
LDH and PDH Activities in the Ischemic Brain and the Effect of Reperfusion—An Ex Vivo MR Study in Rat Brain Slices Using Hyperpolarized [1-13C]Pyruvate

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
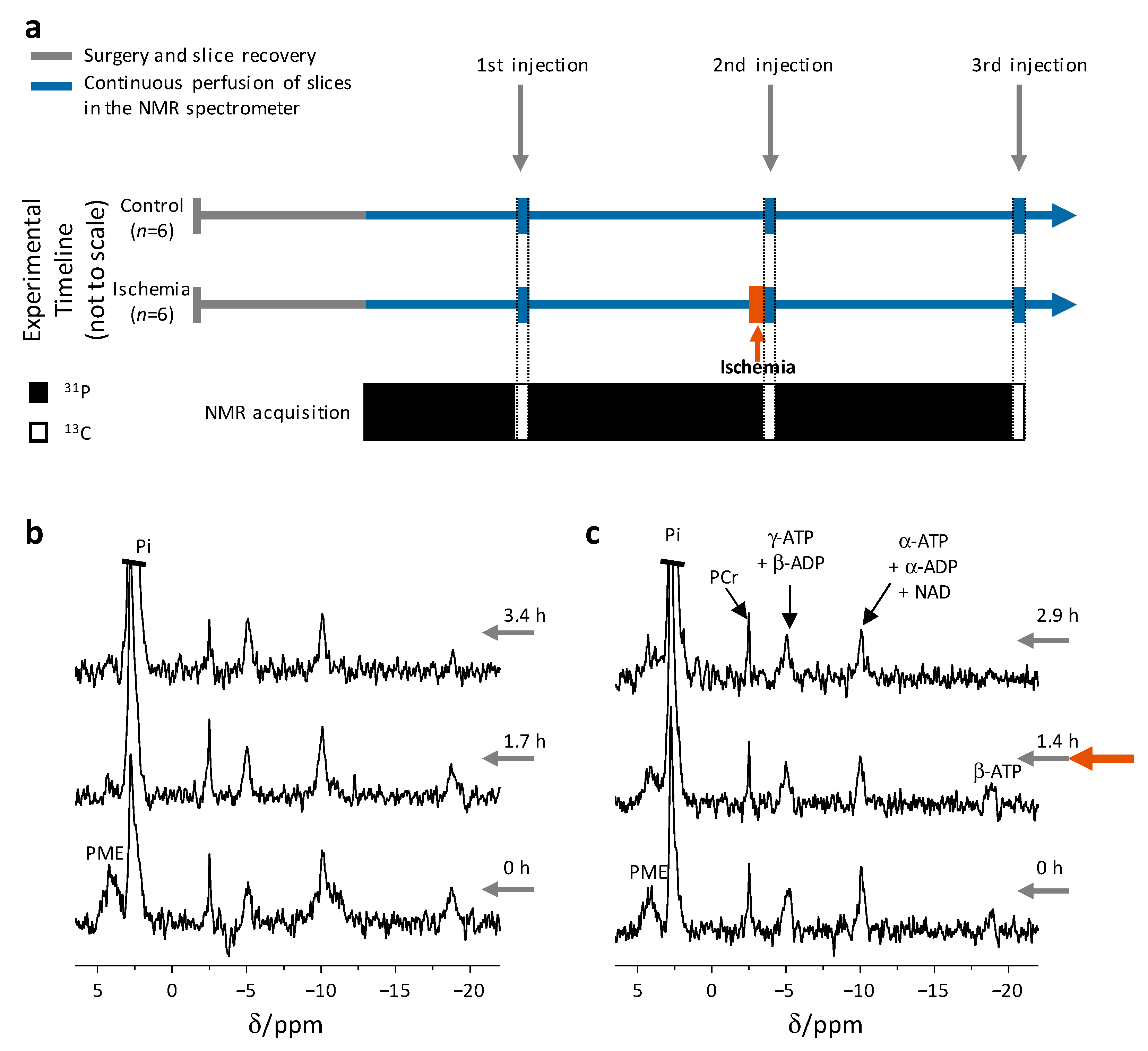
2.1. Brain Slices Remain Viable throughout the Experiment
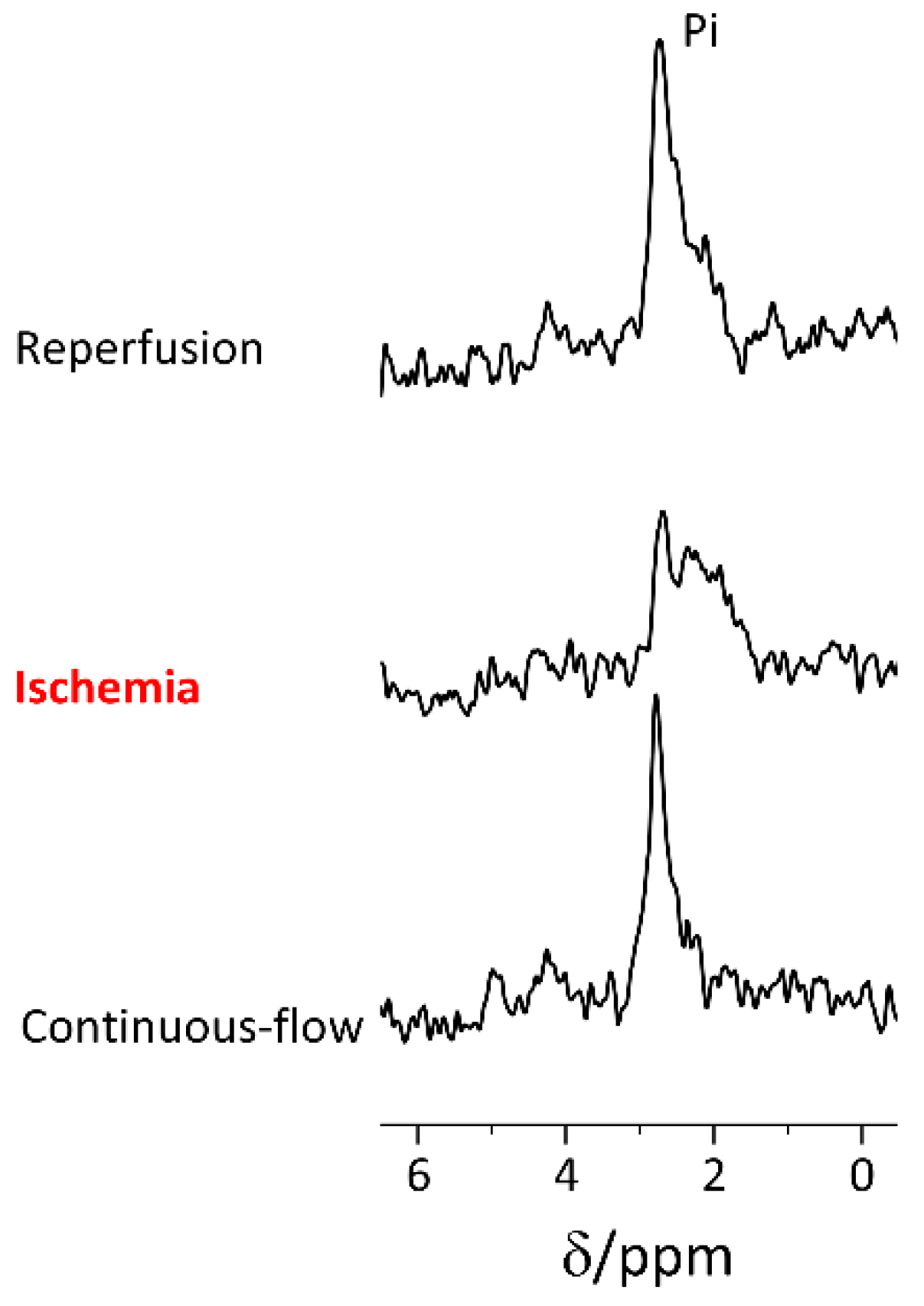
2.2. Rapid Changes in the Slices’ pH and Energetic State Revealed by 31P NMR Spectroscopy
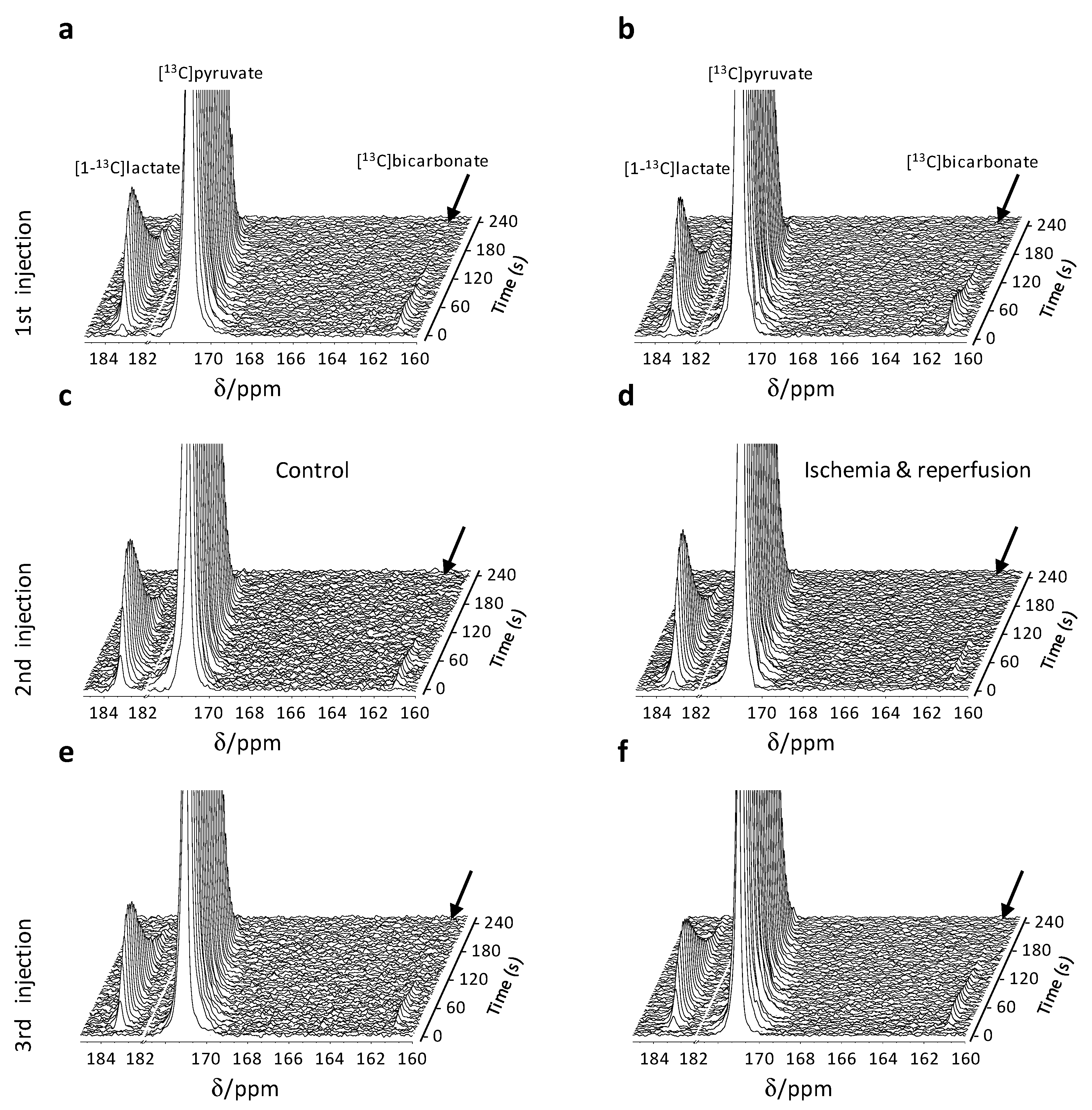
2.3. Hyperpolarized [1-13C]Pyruvate Metabolism Reveals Reversible Metabolic Changes Following an Ischemic Insult and Reperfusion
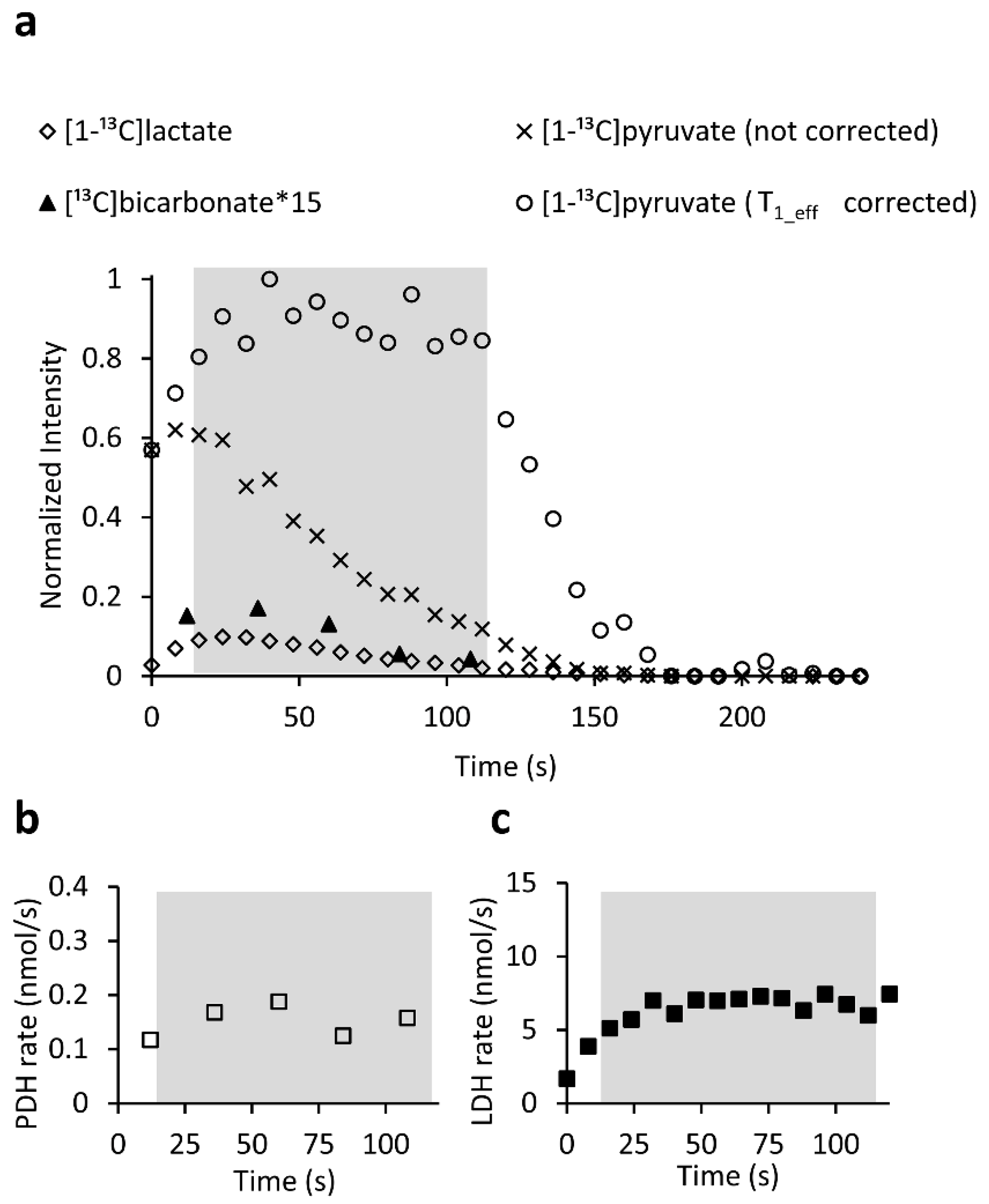
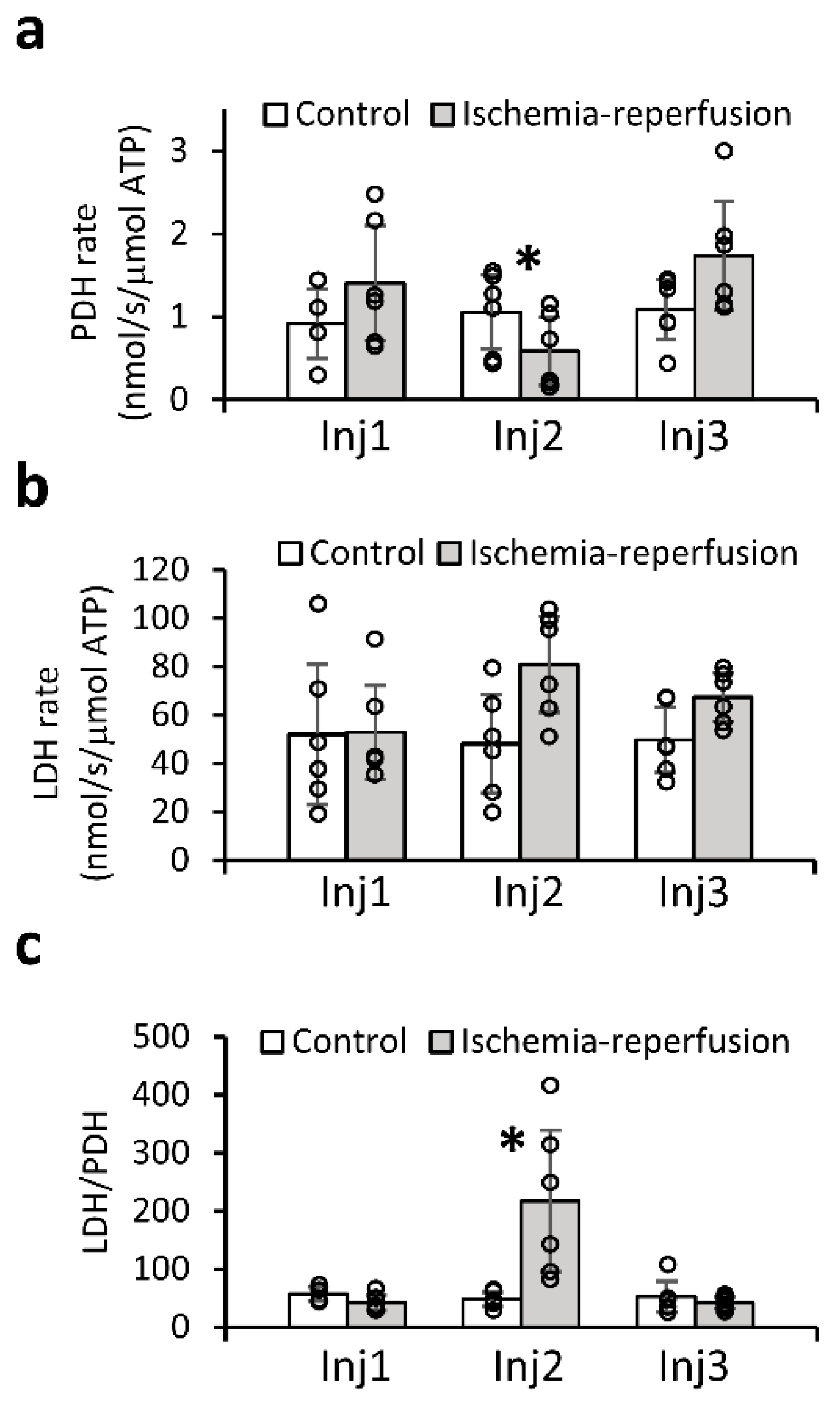
2.4. Quantitative Analysis of the Enzymatic Activities Demonstrates the Utility of the LDH/PDH Ratio as an Indicator of Ischemia-Reperfusion and Recovery of the Brain
3. Discussion
Limitations
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Solutions/Media
4.4. Surgery and Slice Handling
4.5. Perfusion System and the Administration of Hyperpolarized Medium to Brain Slices
4.6. Experimental Workflow
4.7. DNP Spin Polarization and Dissolution
4.8. NMR Spectroscopy
4.8.1. 31P Spectroscopy
4.8.2. Hyperpolarized 13C spectroscopy
4.9. Calculation of Metabolic Rates
4.10. Determination of Slice ATP Content
4.11. Spectral Analysis
4.12. pH Determination
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feigin, V.L.; Nichols, E.; Alam, T.; Bannick, M.S.; Beghi, E.; Blake, N.; Culpepper, W.J.; Dorsey, E.R.; Elbaz, A.; Ellenbogen, R.G.; et al. Global, regional, and national burden of neurological disorders, 1990-2016: A systematic analysis for the global burden of disease study. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef] [Green Version]
- Feigin, V.L.; Forouzanfar, M.H.; Krishnamurthi, R.; Mensah, G.A.; Connor, M.; Bennett, D.A.; Moran, A.E.; Sacco, R.L.; Anderson, L.; Truelsen, T.; et al. Global and regional burden of stroke during 1990-2010: Findings from the global burden of disease study. Lancet 2014, 383, 245–254. [Google Scholar] [CrossRef]
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart disease and stroke statistics-2020 update: A report from the american heart association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef]
- Zerna, C.; Thomalla, G.; Campbell, B.C.V.; Rha, J.H.; Hill, M.D. Current practice and future directions in the diagnosis and acute treatment of ischaemic stroke. Lancet 2018, 392, 1247–1256. [Google Scholar] [CrossRef]
