
Identification of Quinone Degradation as a Triggering Event for Intense Pulsed Light-Elicited Metabolic Changes in Escherichia coli by Metabolomic Fingerprinting

 and
and 
Abstract
:1. Introduction
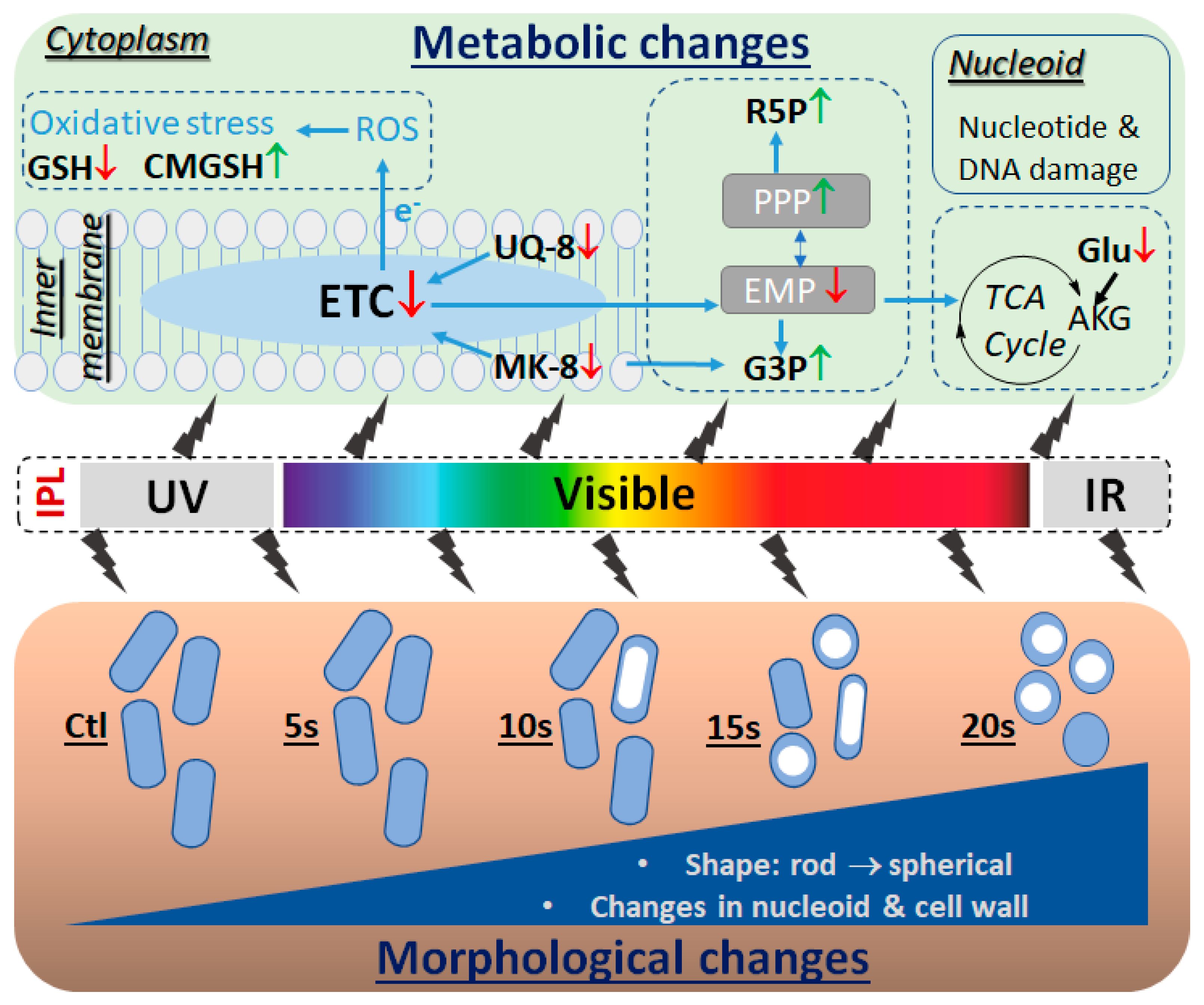
2. Results
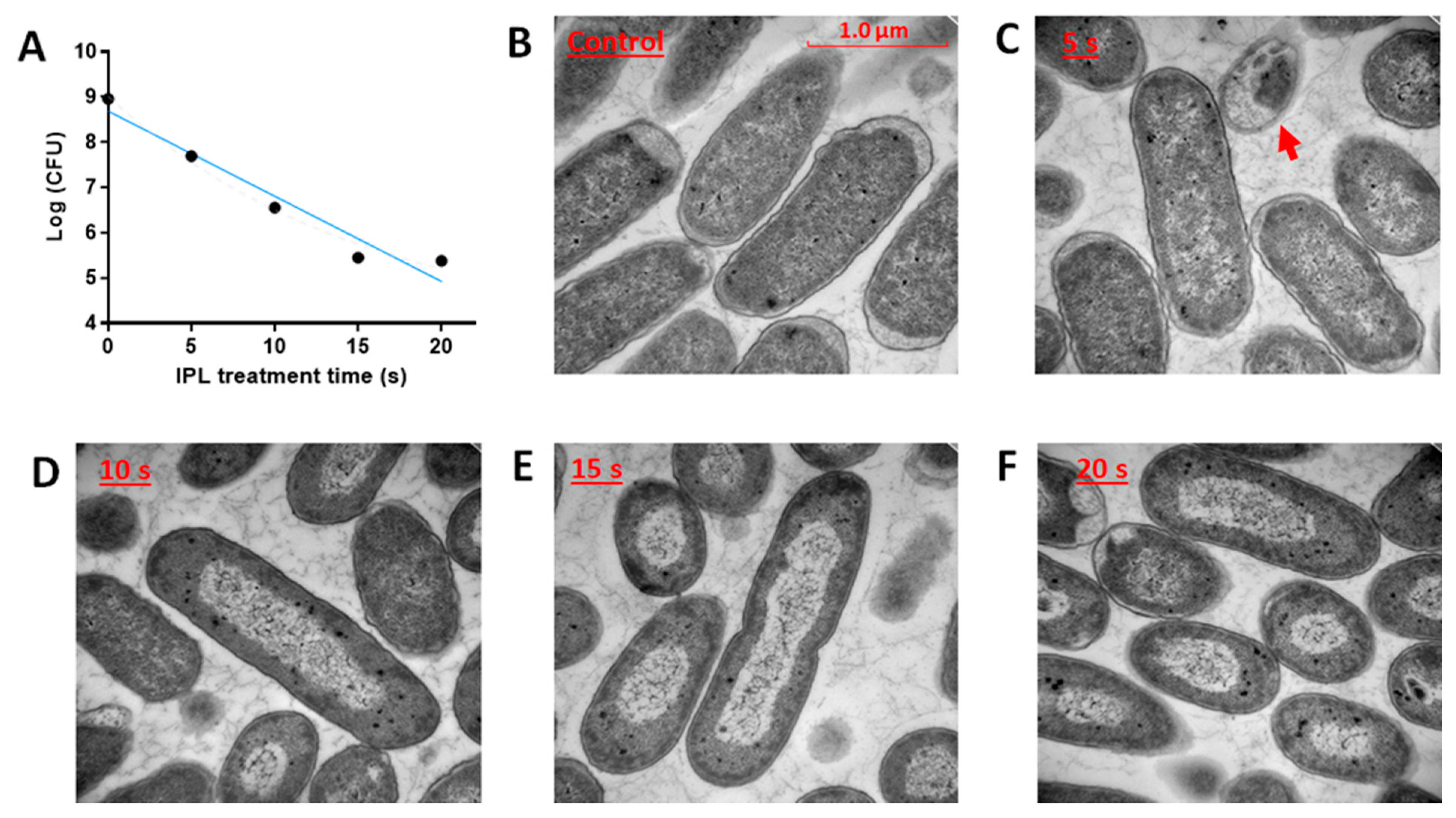
2.1. Effects of IPL on E. coli Growth and Morphology
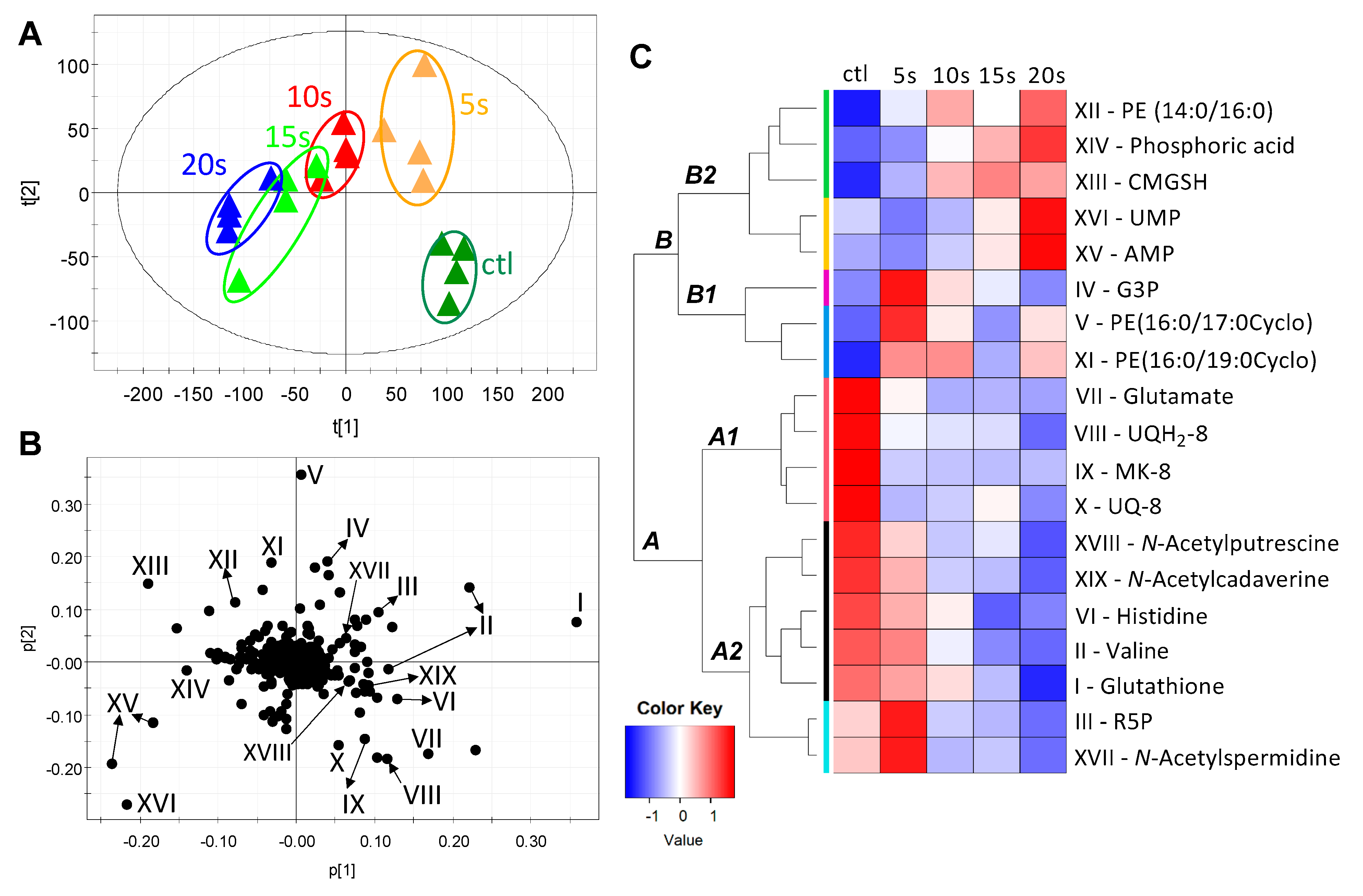
2.2. Modeling and Identification of IPL-Induced Changes in E. coli Metabolome
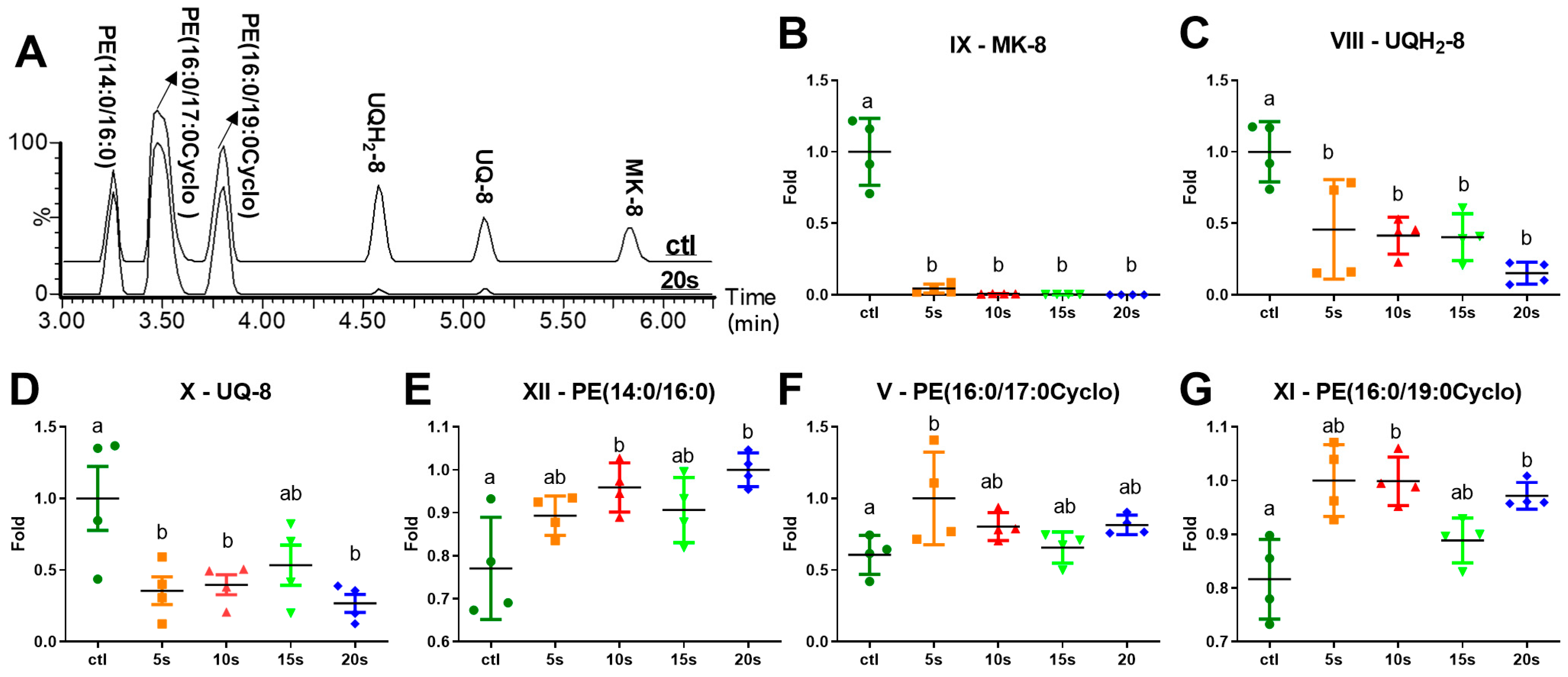
2.3. Influences of IPL Treatment on Membrane Lipids
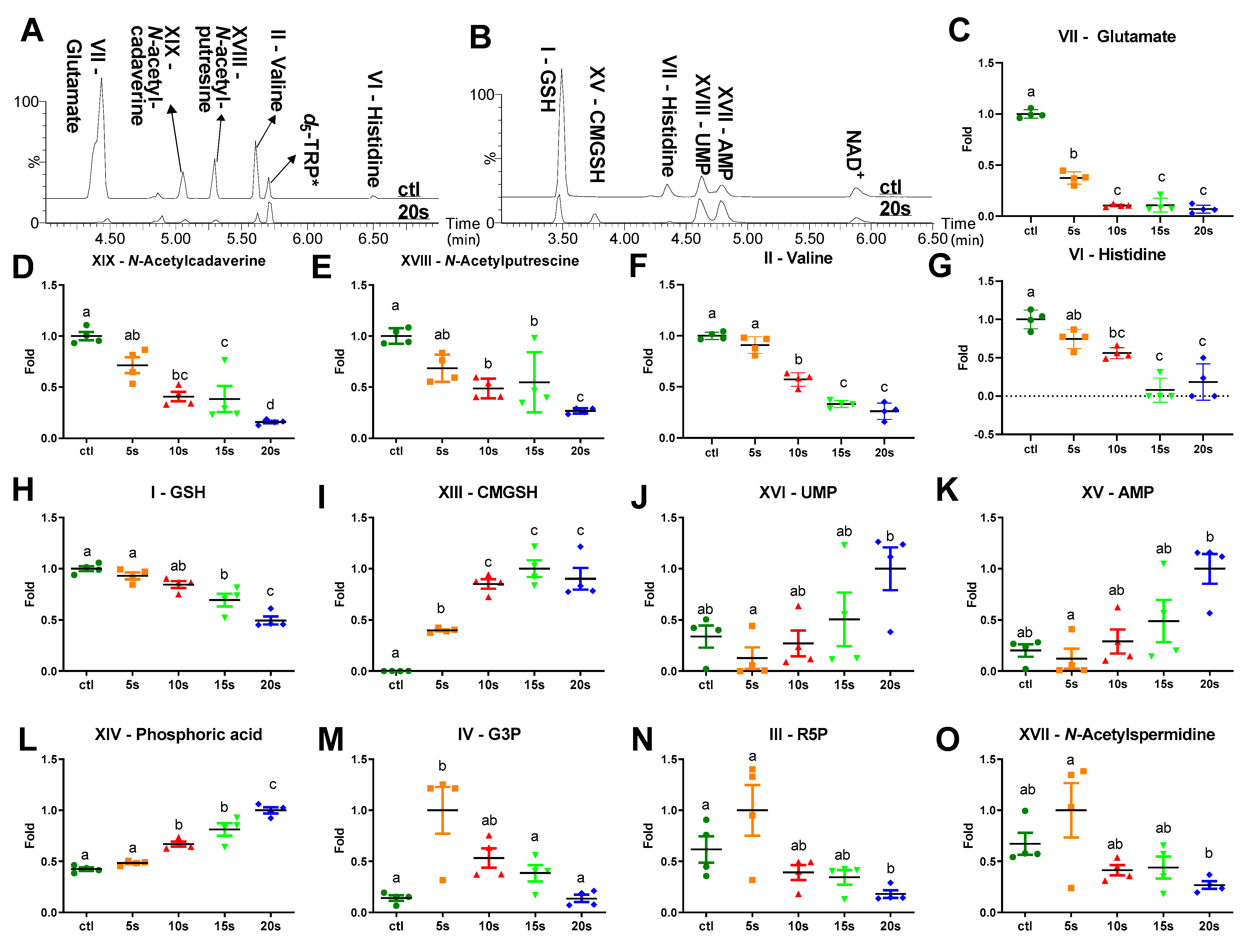
2.4. Influences of IPL Treatment on Cytoplasmic Metabolites
3. Discussion
3.1. Degradation of Quinone Electron Carriers
3.2. Direct Stress Responses
3.3. Disruption of Intermediary Metabolism
3.4. Nucleotide Degradation and Morphological Changes
3.5. Potential Issues Related to the IPL Application in Food Disinfection
4. Materials and Methods
4.1. Culture of E. coli
4.2. Chemicals
4.3. IPL Treatment of E. coli
4.4. Counting of Surviving Cells
4.5. Morphological Analysis
4.6. Sample Preparation for LC-MS Analysis
4.7. Chemical Derivatization
4.8. Synthesis of S-carboxymethyl-glutathione (CMGSH)
4.9. LC-MS Analysis
4.10. Multivariate Data Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bélanger, L.; Garenaux, A.; Harel, J.; Boulianne, M.; Nadeau, E.; Dozois, C.M. Escherichia coli from animal reservoirs as a potential source of human extraintestinal pathogenic E. coli. Pathog. Dis. 2011, 62, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kousta, M.; Mataragas, M.; Skandamis, P.; Drosinos, E.H. Prevalence and sources of cheese contamination with pathogens at farm and processing levels. Food Control 2010, 21, 805–815. [Google Scholar] [CrossRef]
- Heiman, K.E.; Mody, R.K.; Johnson, S.D.; Griffin, P.M.; Gould, L.H. Escherichia coli O157 Outbreaks in the United States, 2003–2012. Emerg. Infect Dis. 2015, 21, 1293. [Google Scholar] [CrossRef]
- Groh, C.D.; MacPherson, D.W.; Groves, D.J. Effect of heat on the sterilization of artificially contaminated water. J. Travel Med. 1996, 3, 11–13. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Singh, A.K. Effect of processing on nutritive values of milk protein. Crit. Rev. Food Sci. Nutr. 2017, 57, 3690–3702. [Google Scholar] [CrossRef]
