
A Study of Blood Fatty Acids Profile in Hyperlipidemic and Normolipidemic Subjects in Association with Common PNPLA3 and ABCB1 Polymorphisms


Abstract
:1. Introduction
2. Results
2.1. Blood FA Profiling
2.1.1. Analytical Performance of the Method
2.1.2. FA Concentrations
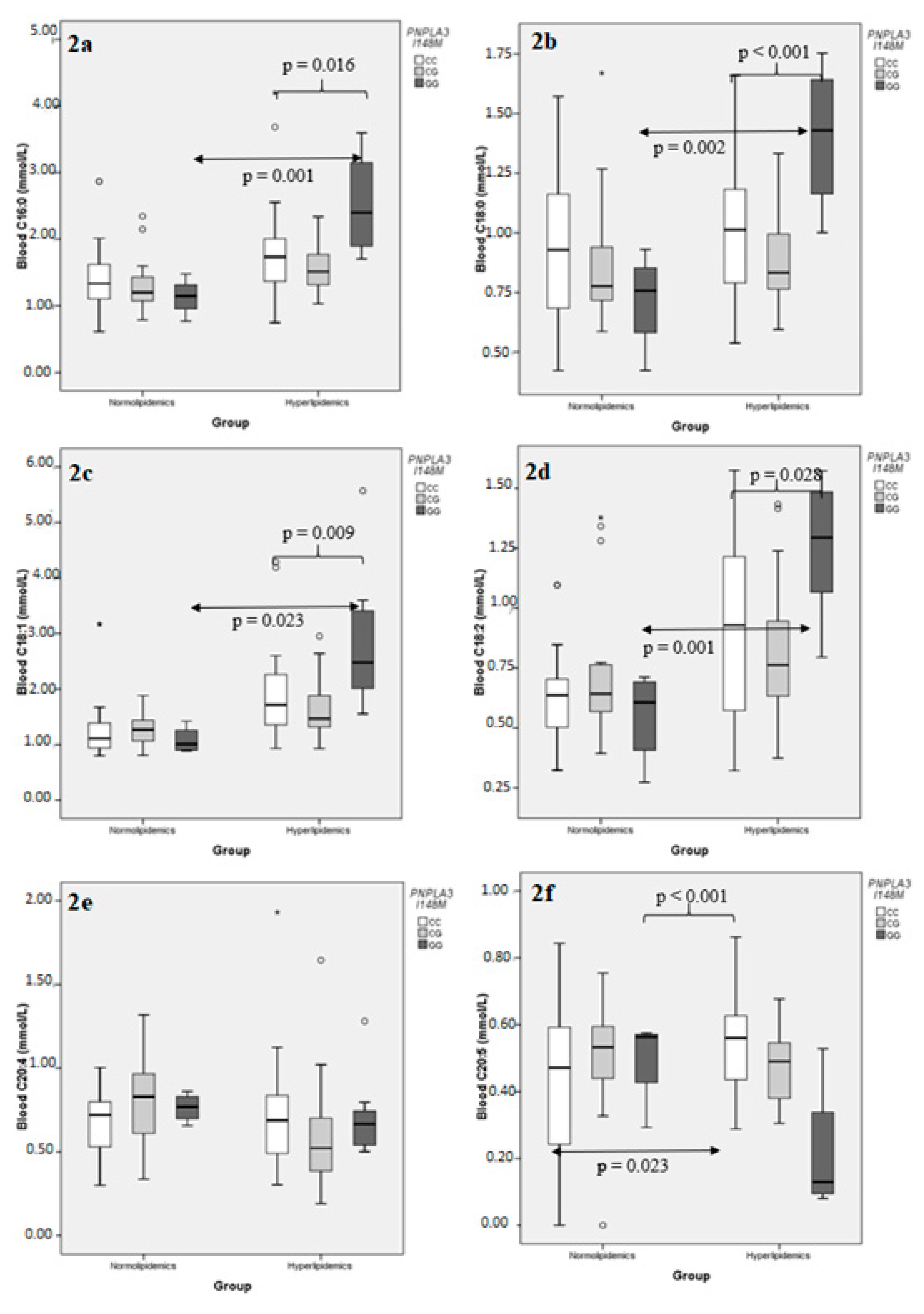
2.2. Association of FA Concentrations with ABCB1 G2677T and PNPLA3 I148M Genotypes
2.2.1. Total Blood FAs
2.2.2. Individual Blood FAs
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Biological Samples
4.3. Genotyping
4.4. FA Profiling
4.4.1. Sample Preparation Procedure
4.4.2. Gas Chromatographic Analysis
4.4.3. Method Validation
4.4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pingitore, P.; Romeo, S. The role of PNPLA3 in health and disease. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 900–906. [Google Scholar] [CrossRef]
- Mitsche, M.A.; Hobbs, H.H.; Cohen, J.C. Patatin-like phospholipase domain–containing protein 3 promotes transfers of essential fatty acids from triglycerides to phospholipids in hepatic lipid droplets. J. Biol. Chem. 2018, 293, 6958–6968. [Google Scholar] [CrossRef] [Green Version]
- Romeo, T.; Ferreti, F.; Consoli, P.; Canese, S.; Andaloro, F. Assessment of the Mediterranean Swordfish based on the Italian harpoon fishery data. ICES CM 2008/K 2008, 16. [Google Scholar] [CrossRef]
- BasuRay, S.; Smagris, E.; Cohen, J.C.; Hobbs, H.H. The PNPLA3 variant associated with fatty liver disease (I148M) accumulates on lipid droplets by evading ubiquitylation. Hepatology 2017, 66, 1111–1124. [Google Scholar] [CrossRef] [Green Version]
- Pirazzi, C.; Valenti, L.; Motta, B.M.; Pingitore, P.; Hedfalk, K.; Mancina, R.M.; Burza, M.A.; Indiveri, C.; Ferro, Y.; Montalcini, T.; et al. PNPLA3 has retinyl-palmitate lipase activity in human hepatic stellate cells. Hum. Mol. Genet. 2014, 23, 4077–4085. [Google Scholar] [CrossRef] [Green Version]
- Luukkonen, P.K.; Nick, A.; Hölttä-Vuori, M.; Thiele, C.; Isokuortti, E.; Lallukka-Brück, S.; Zhou, Y.; Hakkarainen, A.; Lundbom, N.; Peltonen, M.; et al. Human PNPLA3-I148M variant increases hepatic retention of polyunsaturated fatty acids. JCI Insight 2019, 4, e127902. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.D.; Krysko, D.V.; Verfaillie, T.; Kaczmarek, A.; Ferreira, G.B.; Marysael, T.; Rubio, N.; Firczuk, M.; Mathieu, C.; Roebroek, A.J.M.; et al. A novel pathway combining calreticulin exposure and ATP secretion in immunogenic cancer cell death. EMBO J. 2012, 31, 1062–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, L.M.; Murayama, M.; Larkum, M.E. Inhibitory Regulation of Dendritic Activity in vivo. Front. Neural Circuits 2012, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, M.; Sharma, R.; Mason, X.; DeBacker, J.; Jones, A.A.; Dumont, É.C. A Switch in the Neuromodulatory Effects of Dopamine in the Oval Bed Nucleus of the Stria Terminalis Associated with Cocaine Self-Administration in Rats. J. Neurosci. 2011, 31, 8928–8935. [Google Scholar] [CrossRef] [PubMed]
- Hyysalo, J.; Männistö, V.T.; Zhou, Y.; Arola, J.; Kärjä, V.; Leivonen, M.; Juuti, A.; Jaser, N.; Lallukka, S.; Käkelä, P.; et al. A population-based study on the prevalence of NASH using scores validated against liver histology. J. Hepatol. 2014, 60, 839–846. [Google Scholar] [CrossRef]
- Svendsen, K.; Olsen, T.; Nordstrand Rusvik, T.C.; Ulven, S.M.; Holven, K.B.; Retterstøl, K.; Telle-Hansen, V.H. Fatty acid profile and estimated desaturase activities in whole blood are associated with metabolic health. Lipids Heal. Dis. 2020, 19, 102. [Google Scholar] [CrossRef] [PubMed]
- Lankinen, M.; Schwab, U.; Kolehmainen, M.; Paananen, J.; Nygren, H.; Seppänen-Laakso, T.; Poutanen, K.; Hyötyläinen, T.; Risérus, U.; Savolainen, M.J.; et al. A Healthy Nordic Diet Alters the Plasma Lipidomic Profile in Adults with Features of Metabolic Syndrome in a Multicenter Randomized Dietary Intervention. J. Nutr. 2015, 146, 662–672. [Google Scholar] [CrossRef] [Green Version]
- Metherall, P.; Barber, D.C.; Smallwood, R.H.; Brown, B.H. Three-dimensional electrical impedance tomography. Nature 1996, 380, 509–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottova, I.; Hehl, A.B.; Štefanić, S.; Fabriàs, G.; Casas, J.; Schraner, E.; Pieters, J.; Sonda, S. Host Cell P-glycoprotein Is Essential for Cholesterol Uptake and Replication of Toxoplasma gondii. J. Biol. Chem. 2009, 284, 17438–17448. [Google Scholar] [CrossRef] [Green Version]