- Powers, W.J.; Rabinstein, A.A.; Ackerson, T.; Adeoye, O.M.; Bambakidis, N.C.; Becker, K.; Biller, J.; Brown, M.; Demaerschalk, B.M.; Hoh, B.; et al. 2018 guidelines for the early management of patients with acute ischemic stroke: A guideline for healthcare professionals from the american heart association/american stroke association. Stroke 2018, 49, e46–e110. [Google Scholar] [CrossRef] [PubMed]
- Puig, J.; Shankar, J.; Liebeskind, D.; Terceño, M.; Nael, K.; Demchuk, A.M.; Menon, B.; Dowlatshahi, D.; Leiva-Salinas, C.; Wintermark, M.; et al. From “time is brain” to “imaging is brain”: A paradigm shift in the management of acute ischemic stroke. J. Neuroimaging Off. J. Am. Soc. Neuroimaging 2020, 30, 562–571. [Google Scholar] [CrossRef]
- Gomez, C.R. Editorial: Time is brain! J. Stroke Cereb. Dis. 1993, 3, 1–2. [Google Scholar] [CrossRef]
- Patel, M.R.; Edelman, R.R.; Warach, S. Detection of hyperacute primary intraparenchymal hemorrhage by magnetic resonance imaging. Stroke 1996, 27, 2321–2324. [Google Scholar] [CrossRef]
- Fiebach, J.B.; Schellinger, P.D.; Gass, A.; Kucinski, T.; Siebler, M.; Villringer, A.; Olkers, P.; Hirsch, J.G.; Heiland, S.; Wilde, P.; et al. Stroke magnetic resonance imaging is accurate in hyperacute intracerebral hemorrhage: A multicenter study on the validity of stroke imaging. Stroke 2004, 35, 502–506. [Google Scholar] [CrossRef]
- Kang, D.W.; Chalela, J.A.; Dunn, W.; Warach, S. MRI screening before standard tissue plasminogen activator therapy is feasible and safe. Stroke 2005, 36, 1939–1943. [Google Scholar] [CrossRef] [PubMed]
- Ringelstein, E.B. Ultrafast magnetic resonance imaging protocols in stroke. J. Neurol. Neurosurg. Psychiatry 2005, 76, 905. [Google Scholar] [CrossRef] [Green Version]
- U-King-Im, J.M.; Trivedi, R.A.; Graves, M.J.; Harkness, K.; Eales, H.; Joubert, I.; Koo, B.; Antoun, N.; Warburton, E.A.; Gillard, J.H.; et al. Utility of an ultrafast magnetic resonance imaging protocol in recent and semi-recent strokes. J. Neurol. Neurosurg. Psychiatry 2005, 76, 1002–1005. [Google Scholar] [CrossRef] [Green Version]
- Chung, M.S.; Lee, J.Y.; Jung, S.C.; Baek, S.; Shim, W.H.; Park, J.E.; Kim, H.S.; Choi, C.G.; Kim, S.J.; Lee, D.H.; et al. Reliability of fast magnetic resonance imaging for acute ischemic stroke patients using a 1.5-T scanner. Eur. Radiol. 2019, 29, 2641–2650. [Google Scholar] [CrossRef]
- Simonsen, C.Z.; Madsen, M.H.; Schmitz, M.L.; Mikkelsen, I.K.; Fisher, M.; Andersen, G. Sensitivity of diffusion- and perfusion-weighted imaging for diagnosing acute ischemic stroke is 97.5%. Stroke 2015, 46, 98–101. [Google Scholar] [CrossRef] [Green Version]