- Li, M.; Qu, J.H.; Peng, Y.Z. Sterilization of Escherichia coli cells by the application of pulsed magnetic field. J. Environ. Sci. (China) 2004, 16, 348–352. [Google Scholar]
- Hong, Y.C.; Park, H.J.; Lee, B.J.; Kang, W.-S.; Uhm, H.S. Plasma formation using a capillary discharge in water and its application to the sterilization of E. coli. Phys. Plasmas 2010, 17, 053502. [Google Scholar] [CrossRef]
- Laroussi, M.; Sayler, G.S.; Glascock, B.B.; McCurdy, B.; Pearce, M.E.; Bright, N.G.; Malott, C.M. Images of biological samples undergoing sterilization by a glow discharge at atmospheric pressure. IEEE Trans. Plasma Sci. 1999, 27, 34–35. [Google Scholar] [CrossRef]
- Kamihira, M.; Taniguchi, M.; Kobayashi, T. Sterilization of microorganisms with supercritical carbon dioxide. Agric. Biol. Chem. 1987, 51, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Bucheli-Witschel, M.; Bassin, C.; Egli, T. UV-C inactivation in Escherichia coli is affected by growth conditions preceding irradiation, in particular by the specific growth rate. J. Appl. Microbiol. 2010, 109, 1733–1744. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wiertzema, J.; Peng, P.; Cheng, Y.; Liu, J.; Mao, Q.; Ma, Y.; Anderson, E.; Chen, P.; Baumler, D.J.; et al. Effects of intense pulsed light on Cronobacter sakazakii inoculated in non-fat dry milk. J. Food Eng. 2018, 238, 178–187. [Google Scholar] [CrossRef]
- Chen, D.; Mosher, W.; Wiertzema, J.; Peng, P.; Min, M.; Cheng, Y.; An, J.; Ma, Y.; Fan, X.; Niemira, B.A.; et al. Effects of intense pulsed light and gamma irradiation on Bacillus cereus spores in mesquite pod flour. Food Chem. 2021, 344, 128675. [Google Scholar] [CrossRef] [PubMed]
- Elmnasser, N.; Guillou, S.; Leroi, F.; Orange, N.; Bakhrouf, A.; Federighi, M. Pulsed-light system as a novel food decontamination technology: A review. Can. J. Microbiol. 2007, 53, 813–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshita, K.; Shibato, J.; Sameshima, T.; Fukunaga, S.; Isobe, S.; Arihara, K.; Itoh, M. Damage of yeast cells induced by pulsed light irradiation. Int. J. Food Microbiol. 2003, 85, 151–158. [Google Scholar] [CrossRef]