- Le May, C.; Berger, J.M.; Lespine, A.; Pillot, B.; Prieur, X.; Letessier, E.; Hussain, M.M.; Collet, X.; Cariou, B.; Costet, P. Transintestinal Cholesterol Excretion Is an Active Metabolic Process Modulated by PCSK9 and Statin Involving ABCB1. Arter. Thromb. Vasc. Biol. 2013, 33, 1484–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, J.; Rose-Sperling, D.; Hellmich, U.A. Diverse relations between ABC transporters and lipids: An overview. Biochim. Biophys. Acta (BBA) Biomembr. 2017, 1859, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Renaud, F.; Bournaud, F.; Duc, P.-A. A parsec-resolution simulation of the Antennae galaxies: Formation of star clusters during the merger. Mon. Not. R. Astron. Soc. 2015, 446, 2038–2054. [Google Scholar] [CrossRef] [Green Version]
- Foucaud-Vignault, M.; Soayfane, Z.; Ménez, C.; Bertrand-Michel, J.; Martin, P.G.P.; Guillou, H.; Collet, X.; Lespine, A. P-glycoprotein Dysfunction Contributes to Hepatic Steatosis and Obesity in Mice. PLoS ONE 2011, 6, e23614. [Google Scholar] [CrossRef] [Green Version]
- Agapakis, D.; Panderi, A.; Gbandi, E.; Savopoulos, C.; Kouvelas, D.; Hatzitolios, A.I.; Goulas, A. The ABCB1 2677G>T/A polymorphism is associated with baseline blood HDL-cholesterol levels in newly diagnosed hyperlipidemic patients. Hell. J. Cardiol. 2018, 59, 122–126. [Google Scholar] [CrossRef]
- Ichihara, K.; Fukubayashi, Y. Preparation of fatty acid methyl esters for gas-liquid chromatography. J. Lipid Res. 2010, 51, 635–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luukkonen, P.K.; Qadri, S.; Lehtimäki, T.E.; Juuti, A.; Sammalkorpi, H.; Penttilä, A.K.; Hakkarainen, A.; Orho-Melander, M.; Arola, J.; Yki-Järvinen, H. The PNPLA3-I148M Variant Confers an Antiatherogenic Lipid Profile in Insulin-resistant Patients. J. Clin. Endocrinol. Metab. 2021, 106, e300–e315. [Google Scholar] [CrossRef]
- Luukkonen, P.K.; Zhou, Y.; Sädevirta, S.; Leivonen, M.; Arola, J.; Orešič, M.; Hyötyläinen, T.; Yki-Järvinen, H. Hepatic ceramides dissociate steatosis and insulin resistance in patients with non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1167–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, E.A. The degradation of apolipoprotein B100: Multiple opportunities to regulate VLDL triglyceride production by different proteolytic pathways. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2012, 1821, 778–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, D.; Arias, I.M. Intracellular trafficking of P-glycoprotein. Int. J. Biochem. Cell Biol. 2012, 44, 461–464. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, A.; Joshua, V.; Hensvold, A.H.; Jin, T.; Sun, M.; Vivar, N.; Ytterberg, A.J.; Engström, M.; Fernandes-Cerqueira, C.; Amara, K.; et al. Identification of a novel chemokine-dependent molecular mechanism underlying rheumatoid arthritis-associated autoantibody-mediated bone loss. Ann. Rheum. Dis. 2016, 75, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wei, T.-T.; Li, Y.; Li, J.; Fan, Y.; Huang, F.-Q.; Cai, Y.-Y.; Ma, G.; Liu, J.-F.; Chen, Q.-Q.; et al. Functional Metabolomics Characterizes a Key Role for N-Acetylneuraminic Acid in Coronary Artery Diseases. Circulation 2018, 137, 1374–1390. [Google Scholar] [CrossRef]
- Gbandi, E.; Goulas, A.; Sevastianos, V.; Hadziyannis, S.; Panderi, A.; Koskinas, J.; Papatheodoridis, G.; Vasiliadis, T.; Agapakis, D.; Protopapas, A.; et al. Common ABCB1 polymorphisms in Greek patients with chronic hepatitis C infection: A comparison with hyperlipidemic patients and the general population. Pharmacol. Rep. 2016, 68, 476–482. [Google Scholar] [CrossRef]
- Falleti, E.; Fabris, C.; Cmet, S.; Cussigh, A.; Bitetto, D.; Fontanini, E.; Fornasiere, E.; Bignulin, S.; Fumolo, E.; Bignulin, E.; et al. PNPLA3 rs738409C/G polymorphism in cirrhosis: Relationship with the aetiology of liver disease and hepatocellular carcinoma occurrence. Liver Int. 2011, 31, 1137–1143. [Google Scholar] [CrossRef] [PubMed]
- US Department of Health and Human Services. Bioanalytical Method Validation Guidance for Industry; US Department of Health and Human Services: Washington, DC, USA, 2018; p. 44.




{kind=link}
{kind=link}
{kind=link}
| FA | Retention Time (min) | Linearity (mmol/L) | R2 | LOD (mmol/L) | LOQ (mmo/L) |
|---|---|---|---|---|---|
| C14:0 | 11.151 | 0.014–0.415 | 0.991 | 0.001 | 0.004 |
| C15:0 | 12.051 | 0.01–0.158 | 0.999 | 0.002 | 0.005 |
| C15:1 | 12.516 | 0.01–0.449 | 0.993 | 0.002 | 0.007 |
| C16:0 | 13.061 | 0.33–2.522 | 0.998 | 0.002 | 0.008 |
| C16:1 | 13.461 | 0.01–0.825 | 0.992 | 0.002 | 0.005 |
| C17:0 | 14.181 | 0.01–0.17 | 0.999 | 0.001 | 0.004 |
| C17:1 | 14.621 | 0.03–0.521 | 0.996 | 0.007 | 0.024 |
| C18:0 | 15.39 | 0.19–1.344 | 0.99 | 0.012 | 0.039 |
| C18:1cis | 15.772 | 0.32–2.95 | 0.991 | 0.034 | 0.112 |
| C18:2cis | 16.497 | 0.13–1.378 | 0.997 | 0.016 | 0.053 |