- Pound, P.; Ram, R. Are researchers moving away from animal models as a result of poor clinical translation in the field of stroke? An analysis of opinion papers. BMJ Open 2020, 4, e100041. [Google Scholar] [CrossRef]
- Schanne, F.A.X.; Gupta, R.K.; Stanton, P.K. 31P-NMR study of transient ischemia in rat hippocampal slices in vitro. Biochim. Biophys. Acta 1993, 1158, 257–263. [Google Scholar] [CrossRef]
- Espanol, M.T.; Litt, L.; Chang, L.H.; James, T.L.; Weinstein, P.R.; Chan, P.H. Adult rat brain-slice preparation for nuclear magnetic resonance spectroscopy studies of hypoxia. Anesthesiology 1996, 84, 201–210. [Google Scholar] [CrossRef]
- Komatsumoto, S.; Nioka, S.; Greenberg, J.H.; Yoshizaki, K.; Subramanian, V.H.; Chance, B.; Reivich, M. Cerebral energy metabolism measured in vivo by 31P-NMR in middle cerebral artery occlusion in the cat—Relation to severity of stroke. J. Cereb. Blood Flow Metab. 1987, 7, 557–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thulborn, K.R.; du Boulay, G.H.; Duchen, L.W.; Radda, G. A 31P nuclear magnetic resonance in vivo study of cerebral ischaemia in the gerbil. J. Cereb. Blood Flow Metab. 1982, 2, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Horikawa, Y.; Naruse, S.; Hirakawa, K.; Tanaka, C.; Nishikawa, H.; Watari, H. In vivo studies of energy metabolism in experimental cerebral ischemia using topical magnetic resonance. Changes in 31P-nuclear magnetic resonance spectra compared with electroencephalograms and regional cerebral blood flow. J. Cereb. Blood Flow Metab. 1985, 5, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.R.; Welch, K.M.A.; Helpern, J.A.; Chopp, M.; Bruce, R.; Selwa, J.; Smith, M.B. Prolonged deterioration of ischemic brain energy metabolism and acidosis associated with hyperglycemia: Human cerebral infarction studied by serial 31P NMR spectroscopy. Ann. Neurol. 1988, 23, 416–418. [Google Scholar] [CrossRef] [PubMed]
- Ardenkjær-Larsen, J.H.; Fridlund, B.; Gram, A.; Hansson, G.; Hansson, L.; Lerche, M.H.; Servin, R.; Thaning, M.; Golman, K. Increase in signal-to-noise ratio of > 10,000 times in liquid-state NMR. Proc. Natl. Acad. Sci. USA 2003, 100, 10158. [Google Scholar] [CrossRef] [Green Version]