- Kramer, B.; Muranyi, P. Effect of pulsed light on structural and physiological properties of Listeria innocua and Escherichia coli. J. Appl. Microbiol. 2014, 116, 596–611. [Google Scholar] [CrossRef]
- Cheigh, C.; Park, M.; Chung, M.; Shin, J.; Park, Y. Comparison of intense pulsed light-and ultraviolet (UVC)-induced cell damage in Listeria monocytogenes and Escherichia coli O157: H7. Food Control 2012, 25, 654–659. [Google Scholar] [CrossRef]
- Mashego, M.R.; Rumbold, K.; De Mey, M.; Vandamme, E.; Soetaert, W.; Heijnen, J.J. Microbial metabolomics: Past, present and future methodologies. Biotechnol. Lett. 2007, 29, 1–16. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, L.; Laserna, A.K.C.; He, Y.; Feng, X.; Yang, H. Synergistic action of electrolyzed water and mild heat for enhanced microbial inactivation of Escherichia coli O157:H7 revealed by metabolomics analysis. Food Control 2020, 110, 107026. [Google Scholar] [CrossRef]
- Ahmed, A.E.-S.I.; Wardell, J.N.; Thumser, A.E.; Avignone-Rossa, C.A.; Cavalli, G.; Hay, J.N.; Bushell, M.E. Metabolomic profiling can differentiate between bactericidal effects of free and polymer bound halogen. J. Appl. Polym. Sci. 2011, 119, 709–718. [Google Scholar] [CrossRef] [Green Version]
- Planchon, M.; Léger, T.; Spalla, O.; Huber, G.; Ferrari, R. Metabolomic and proteomic investigations of impacts of titanium dioxide nanoparticles on Escherichia coli. PLoS ONE 2017, 12, e0178437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowicka, B.; Kruk, J. Occurrence, biosynthesis and function of isoprenoid quinones. Biochim. Biophys. Acta (BBA)—Bioenerg. 2010, 1797, 1587–1605. [Google Scholar] [CrossRef] [Green Version]
- Aussel, L.; Pierrel, F.; Loiseau, L.; Lombard, M.; Fontecave, M.; Barras, F. Biosynthesis and physiology of coenzyme Q in bacteria. Biochim. Biophys. Acta (BBA)—Bioenerg. 2014, 1837, 1004–1011. [Google Scholar] [CrossRef]
- Shokri, A.; Larsson, G. Characterisation of the Escherichia coli membrane structure and function during fedbatch cultivation. Microb. Cell Factories 2004, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-H.; Zeng, X.-A.; Wang, M.-S.; Brennan, C.S.; Gong, D. Modification of membrane properties and fatty acids biosynthesis-related genes in Escherichia coli and Staphylococcus aureus: Implications for the antibacterial mechanism of naringenin. Biochim. Biophys. Acta (Bba)—Biomembr. 2018, 1860, 481–490. [Google Scholar] [CrossRef]
