 941–951. [Google Scholar] [CrossRef]
- Floková, K.; Tarkowská, D.; Miersch, O.; Strnad, M.; Wasternack, C.; Novák, O. UHPLC–MS/MS based target profiling of stress-induced phytohormones. Phytochemistry 2014, 105, 147–157. [Google Scholar] [CrossRef]
- Wu, Z.; Gao, W.; Phelps, M.A.; Wu, D.; Miller, D.D.; Dalton, J.T. Favorable effects of weak acids on negative-ion electrospray ionization mass spectrometry. Anal. Chem. 2004, 76, 839–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Y.; Jenke, D. Increasing the sensitivity of an LC-MS method for screening material extracts for organic extractables via mobile phase optimization. J. Chromatogr. Sci. 2012, 50, 213–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalczyk, M.; Sandberg, G. Quantitative analysis of indole-3-acetic acid metabolites in Arabidopsis. Plant Physiol. 2001, 127, 1845–1853. [Google Scholar] [CrossRef]
- Correia, B.; Pintó-Marijuan, M.; Castro, B.B.; Brossa, R.; López-Carbonell, M.; Pinto, G. Hormonal dynamics during recovery from drought in two Eucalyptus globulus genotypes: From root to leaf. Plant Physiol. Biochem. 2014, 82, 151–160. [Google Scholar] [CrossRef]
- Thakare, R.; Chhonker, Y.S.; Gautam, N.; Alamoudi, J.A. Quantitative analysis of endogenous compounds. J. Pharm. Biomed. Anal. 2016, 128, 426–437. [Google Scholar] [CrossRef]
- Koek, M.M.; Jellema, R.H.; van der Greef, J.; Tas, A.C.; Hankemeier, T. Quantitative metabolomics based on gas chromatography mass spectrometry: Status and perspectives. Metabolomics 2011, 7, 307–328. [Google Scholar] [CrossRef] [Green Version]
- George, E.F.; Hall, M.A.; Klerk, G.J.D. Plant growth regulators I: Introduction; auxins, their analogues and inhibitors. In Plant Propagation by Tissue Culture; George, E.F., Hall, M.A., Klerk, G.J.D., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 175–204. [Google Scholar]
- Hirao, T.; Fukatsu, E.; Watanabe, A. Characterization of resistance to pine wood nematode infection in Pinus thunbergii using suppression subtractive hybridization. BMC Plant Biol. 2012, 12, 13. [Google Scholar] [CrossRef] [Green Version]
- Balint-Kurti, P. The plant hypersensitive response: Concepts, control and consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, R.F. Pathogenesis in pine wilt caused by pinewood nematode, Bursaphelenchus xylophilus. J. Nematol. 1988, 20, 236–244. [Google Scholar] [PubMed]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, S.; Evans, H.; Ikegami, M. Modeling PWN-induced wilt expression: A mechanistic approach. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Mota, M.M., Vieira, P., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 259–278. [Google Scholar]
- Li, Z.; Zhang, Q.; Zhou, X. A 2-Cys peroxiredoxin in response to oxidative stress in the pine wood nematode, Bursaphelenchus xylophilus. Sci. Rep. 2016, 6, 27438. [Google Scholar] [CrossRef]
- Yin, R.; Liu, X.; Yu, J.; Ji, Y.; Liu, J.; Cheng, L.; Zhou, J. Up-regulation of autophagy by low concentration of salicylic acid delays methyl jasmonate-induced leaf senescence. Sci. Rep. 2020, 10, 11472. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, A.A.; Gori, A.; Da-Silva, C.J.; Brunetti, C. Abscisic acid biosynthesis and signaling in plants: Key targets to improve water use efficiency and drought tolerance. Appl. Sci. 2020, 10, 6322. [Google Scholar] [CrossRef]
- Denancé, N.; Sánchez-Vallet, A.; Goffner, D.; Molina, A. Disease resistance or growth: The role of plant hormones in balancing immune responses and fitness costs. Front. Plant Sci. 2013, 4, 155. [Google Scholar] [CrossRef] [Green Version]
- Mauch-Mani, B.; Mauch, F. The role of abscisic acid in plant-pathogen interactions. Curr. Opin. Plant Biol. 2005, 8, 409–414. [Google Scholar] [CrossRef]
- Ribeiro, B.; Espada, M.; Vu, T.; Nóbrega, F.; Mota, M.; Carrasquinho, I. Pine wilt disease: Detection of the pinewood nematode (Bursaphelenchus xylophilus) as a tool for a pine breeding programme. For. Pathol. 2012, 42, 521–525. [Google Scholar] [CrossRef]
- Matuszewski, B.K.; Constanzer, M.L.; Chavez-Eng, C.M. Strategies for the assessment of matrix effect in quantitative bioanalytical methods based on HPLC-MS/MS. Anal. Chem. 2003, 75, 3019–3030. [Google Scholar] [CrossRef]
- Hartmann, C.; Smeyers-Verbeke, J.; Massart, D.L.; McDowall, R.D. Validation of bioanalytical chromatographic methods. J. Pharm. Biomed. Anal. 1998, 17, 193–218. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2018; Available online: http://www.rstudio.com/ (accessed on 8 December 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 8 December 2020).
- Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3-3. 2020. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 8 December 2020).
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Thomas, L.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. Gplots: Various R Programming Tools for Plotting Data. R Package Version 3.0.3. 2020. Available online: https://CRAN.R-project.org/package=gplots (accessed on 8 December 2020).


| Analyte (PH) | IM | tR (min) | SRM | CE (V) | Analyte (IS) | IM | tR (min) | SRM | CE (V) |
|---|---|---|---|---|---|---|---|---|---|
| ABA | – | 4.08 | 263.1 > 153.1 * | 10.354 | d6-ABA | – | 4.07 | 269.1 > 159.1 | 10.354 |
| 263.1 > 219.1 | 13.438 | ||||||||
| BA | – | 3.80 | 121.2 > 77.2 * | 11.719 | d5-BA | – | 3.78 | 126.2 > 82.2 * | 10.253 |
| 121.2 > 93.0 | 10.809 | 126.2 > 125.6 | 35.129 | ||||||
| GA | – | 3.46 | 345.2 > 239.1 * | 15.562 | d2-GA9 | – | 5.03 | 317.2 > 273.2 | 21.174 |
| 345.2 > 221.1 | 25.421 | ||||||||
| GA9 | – | 5.02 | 315.2 > 271.2 * | 21.022 | d2-GA9 | – | 5.03 | 317.2 > 273.2 | 21.174 |
| 315.2 > 253.1 | 27.242 | ||||||||
| IAA | – | 3.84 | 174.1 > 130.1 * | 10.253 | d5-IAA | – | 3.83 | 179.1 > 135.2 * | 10.253 |
| 174.1 > 128.1 | 19.152 | 179.1 > 133.1 | 19.404 | ||||||
| IAA-ME | + | 4.45 | 190.2 > 130.1 * | 10.253 | d5-IAA-ME | + | 4.45 | 195.1 > 134.1 * | 10.253 |
| 190.2 > 103.2 | 35.989 | 195.1 > 135.1 | 13.640 | ||||||
| IBA | + | 4.35 | 204.2 > 130.1 * | 25.876 | d5-IAA-ME | + | 4.45 | 195.1 > 134.1 * | 10.253 |
| 204.2 > 144.1 | 22.893 | 195.1 > 135.1 | 13.640 | ||||||
| IBA-ME | + | 4.85 | 218.2 > 186.0 * | 10.253 | d5-IAA-ME | + | 4.45 | 195.1 > 134.1 * | 10.253 |
| 218.2 > 130.1 | 27.646 | 195.1 > 135.1 | 13.640 | ||||||
| iP | + | 3.00 | 204.2 > 136.0 * | 15.511 | d5-Zea | + | 0.60 | 225.3 > 137.1 * | 17.888 |
| 204.2 > 148.0 | 10.253 | 225.3 > 148.0 | 15.360 | ||||||
| JA | – | 4.39 | 209.1 > 59.3 * | 14.096 | DHJA | – | 4.59 | 211.2 > 59.3 * | 14.096 |
| 209.1 > 165.1 | 10.253 | 211.2 > 167.1 | 17.685 | ||||||
| JA-ME | + | 4.92 | 225.2 > 151.1 * | 12.073 | DHJA-ME | + | 4.80 | 227.2 > 135.1 * | 10.253 |
| 225.2 > 133.1 | 14.803 | 227.2 > 153.1 | 12.225 | ||||||
| SA | – | 4.07 | 137.2 > 93.2 * | 16.219 | d4-SA | – | 4.10 | 141.1 > 97.2 * | 17.180 |
| 137.2 > 65.3 | 28.455 | 141.1 > 69.3 | 29.719 | ||||||