- Harris, T.; Uppala, S.; Lev-Cohain, N.; Adler-Levy, Y.; Shaul, D.; Nardi-Schreiber, A.; Sapir, G.; Azar, A.; Gamliel, A.; Sosna, J.; et al. Hyperpolarized product selective saturating-excitations for determination of changes in metabolic reaction rates in real-time. NMR Biomed. 2020, 33, e4189. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.H.; Lau, J.Y.C.; Chen, A.P.; Geraghty, B.J.; Perks, W.J.; Roifman, I.; Wright, G.A.; Connelly, K.A. Hyperpolarized 13C metabolic MRI of the human heart: Initial experience. Circ. Res. 2016, 119, 1177–1182. [Google Scholar] [CrossRef]
- Park, I.; Larson, P.E.Z.; Gordon, J.W.; Carvajal, L.; Chen, H.-Y.; Bok, R.; Van Criekinge, M.; Ferrone, M.; Slater, J.B.; Xu, D.; et al. Development of methods and feasibility of using hyperpolarized carbon-13 imaging data for evaluating brain metabolism in patient studies. Magn. Reson. Med. 2018, 80, 864–873. [Google Scholar] [CrossRef] [Green Version]
- Nelson, S.J.; Kurhanewicz, J.; Vigneron, D.B.; Larson, P.E.Z.; Harzstark, A.L.; Ferrone, M.; van Criekinge, M.; Chang, J.W.; Bok, R.; Park, I.; et al. Metabolic imaging of patients with prostate cancer using hyperpolarized [1-¹³C]pyruvate. Sci. Transl. Med. 2013, 5, 198ra108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, S.E.; Kettunen, M.I.; Gallagher, F.A.; Hu, D.E.; Lerche, M.; Wolber, J.; Golman, K.; Ardenkjaer-Larsen, J.H.; Brindle, K.M. Detecting tumor response to treatment using hyperpolarized 13C magnetic resonance imaging and spectroscopy. Nat. Med. 2007, 13, 1382–1387. [Google Scholar] [CrossRef]
- Gallagher, F.A.; Kettunen, M.I.; Day, S.E.; Hu, D.-E.; Ardenkjaer-Larsen, J.H.; in’t Zandt, R.; Jensen, P.R.; Karlsson, M.; Golman, K.; Lerche, M.H.; et al. Magnetic resonance imaging of pH in vivo using hyperpolarized 13C-labelled bicarbonate. Nature 2008, 453, 940–943. [Google Scholar] [CrossRef]
- Gallagher, F.A.; Kettunen, M.I.; Hu, D.E.; Jensen, P.R.; Zandt, R.I.T.; Karlsson, M.; Gisselsson, A.; Nelson, S.K.; Witney, T.H.; Bohndiek, S.E.; et al. Production of hyperpolarized [1,4-C-13(2)]malate from [1,4-C-13(2)]fumarate is a marker of cell necrosis and treatment response in tumors. Proc. Natl. Acad. Sci. USA 2009, 106, 19801–19806. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, T.B.; Serrao, E.M.; Kennedy, B.W.C.; Hu, D.E.; Kettunen, M.I.; Brindle, K.M. Magnetic resonance imaging of tumor glycolysis using hyperpolarized 13C-labeled glucose. Nat. Med. 2014, 20, 93. [Google Scholar] [CrossRef] [Green Version]