- Madden, J.J.; Boatwright, D.T.; Jagger, J. Action spectra for modification of Escherichia Coli B/r menaquinone by near-ultraviolet and visible radiations (313–578 nm). Photochem. Photobiol. 1981, 33, 305–311. [Google Scholar] [CrossRef]
- Werbin, H.; Lakchaura, B.D.; Jagger, J. Near-ultraviolet modification of Escherichia Coli B Ubiquinone in vivo and in vitro. Photochem. Photobiol. 1974, 19, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Søballe, B.; Poole, R.K. Microbial ubiquinones: Multiple roles in respiration, gene regulation and oxidative stress management. Microbiology 1999, 145, 1817–1830. [Google Scholar] [CrossRef] [Green Version]
- Oms-Oliu, G.; Martín-Belloso, O.; Soliva-Fortuny, R. Pulsed light treatments for food preservation. A review. Food Bioprocess Technol. 2008, 3, 13. [Google Scholar] [CrossRef]
- Allocati, N.; Federici, L.; Masulli, M.; Di Ilio, C. Glutathione transferases in bacteria. FEBS J. 2009, 276, 58–75. [Google Scholar] [CrossRef] [PubMed]
- Elmore, M.J.; Lamb, A.J.; Ritchie, G.Y.; Douglas, R.M.; Munro, A.; Gajewska, A.; Booth, I.R. Activation potassium efflux from Escherichia coli by glutathione metabolites. Mol. Microbiol. 1990, 4, 405–412. [Google Scholar] [CrossRef]
- Carper, S.; Willis, D.; Manning, K.; Gerner, E. Spermidine acetylation in response to a variety of stresses in Escherichia coli. J. Biol. Chem. 1991, 266, 12439–12441. [Google Scholar] [CrossRef]
- Peekhaus, N.; Conway, T. What’s for Dinner?: Entner-Doudoroff Metabolism in Escherichia coli. J. Bacteriol. 1998, 180, 3495–3502. [Google Scholar] [CrossRef] [Green Version]
- Varga, M.E.; Weiner, J.H. Physiological role of GlpB of anaerobic glycerol-3-phosphate dehydrogenase of Escherichia coli. Biochem. Cell Biol. 1995, 73, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Kuehne, A.; Emmert, H.; Soehle, J.; Winnefeld, M.; Fischer, F.; Wenck, H.; Gallinat, S.; Terstegen, L.; Lucius, R.; Hildebrand, J. Acute activation of oxidative pentose phosphate pathway as first-line response to oxidative stress in human