| Zea | + | 0.61 | 220.2 > 136.1 * | 17.180 | d5-Zea | + | 0.60 | 225.3 > 137.1 * | 17.888 |
| 220.2 > 148.0 | 14.753 | 225.3 > 148.0 | 15.360 | ||||||
| ZeaR | + | 0.63 | 352.2 > 220.0 * | 19.000 | d5-Zea | + | 0.60 | 225.3 > 137.1 * | 17.888 |
| 352.2 > 136.1 | 31.893 | 225.3 > 148.0 | 15.360 |
| Analyte (PH) | Conc. Range (ng/mL) | Linearity (Solvent) | R2 | Linearity (Matrix) | R2 | ME (%) | LOD (ng/mL) | LOQS (ng/mL) | LOQM (ng/g) |
|---|---|---|---|---|---|---|---|---|---|
| ABA | 5–1500 | 0.00251x + 0.0143 | 0.993 | 0.00218x + 0.0234 | 0.991 | –13 | 1 | 5 | 5 |
| BA | 50–5000 | 0.000428x + 0.0969 | 0.996 | 0.000478x + 0.0980 | 0.998 | +12 | 10 | 50 | 50 |
| GA | 100–1500 | 0.000053x − 0.0019 | 0.993 | 0.000119x − 0.00441 | 0.991 | +123 | 5 | 10 | 50 |
| GA9 | 100–1500 | 0.00182x + 0.0172 | 0.996 | 0.00185x + 0.0226 | 0.991 | +2 | 5 | 50 | 100 |
| IAA | 50–1500 | 0.00255x + 0.00695 | 0.992 | 0.00155x + 0.00850 | 0.992 | –41 | 5 | 50 | 50 |
| IAA-ME | 50–1500 | 0.00322x − 0.0398 | 0.990 | 0.00322x − 0.0343 | 0.990 | –0.1 | 5 | 10 | 50 |
| IBA | 50–1500 | 0.00129x − 0.0231 | 0.994 | 0.00149x − 0.2189 | 0.992 | +15 | 5 | 10 | 50 |
| IBA-ME | 50–1500 | 0.00461x − 0.117 | 0.996 | 0.00495x − 0.0332 | 0.991 | +7 | 1 | 10 | 50 |
| iP | 1–50 | 0.0665x − 0.0222 | 0.996 | 0.0693x − 0.0269 | 0.993 | +4 | 0.005 | 0.05 | 0.5 |
| JA | 50–5000 | 0.000212x + 0.0190 | 0.991 | 0.000355x + 0.0195 | 0.990 | +67 | 5 | 50 | 50 |
| JA-ME | 100–1500 | 0.00365x − 0.0792 | 0.996 | 0.000522x − 0.0317 | 0.991 | –85 | 50 | 50 | 100 |
| SA | 5–50 | 0.0119x + 0.0299 | 0.995 | 0.00901x + 0.176 | 0.991 | –24 | 0.1 | 0.5 | 5 |
| Zea | 5–50 | 0.0160x + 0.000412 | 0.998 | 0.0177x + 0.000550 | 0.990 | +11 | 0.001 | 0.01 | 0.1 |
| ZeaR | 5–50 | 0.0102x + 0.00246 | 0.999 | 0.0117x + 0.00345 | 0.990 | +16 | 0.001 | 0.01 | 0.1 |
| Analyte (PH) | Concentration (ng/mL) | Analytical Recovery (%) ± SE | Instrument Precision (RSD, %) | Intraday Precision (RSD, %) | Interday Precision (RSD, %) |
|---|---|---|---|---|---|
| ABA | 1500 | 84.9 ± 4.4 | 2.8 | 2.1 | 11.9 |
| 500 | 80.5 ± 5.9 | 5.3 | 5.8 | 10.7 | |
| 50 | 73.6 ± 5.4 | 14.2 | 8.9 | 22.2 | |
| BA | 5000 | 94.1 ± 2.0 | 0.6 | 3.0 | 6.0 |
| 2000 | 84.7 ± 5.2 | 0.7 | 0.7 | 4.4 | |
| 500 | 79.9 ± 14.8 | 2.7 | 1.8 | 6.6 | |
| GA | 1500 | 83.8 ± 8.1 | 4.6 | 0.5 | 7.4 |
| 500 | 88.5 ± 9.1 | 9.0 | 4.5 | 6.3 | |
| 100 | 63.8 ± 20.0 | 7.7 | 12.8 | 22.0 | |