- Chavarria, L.; Romero-Gimenez, J.; Monteagudo, E.; Lope-Piedrafita, S.; Cordoba, J. Real-time assessment of 13C metabolism reveals an early lactate increase in the brain of rats with acute liver failure. NMR Biomed. 2015, 28, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Josan, S.; Hurd, R.; Billingsley, K.; Senadheera, L.; Park, J.M.; Yen, Y.F.; Pfefferbaum, A.; Spielman, D.; Mayer, D. Effects of isoflurane anesthesia on hyperpolarized 13C metabolic measurements in rat brain. Magn. Reson. Med. 2013, 70, 1117–1124. [Google Scholar] [CrossRef] [Green Version]
- Mayer, D.; Yen, Y.F.; Takahashi, A.; Josan, S.; Tropp, J.; Rutt, B.K.; Hurd, R.E.; Spielman, D.M.; Pfefferbaum, A. Dynamic and high-resolution metabolic imaging of hyperpolarized [1-13C]pyruvate in the rat brain using a high-performance gradient insert. Magn. Reson. Med. 2011, 65, 1228–1233. [Google Scholar] [CrossRef] [Green Version]
- Marjańska, M.; Iltis, I.; Shestov, A.A.; Deelchand, D.K.; Nelson, C.; Uğurbil, K.; Henry, P.-G. In vivo 13C spectroscopy in the rat brain using hyperpolarized [1-13C]pyruvate and [2-13C]pyruvate. J. Magn. Reson. 2010, 206, 210–218. [Google Scholar] [CrossRef] [Green Version]
- Guglielmetti, C.; Najac, C.; Didonna, A.; Van der Linden, A.; Ronen, S.M.; Chaumeil, M.M. Hyperpolarized 13C MR metabolic imaging can detect neuroinflammation in vivo in a multiple sclerosis murine model. Proc. Natl. Acad. Sci. USA 2017, 114, E6982. [Google Scholar] [CrossRef] [Green Version]
- DeVience, S.J.; Lu, X.; Proctor, J.; Rangghran, P.; Melhem, E.R.; Gullapalli, R.; Fiskum, G.M.; Mayer, D. Metabolic imaging of energy metabolism in traumatic brain injury using hyperpolarized [1-13C]pyruvate. Sci. Rep. 2017, 7, 1907. [Google Scholar] [CrossRef]
- Marjańska, M.; Shestov, A.A.; Deelchand, D.K.; Kittelson, E.; Henry, P.G. Brain metabolism under different anesthetic conditions using hyperpolarized [1-13C]pyruvate and [2-13C]pyruvate. NMR Biomed. 2018, 31, e4012. [Google Scholar] [CrossRef]
- Butt, S.A.; Sogaard, L.V.; Magnusson, P.O.; Lauritzen, M.H.; Laustsen, C.; Akeson, P.; Ardenkjaer-Larsen, J.H. Imaging cerebral 2-ketoisocaproate metabolism with hyperpolarized 13C magnetic resonance spectroscopic imaging. J. Cereb. Blood. Flow Metab. 2012, 32, 1508–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishkovsky, M.; Comment, A. Hyperpolarized MRS: New tool to study real-time brain function and metabolism. Anal. Biochem. 2017, 529, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Hurd, R.E.; Yen, Y.-F.; Tropp, J.; Pfefferbaum, A.; Spielman, D.M.; Mayer, D. Cerebral dynamics and metabolism of hyperpolarized [1-13C]pyruvate using time-resolved MR spectroscopic imaging. J. Cereb. Blood Flow Metab. 