skin cells. Mol. Cell 2015, 59, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Christodoulou, D.; Link, H.; Fuhrer, T.; Kochanowski, K.; Gerosa, L.; Sauer, U. Reserve Flux Capacity in the Pentose Phosphate Pathway Enables Escherichia coli’s Rapid Response to Oxidative Stress. Cell Syst. 2018, 6, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reitzer, L. Nitrogen assimilation and global regulation in Escherichia coli. Annu. Rev. Microbiol. 2003, 57, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Mandelstam, J. The free amino acids in growing and non-growing populations of Escherichia coli. Biochem. J. 1958, 69, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Birsoy, K.; Wang, T.; Chen, W.W.; Freinkman, E.; Abu-Remaileh, M.; Sabatini, D.M. An Essential Role of the Mitochondrial Electron Transport Chain in Cell Proliferation Is to Enable Aspartate Synthesis. Cell 2015, 162, 540–551. [Google Scholar] [CrossRef] [Green Version]
- Malecki, M.; Kamrad, S.; Ralser, M.; Bähler, J. Mitochondrial respiration is required to provide amino acids during fermentative proliferation of fission yeast. EMBO Rep. 2020, 21, e50845. [Google Scholar] [CrossRef] [PubMed]
- Marcus, M.; Halpern, Y.S. The metabolic pathway of glutamate in Escherichia coli K-12. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1969, 177, 314–320. [Google Scholar] [CrossRef]
- Plaitakis, A.; Kalef-Ezra, E.; Kotzamani, D.; Zaganas, I.; Spanaki, C. The Glutamate Dehydrogenase Pathway and Its Roles in Cell and Tissue Biology in Health and Disease. Biology 2017, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemire, J.; Milandu, Y.; Auger, C.; Bignucolo, A.; Appanna, V.P.; Appanna, V.D. Histidine is a source of the antioxidant, α-ketoglutarate, in Pseudomonas fluorescens challenged by oxidative stress. FEMS Microbiol. Lett. 2010, 309, 170–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mailloux, R.J.; Singh, R.; Brewer, G.; Auger, C.; Lemire, J.; Appanna, V.D. α-Ketoglutarate Dehydrogenase and Glutamate Dehydrogenase Work in Tandem To Modulate the Antioxidant α-Ketoglutarate during Oxidative Stress in Pseudomonas fluorescens. J. Bacteriol. 2009, 191, 3804–3810. [Google Scholar] [CrossRef] [Green Version]