| GA9 | 1500 | 97.5 ± 0.3 | 2.7 | 0.3 | 1.7 |
| 500 | 88.3 ± 3.8 | 2.3 | 1.6 | 1.7 | |
| 100 | 81.7 ± 5.5 | 5.7 | 2.1 | 6.8 | |
| IAA | 1500 | 71.2 ± 7.7 | 10.6 | 2.4 | 10.1 |
| 500 | 70.8 ± 8.9 | 13.4 | 5.5 | 12.3 | |
| 50 | 52.9 ± 14.2 | 12.6 | 16.9 | 19.3 | |
| IAA-ME | 1500 | 94.7 ± 4.0 | 1.4 | 1.7 | 11.7 |
| 500 | 89.2 ± 2.8 | 2.4 | 1.2 | 10.0 | |
| 50 | 81.9 ± 1.9 | 9.9 | 5.8 | 22.9 | |
| IBA | 1500 | 93.1 ± 8.8 | 4.0 | 4.0 | 16.4 |
| 500 | 74.5 ± 5.6 | 2.4 | 3.9 | 15.9 | |
| 50 | 67.2 ± 13.4 | 8.5 | 3.6 | 22.9 | |
| IBA-ME | 1500 | 90.6 ± 9.2 | 3.1 | 1.4 | 17.9 |
| 500 | 89.0 ± 0.4 | 2.8 | 9.0 | 21.6 | |
| 50 | 76.8 ± 3.9 | 6.3 | 12.3 | 18.4 | |
| iP | 50 | 89.5 ± 2.9 | 1.3 | 1.7 | 20.2 |
| 35 | 83.9 ± 9.2 | 3.8 | 1.0 | 20.3 | |
| 5 | 67.9 ± 10.0 | 2.7 | 3.7 | 25.0 | |
| JA | 5000 | 100.6 ± 7.5 | 2.4 | 0.3 | 5.6 |
| 2000 | 71.1 ± 1.3 | 5.3 | 5.0 | 6.5 | |
| 500 | 67.6 ± 10.5 | 13.5 | 8.2 | 9.1 | |
| JA-ME | 1500 | 90.3 ± 3.3 | 3.7 | 3.6 | 22.8 |
| 500 | 80.0 ± 3.6 | 3.9 | 3.7 | 22.1 | |
| 100 | 71.3 ± 28.5 | 15.9 | 13.4 | 18.1 | |
| SA | 50 | 92.2 ± 2.6 | 14.2 | 1.2 | 2.8 |
| 35 | 88.9 ± 3.5 | 14.4 | 4.0 | 4.3 | |
| 5 | 70.8 ± 3.2 | 1.3 | 2.2 | 5.5 | |
| Zea | 50 | 97.5 ± 3.6 | 2.3 | 1.6 | 3.4 |
| 35 | 84.4 ± 1.2 | 0.9 | 2.1 | 3.9 | |
| 5 | 69.2 ± 17.6 | 2.0 | 3.3 | 4.6 | |
| ZeaR | 50 | 92.9 ± 13.1 | 2.5 | 1.3 | 8.1 |
| 35 | 85.6 ± 1.0 | 2.0 | 3.2 | 10.3 | |
| 5 | 66.9 ± 11.2 | 5.5 | 1.5 | 21.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, A.M.; Langer, S.; Carrasquinho, I.; Bergström, E.; Larson, T.; Thomas-Oates, J.; António, C. Pinus pinaster Early Hormonal Defence Responses to Pinewood Nematode (Bursaphelenchus xylophilus) Infection. Metabolites 2021, 11, 227. https://doi.org/10.3390/metabo11040227
Rodrigues AM, Langer S, Carrasquinho I, Bergström E, Larson T, Thomas-Oates J, António C. Pinus pinaster Early Hormonal Defence Responses to Pinewood Nematode (Bursaphelenchus xylophilus) Infection. Metabolites. 2021; 11(4):227. https://doi.org/10.3390/metabo11040227
Chicago/Turabian StyleRodrigues, Ana M., Swen Langer, Isabel Carrasquinho, Ed Bergström, Tony Larson, Jane Thomas-Oates, and Carla António. 2021. "Pinus pinaster Early Hormonal Defence Responses to Pinewood Nematode (Bursaphelenchus xylophilus) Infection" Metabolites 11, no. 4: 227. https://doi.org/10.3390/metabo11040227