2010, 30, 1734–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.J.; Grist, J.T.; Serres, S.; Larkin, J.R.; Lau, A.Z.; Ray, K.; Fisher, K.R.; Hansen, E.; Tougaard, R.S.; Nielsen, P.M. 13C pyruvate transport across the blood-brain barrier in preclinical hyperpolarised MRI. Sci. Rep. 2018, 8, 15082. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Ringgaard, S.; Mariager, C.Ø.; Bertelsen, L.B.; Schroeder, M.; Qi, H.; Laustsen, C.; Stødkilde-Jørgensen, H. Hyperpolarized 13C magnetic resonance imaging can detect metabolic changes characteristic of penumbra in ischemic stroke. Tomography 2017, 3, 67–73. [Google Scholar] [CrossRef]
- Hyacinthe, J.-N.; Buscemi, L.; Lê, T.P.; Lepore, M.; Hirt, L.; Mishkovsky, M. Evaluating the potential of hyperpolarised [1-13C] L-lactate as a neuroprotectant metabolic biosensor for stroke. Sci. Rep. 2020, 10, 5507. [Google Scholar] [CrossRef]
- Grist, J.T.; McLean, M.A.; Riemer, F.; Schulte, R.F.; Deen, S.S.; Zaccagna, F.; Woitek, R.; Daniels, C.J.; Kaggie, J.D.; Matyz, T.; et al. Quantifying normal human brain metabolism using hyperpolarized [1-13C]pyruvate and magnetic resonance imaging. NeuroImage 2019, 189, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Miloushev, V.Z.; Granlund, K.L.; Boltyanskiy, R.; Lyashchenko, S.K.; DeAngelis, L.M.; Mellinghoff, I.K.; Brennan, C.W.; Tabar, V.; Yang, T.J.; Holodny, A.I.; et al. Metabolic imaging of the human brain with hyperpolarized 13C pyruvate demonstrates 13C lactate production in brain tumor patients. Cancer Res. 2018, 78, 3755–3760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, T.; Azar, A.; Sapir, G.; Gamliel, A.; Nardi-Schreiber, A.; Sosna, J.; Gomori, J.M.; Katz-Brull, R. Real-time ex-vivo measurement of brain metabolism using hyperpolarized [1-13C]pyruvate. Sci. Rep. 2018, 8, 9564. [Google Scholar] [CrossRef] [PubMed]
- Gillies, R.J.; Raghunand, N.; Garcia-Martin, M.L.; Gatenby, R.A. pH imaging. A review of pH measurement methods and applications in cancers. IEEE Eng. Med. Biol. Mag. 2004, 23, 57–64. [Google Scholar] [CrossRef]
- Moon, R.B.; Richards, J.H. Determination of intracellular pH by 31P magnetic resonance. J. Biol. Chem. 1973, 248, 7276–7278. [Google Scholar] [CrossRef]
- Lutz, N.W.; Le Fur, Y.; Chiche, J.; Pouysségur, J.; Cozzone, P.J. Quantitative in vivo characterization of intracellular and extracellular pH profiles in heterogeneous tumors: A novel method enabling multiparametric pH analysis. Cancer Res. 2013, 73, 4616–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittingham, T.; Lust, W.; Passonneau, J. An in vitro model of ischemia: Metabolic and electrical alterations in the hippocampal slice. J. Neurosci. 1984, 4, 793–802. [Google Scholar] [CrossRef]