- Legendre, F.; Maclean, A.; Appanna, V.P.; Appanna, V.D. Biochemical pathways to α-ketoglutarate, a multi-faceted metabolite. World J. Microbiol. Biotechnol. 2020, 36, 123. [Google Scholar] [CrossRef]
- Chai, Q.; Singh, B.; Peisker, K.; Metzendorf, N.; Ge, X.; Dasgupta, S.; Sanyal, S. Organization of Ribosomes and Nucleoids in Escherichia coli Cells during Growth and in Quiescence. J. Biol. Chem. 2014, 289, 11342–11352. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Bi, X.; Hao, Y.; Liao, X. Induction of Viable but Nonculturable Escherichia coli O157:H7 by High Pressure CO2 and Its Characteristics. PLoS ONE 2013, 8, e62388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signoretto, C.; Lleò, M.D.M.; Canepari, P. Modification of the Peptidoglycan of Escherichia coli in the Viable But Nonculturable State. Curr. Microbiol. 2002, 44, 125–131. [Google Scholar] [CrossRef]
- Wei, C.; Zhao, X. Induction of Viable but Nonculturable Escherichia coli O157:H7 by Low Temperature and Its Resuscitation. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Wang, Y.; An, H.; Hao, Y.; Hu, X.; Liao, X. New Insights into the Formation of Viable but Nonculturable Escherichia coli O157:H7 Induced by High-Pressure CO2. mBio 2016, 7, e00961-16. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.D. Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 2010, 34, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Kramer, B.; Wunderlich, J.; Muranyi, P. Recent findings in pulsed light disinfection. J. Appl. Microbiol. 2017, 122, 830–856. [Google Scholar] [CrossRef] [Green Version]
- Abedi-Moghaddam, N.; Bulic, A.; Herderson, L.; Lam, E. Survival of Escherichia coli to UV irradiation during exponential and stationary phases of growth. J. Exp. Microbiol. Immunol 2004, 5, 44–49. [Google Scholar]
- Szenk, M.; Dill, K.A.; De Graff, A.M.R. Why Do Fast-Growing Bacteria Enter Overflow Metabolism? Testing the Membrane Real Estate Hypothesis. Cell Syst. 2017, 5, 95–104. [Google Scholar] [CrossRef]