| C20:0 | 18.013 | 0.007–0.071 | 0.998 | 0.002 | 0.005 |
| C20:1n9 | 18.439 | 0.008–0.122 | 0.999 | 0.001 | 0.004 |
| C20:2 | 19.2 | 0.006–0.092 | 0.997 | 0.001 | 0.003 |
| C20:3n6 | 19.656 | 0.03–0.535 | 0.997 | 0.003 | 0.01 |
| C20:4n6 | 19.957 | 0.29–0.993 | 0.997 | 0.027 | 0.089 |
| C20:5 | 20.953 | 0.016–0.723 | 0.997 | 0.003 | 0.009 |
| C23:0 | 22.4 | 0.01–0.189 | 0.999 | 0.003 | 0.009 |
| C24:0 | 24.259 | 0.02–0.312 | 0.997 | 0.002 | 0.006 |
| C24:1 | 24.987 | 0.03–0.533 | 0.998 | 0.003 | 0.009 |
| C22:6 | 25.129 | 0.02–1.698 | 0.995 | 0.002 | 0.008 |
| Demographic Characteristics and Blood Lipid Parameters | Group | p1 | |
|---|---|---|---|
| Normolipidemics | Hyperlipidemics | ||
| n | 42 | 62 | |
| Age (years ± SD) | 51.7 ± 14.38 | 55.0 ± 13.75 | 0.237 |
| BMI (kg/m2 ± SD) | 25.7 ± 2.91 | 27.3 ± 2.90 | 0.009 |
| Sex (females, %) | 54.5 | 65.1 | 0.313 |
| Smoking (yes, %) | 9.5 | 1.6 | 0.155 |
| T2D 2 (yes, %) | 16.7 | 36.5 | 0.030 |
| HbA1c 3 (% ± SD) | NA 4 | 5.75 ± 0.927 | – |
| TC 5 (mmol/L ± SD) | 4.76 ± 0.689 | 6.53 ± 0.986 | <0.001 |
| TG 6 (mmol/L ± SD) | 1.14 ± 0.440 | 2.40 ± 1.029 | <0.001 |
| PNPLA3 I148M (n, %) | |||
| CC (II) | 21 (52.5) | 26 (42.6) | 0.616 |
| CG (IM) | 15 (37.5) | 27 (44.3) | |
| GG (MM) | 4 (10.0) | 8 (13.1) | |
| Deviation from Hardy–Weinberg equilibrium, p 7 | 0.864 | 0.842 | |
| ABCB1 G2677T (n, %) | |||
| GG | 15 (35.7) | 26 (41.9) | 0.807 |
| GT | 16 (38.1) | 22 (35.5) | |
| TT | 11 (26.2) | 14 (22.6) | |
| Deviation from Hardy–Weinberg equilibrium, p 7 | 0.327 | 0.115 | |
| FA (mmol/L ± SD) | Group | p1 | |
|---|---|---|---|
| Normolipidemics | Hyperlipidemics | ||
| C14:0 | 0.16 ± 0.079 | 0.25 ± 0.084 | <0.001 |
| C15:0 | 0.06 ± 0.021 | 0.08 ± 0.034 | 0.002 |
| C15:1 | 0.29 ± 0.092 | 0.27 ± 0.087 | 0.468 |
| C16:0 | 1.37 ± 0.487 | 1.81 ± 0.678 | <0.001 |
| C16:1 | 0.27 ± 0.209 | 0.45 ± 0.329 | <0.001 |
| C17:0 | 0.08 ± 0.024 | 0.10 ± 0.035 | 0.005 |
| C17:1 | 0.17 ± 0.061 | 0.16 ± 0.058 | 0.635 |
| C18:0 | 0.90 ± 0.307 | 1.01 ± 0.294 | 0.043 |
| C18:1 | 1.30 ± 0.524 | 1.89 ± 0.855 | <0.001 |
| C18:2 | 0.68 ± 0.252 | 0.91 ± 0.348 | 0.001 |
| C20:0 | 0.01 ± 0.003 | 0.01 ± 0.027 | 0.164 |
| C20:1n9 | 0.05 ± 0.018 | 0.06 ± 0.028 | 0.054 |
| C20:2 | 0.02 ± 0.023 | 0.03 ± 0.035 | 0.023 |
| C20:3n6 | 0.24 ± 0.080 | 0.31 ± 0.157 | 0.029 |