- Lev-Cohain, N.; Sapir, G.; Uppala, S.; Nardi-Schreiber, A.; Goldberg, S.N.; Adler-Levy, Y.; Sosna, J.; Gomori, J.M.; Katz-Brull, R. Differentiation of heterogeneous mouse liver from HCC by hyperpolarized 13C magnetic resonance. Science 2021, 3, 8. [Google Scholar] [CrossRef]
- Mikrogeorgiou, A.; Chen, Y.; Lee, B.S.; Bok, R.; Sheldon, R.A.; Barkovich, A.J.; Xu, D.; Ferriero, D.M. A metabolomics study of hypoxia ischemia during mouse brain development using hyperpolarized 13C. Dev. Neurosci. 2020, 42, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Rosafio, K.; Castillo, X.; Hirt, L.; Pellerin, L. Cell-specific modulation of monocarboxylate transporter expression contributes to the metabolic reprograming taking place following cerebral ischemia. Neuroscience 2016, 317, 108–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Page, L.M.; Guglielmetti, C.; Taglang, C.; Chaumeil, M.M. Imaging brain metabolism using hyperpolarized 13C magnetic resonance spectroscopy. Trends Neurosci. 2020, 43, 343–354. [Google Scholar] [CrossRef]
- Slupe, A.M.; Kirsch, J.R. Effects of anesthesia on cerebral blood flow, metabolism, and neuroprotection. J. Cereb. Blood Flow Metab. 2018, 38, 2192–2208. [Google Scholar] [CrossRef]
- Arikan, F.; Martínez-Valverde, T.; Sánchez-Guerrero, Á.; Campos, M.; Esteves, M.; Gandara, D.; Torné, R.; Castro, L.; Dalmau, A.; Tibau, J.; et al. Malignant infarction of the middle cerebral artery in a porcine model. A pilot study. PLoS ONE 2017, 12, e0172637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorby-Adams, A.J.; Vink, R.; Turner, R.J. Large animal models of stroke and traumatic brain injury as translational tools. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R165–R190. [Google Scholar] [CrossRef]
- Topping, G.J.; Heid, I.; Trajkovic-Arsic, M.; Kritzner, L.; Grashei, M.; Hundshammer, C.; Aigner, M.; Skinner, J.G.; Braren, R.; Schilling, F. Hyperpolarized 13C spectroscopy with simple slice-and-frequency-selective excitation. Biomedicines 2021, 9, 121. [Google Scholar] [CrossRef] [PubMed]
- Kettunen, M.I.; Hu, D.E.; Witney, T.H.; McLaughlin, R.; Gallagher, F.A.; Bohndiek, S.E.; Day, S.E.; Brindle, K.M. Magnetization transfer measurements of exchange between hyperpolarized [1-13C]pyruvate and [1-13C]lactate in a murine lymphoma. Magn. Reson. Med. 2010, 63, 872–880. [Google Scholar] [CrossRef]