- Murli, S.; Opperman, T.; Smith, B.T.; Walker, G.C. A Role for the umuDC Gene Products of Escherichia coli in Increasing Resistance to DNA Damage in Stationary Phase by Inhibiting the Transition to Exponential Growth. J. Bacteriol. 2000, 182, 1127–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, T.K.; Knabel, S.J.; Kwan, B.W. Bacterial Persister Cell Formation and Dormancy. Appl Environ. Microbiol. 2013, 79, 7116–7121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, D.; Schultz, J.; Matin, A. Starvation-induced cross protection against heat or H2O2 challenge in Escherichia coli. J. Bacteriol. 1988, 170, 3910–3914. [Google Scholar] [CrossRef] [Green Version]
- Schraft, H.; Watterworth, L.A. Enumeration of heterotrophs, fecal coliforms and Escherichia coli in water: Comparison of 3M Petrifilm plates with standard plating procedures. J. Microbiol. Methods 2005, 60, 335–342. [Google Scholar] [CrossRef]
- Loughlin, A.F.; Skiles, G.L.; Alberts, D.W.; Schaefer, W.H. An ion exchange liquid chromatography/mass spectrometry method for the determination of reduced and oxidized glutathione and glutathione conjugates in hepatocytes. J. Pharm. Biomed. Anal. 2001, 26, 131–142. [Google Scholar] [CrossRef]
- Ma, Y.; Zhou, W.; Chen, P.; Urriola, P.E.; Shurson, G.; Ruan, R.; Chen, C. Metabolomic Evaluation of Scenedesmus sp. as a Feed Ingredient Revealed Dose-Dependent Effects on Redox Balance, Intermediary and Microbial Metabolism in a Mouse Model. Nutrients 2019, 11, 1971. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yao, D.; Urriola, P.E.; Hanson, A.R.; Saqui-Salces, M.; Kerr, B.J.; Shurson, G.C.; Chen, C. Identification of activation of tryptophan–NAD+ pathway as a prominent metabolic response to thermally oxidized oil through metabolomics-guided biochemical analysis. J. Nutr. Biochem. 2018, 57, 255–267. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ions | Modes of Ion Detection | Detected m/z | Identity | Formula | Mass Deviation (ppm) | KEGG ID |
|---|---|---|---|---|---|---|
| I | [M − H]− | 306.0752 | Glutathione * | C10H17N3O6S | −2.61 | C00051 |
| II | [M + H]+ | 118.0867 | Valine * | C5H11NO2 | −0.85 | C00183 |
| II | [M + DC]+ | 351.1369 | Valine * | C5H11NO2 | −2.85 | C00183 |
| III | [M − H]- | 229.0111 | Ribose 5-phosphate * | C5H11O8P | −0.87 | C00117 |
| IV | [M − H]- | 171.0041 | Glycerol 3-phosphate * | C3H9O6P | −0.58 | C00093 |
| V | [M + H]+ | 704.5238 | PE(16:0/17:0Cyclo) #,† | C38H74NO8P | 1.14 | NA |