| C20:4n6 | 0.74 ± 0.229 | 0.65 ± 0.307 | 0.015 |
| C20:5 | 0.44 ± 0.238 | 0.46 ± 0.175 | 0.977 |
| C23:0 | 0.07 ± 0.019 | 0.08 ± 0.035 | 0.263 |
| C24:0 | 0.17 ± 0.045 | 0.16 ± 0.040 | 0.085 |
| C24:1 | 0.31 ± 0.045 | 0.28 ± 0.069 | 0.021 |
| C22:6 | 0.40 ± 0.945 | 0.34 ± 0.169 | 0.019 |
| Total | 7.72 ± 2.247 | 9.29 ± 2.970 | 0.003 |
| Genotype | Group | p1 | |
|---|---|---|---|
| PNPLA3 I148M | Normolipidemics (n) | Hyperlipidemics (n) | |
| CC (II) | 7.59 ± 2.463 (21) | 9.44 ± 3.374 (26) | 0.041 |
| CG (IM) | 8.08 ± 2.262 (15) | 8.25 ± 1.946 (27) | 0.795 |
| GG (MM) | 6.70 ± 1.421 (4) | 12.57 ± 2.541 (8) | 0.002 |
| p2 | 0.601 | 0.006 | |
| ABCB1 G2677T | |||
| GG | 8.23 ± 2.524 (15) | 8.42 ± 2.380 (26) | 0.817 |
| GT | 7.92 ± 2.470 (16) | 9.43 ± 2.577 (22) | 0.078 |
| TT | 6.73 ± 1.044 (11) | 10.78 ± 4.044 (14) | 0.003 |
| p2 | 0.282 | 0.056 | |
| FA | Adjusted R2 | ||
|---|---|---|---|
| PNPLA3 I148M (p) | +BMI (p) | +BMI + HbA1c (p) | |
| C16:0 | 0.145 (0.001) | 0.150 (0.259) | 0.171 (0.118) |
| C18:0 | 0.262 (<0.001) | 0.256 (0.470) | 0.276 (0.108) |
| C18:1 | 0.175 (<0.001) | 0.178 (0.277) | 0.226 (0.038) |
| C18:2 | 0.132 (0.002) | 0.164 (0.075) | 0.151 (0.754) |
| C20:4 | −0.011 (0.562) | −0.004 (0.422) | −0.013 (0.531) |
| C20:5 | 0.325 (<0.001) | 0.313 (0.921) | 0.304 (0.644) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mouskeftara, T.; Goulas, A.; Ioannidou, D.; Ntenti, C.; Agapakis, D.; Assimopoulou, A.; Gika, H. A Study of Blood Fatty Acids Profile in Hyperlipidemic and Normolipidemic Subjects in Association with Common PNPLA3 and ABCB1 Polymorphisms. Metabolites 2021, 11, 90. https://doi.org/10.3390/metabo11020090
Mouskeftara T, Goulas A, Ioannidou D, Ntenti C, Agapakis D, Assimopoulou A, Gika H. A Study of Blood Fatty Acids Profile in Hyperlipidemic and Normolipidemic Subjects in Association with Common PNPLA3 and ABCB1 Polymorphisms. Metabolites. 2021; 11(2):90. https://doi.org/10.3390/metabo11020090
Chicago/Turabian StyleMouskeftara, Thomai, Antonis Goulas, Despoina Ioannidou, Charikleia Ntenti, Dimitris Agapakis, Andreana Assimopoulou, and Helen Gika. 2021. "A Study of Blood Fatty Acids Profile in Hyperlipidemic and Normolipidemic Subjects in Association with Common PNPLA3 and ABCB1 Polymorphisms" Metabolites 11, no. 2: 90. https://doi.org/10.3390/metabo11020090