- Schulte, R.F.; Sperl, J.I.; Weidl, E.; Menzel, M.I.; Janich, M.A.; Khegai, O.; Durst, M.; Ardenkjaer-Larsen, J.H.; Glaser, S.J.; Haase, A. Saturation-recovery metabolic-exchange rate imaging with hyperpolarized [1-13C] pyruvate using spectral-spatial excitation. Magn. Reson. Med. 2013, 69, 1209–1216. [Google Scholar] [CrossRef] [Green Version]
- Ting, J.T.; Daigle, T.L.; Chen, Q.; Feng, G. Acute brain slice methods for adult and aging animals: Application of targeted patch clamp analysis and optogenetics. Methods Mol. Biol. 2014, 1183, 221–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ting, J.T.; Lee, B.R.; Chong, P.; Soler-Llavina, G.; Cobbs, C.; Koch, C.; Zeng, H. Preparation of acute brain slices using an optimized N-methyl-D-glucamine protective recovery method. J. Vis. Exp. 2018, 132, e53825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardi-Schreiber, A.; Gamliel, A.; Harris, T.; Sapir, G.; Sosna, J.; Gomori, J.M.; Katz-Brull, R. Biochemical phosphates observed using hyperpolarized 31P in physiological aqueous solutions. Nat. Commun. 2017, 8, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massiot, D.; Fayon, F.; Capron, M.; King, I.; Le Calvé, S.; Alonso, B.; Durand, J.-O.; Bujoli, B.; Gan, Z.; Hoatson, G. Modelling one- and two-dimensional solid-state NMR spectra. Magm. Reson. Chem. 2002, 40, 70–76. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice-Hall, Inc.: Upper Saddle River, NJ, USA, 2010; pp. 226–230, 253–257. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sapir, G.; Shaul, D.; Lev-Cohain, N.; Sosna, J.; Gomori, M.J.; Katz-Brull, R. LDH and PDH Activities in the Ischemic Brain and the Effect of Reperfusion—An Ex Vivo MR Study in Rat Brain Slices Using Hyperpolarized [1-13C]Pyruvate. Metabolites 2021, 11, 210. https://doi.org/10.3390/metabo11040210
Sapir G, Shaul D, Lev-Cohain N, Sosna J, Gomori MJ, Katz-Brull R. LDH and PDH Activities in the Ischemic Brain and the Effect of Reperfusion—An Ex Vivo MR Study in Rat Brain Slices Using Hyperpolarized [1-13C]Pyruvate. Metabolites. 2021; 11(4):210. https://doi.org/10.3390/metabo11040210
Chicago/Turabian StyleSapir, Gal, David Shaul, Naama Lev-Cohain, Jacob Sosna, Moshe J. Gomori, and Rachel Katz-Brull. 2021. "LDH and PDH Activities in the Ischemic Brain and the Effect of Reperfusion—An Ex Vivo MR Study in Rat Brain Slices Using Hyperpolarized [1-13C]Pyruvate" Metabolites 11, no. 4: 210. https://doi.org/10.3390/metabo11040210