| VI | [M − H]- | 154.062 | Histidine * | C6H9N3O2 | 1.95 | C00135 |
| VII | [M + DC]+ | 381.1111 | Glutamate * | C5H9NO4 | −0.26 | C00025 |
| VIII | [M + NH4]+ | 746.6103 | Ubiquinol-8 # | C49H76O4 | 2.01 | C00390 |
| IX | [M + NH4]+ | 734.5888 | Menaquinone-8 # | C51H72O2 | 1.63 | C00828 |
| X | [M + H]+ | 727.5684 | Ubiquinone-8 # | C49H74O4 | 2.61 | C17569 |
| XI | [M + H]+ | 732.5553 | PE(16:0/19:0Cyclo) #,† | C40H78NO8P | 1.37 | NA |
| XII | [M + H]+ | 664.4928 | PE(14:0/16:0) #,† | C35H70NO8P | 1.66 | C00350 |
| XIII | [M − H]− | 364.0802 | S-carboxymethyl-glutathione * | C12H19N3O8S | −3.57 | C14862 |
| XIV | [M + H]+ | 98.9848 | Phosphoric acid * | H3PO4 | 1.01 | C00009 |
| XV | [M − H]− | 346.0543 | Adenosine monophosphate * | C10H14N5O7P | −2.89 | C00020 |
| XV | [M + H]+ | 348.0701 | Adenosine monophosphate * | C10H14N5O7P | −2.30 | C00020 |
| XVI | [M − H]- | 323.0269 | Uridine monophosphate * | C9H13N2O9P | −3.41 | C00105 |
| XVII | [M + H]+ | 188.1761 | N-Acetylspermidine # | C9H21N3O | −1.06 | C01029 |
| XVIII | [M + DC]+ | 364.1688 | N-Acetylputrescine * | C6H14N2O | −1.92 | C02714 |
| XIX | [M + DC]+ | 378.1845 | N-Acetylcadaverine # | C7H16N2O | −1.85 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, Q.; Liu, J.; Wiertzema, J.R.; Chen, D.; Chen, P.; Baumler, D.J.; Ruan, R.; Chen, C. Identification of Quinone Degradation as a Triggering Event for Intense Pulsed Light-Elicited Metabolic Changes in Escherichia coli by Metabolomic Fingerprinting. Metabolites 2021, 11, 102. https://doi.org/10.3390/metabo11020102
Mao Q, Liu J, Wiertzema JR, Chen D, Chen P, Baumler DJ, Ruan R, Chen C. Identification of Quinone Degradation as a Triggering Event for Intense Pulsed Light-Elicited Metabolic Changes in Escherichia coli by Metabolomic Fingerprinting. Metabolites. 2021; 11(2):102. https://doi.org/10.3390/metabo11020102
Chicago/Turabian StyleMao, Qingqing, Juer Liu, Justin R. Wiertzema, Dongjie Chen, Paul Chen, David J. Baumler, Roger Ruan, and Chi Chen. 2021. "Identification of Quinone Degradation as a Triggering Event for Intense Pulsed Light-Elicited Metabolic Changes in Escherichia coli by Metabolomic Fingerprinting" Metabolites 11, no. 2: 102. https://doi.org/10.3390/metabo11020102