
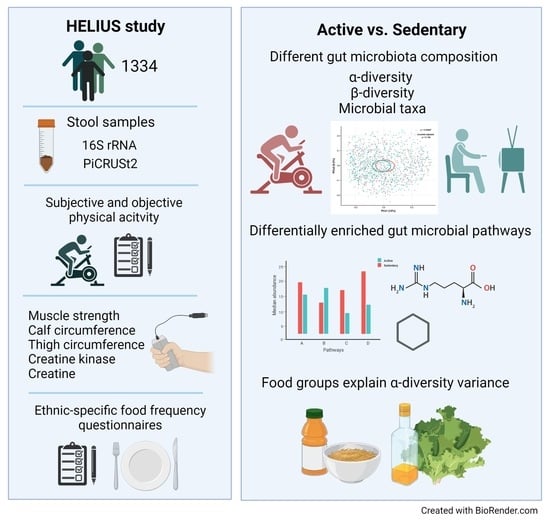
Physical Activity and Dietary Composition Relate to Differences in Gut Microbial Patterns in a Multi-Ethnic Cohort—The HELIUS Study

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Characterization of the Study Population by Physical Activity Level
2.2. Dietary Intake in Relation to Physical Activity
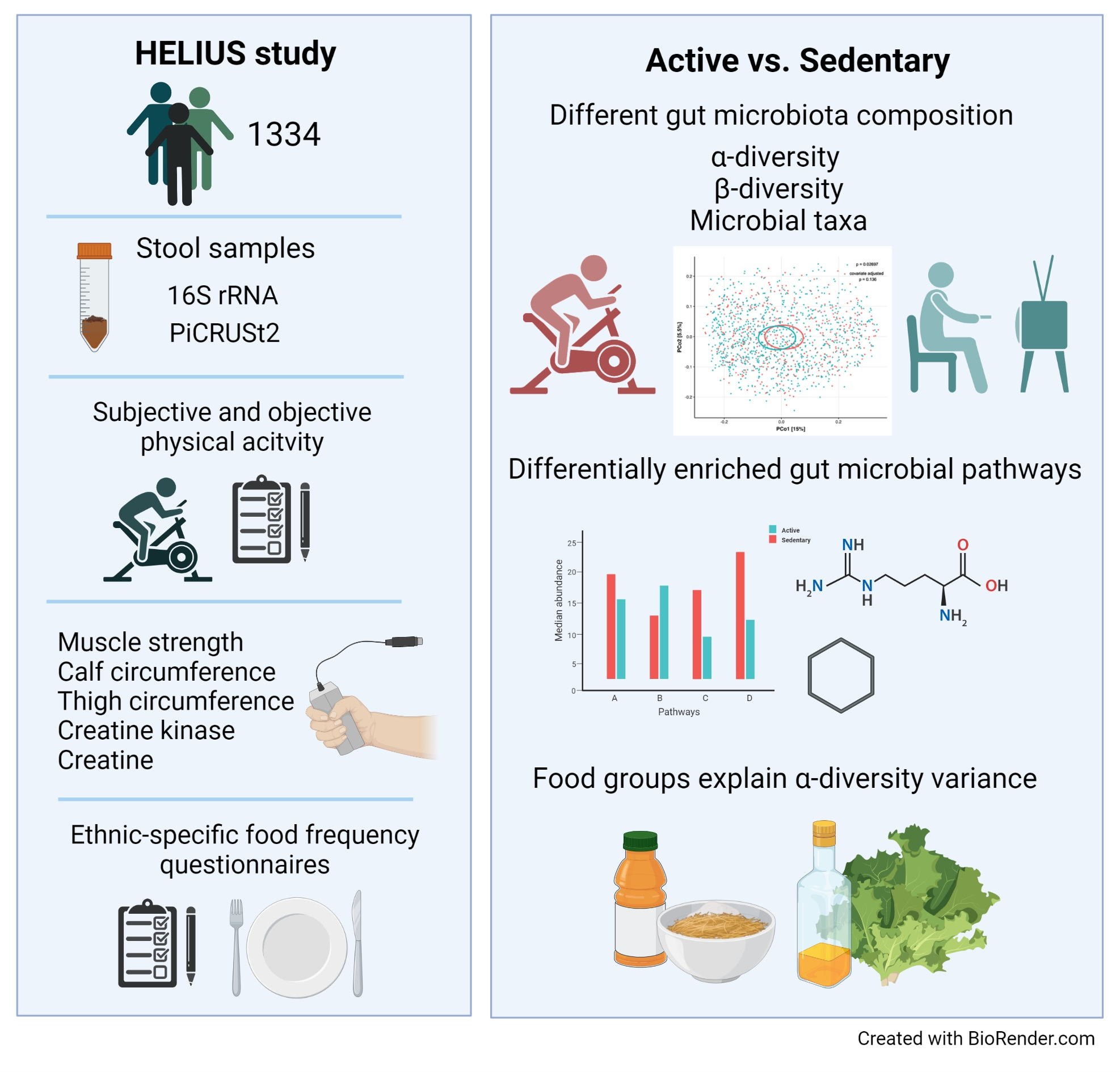
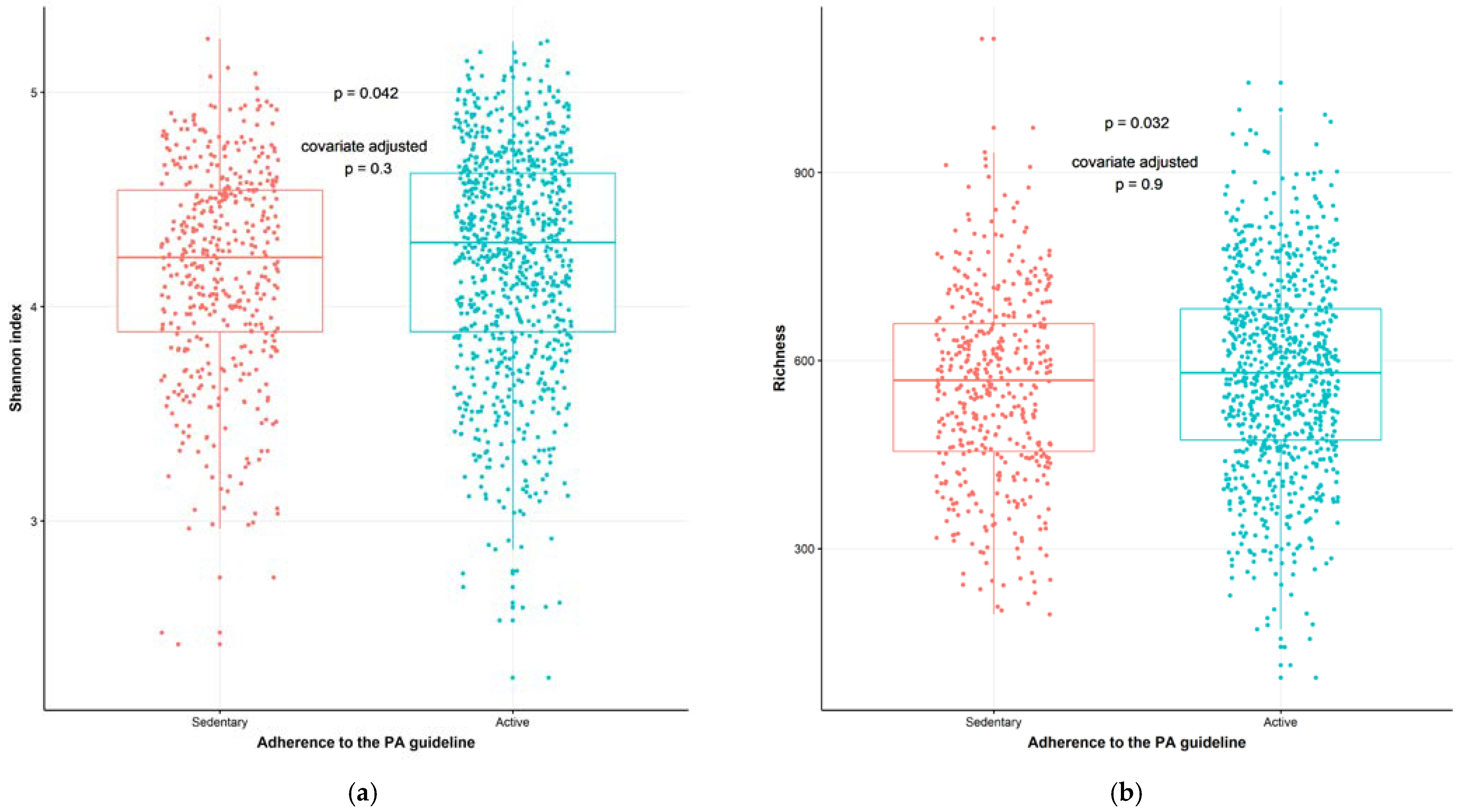
2.3. Physical Activity Associates with Gut Microbiata Composition
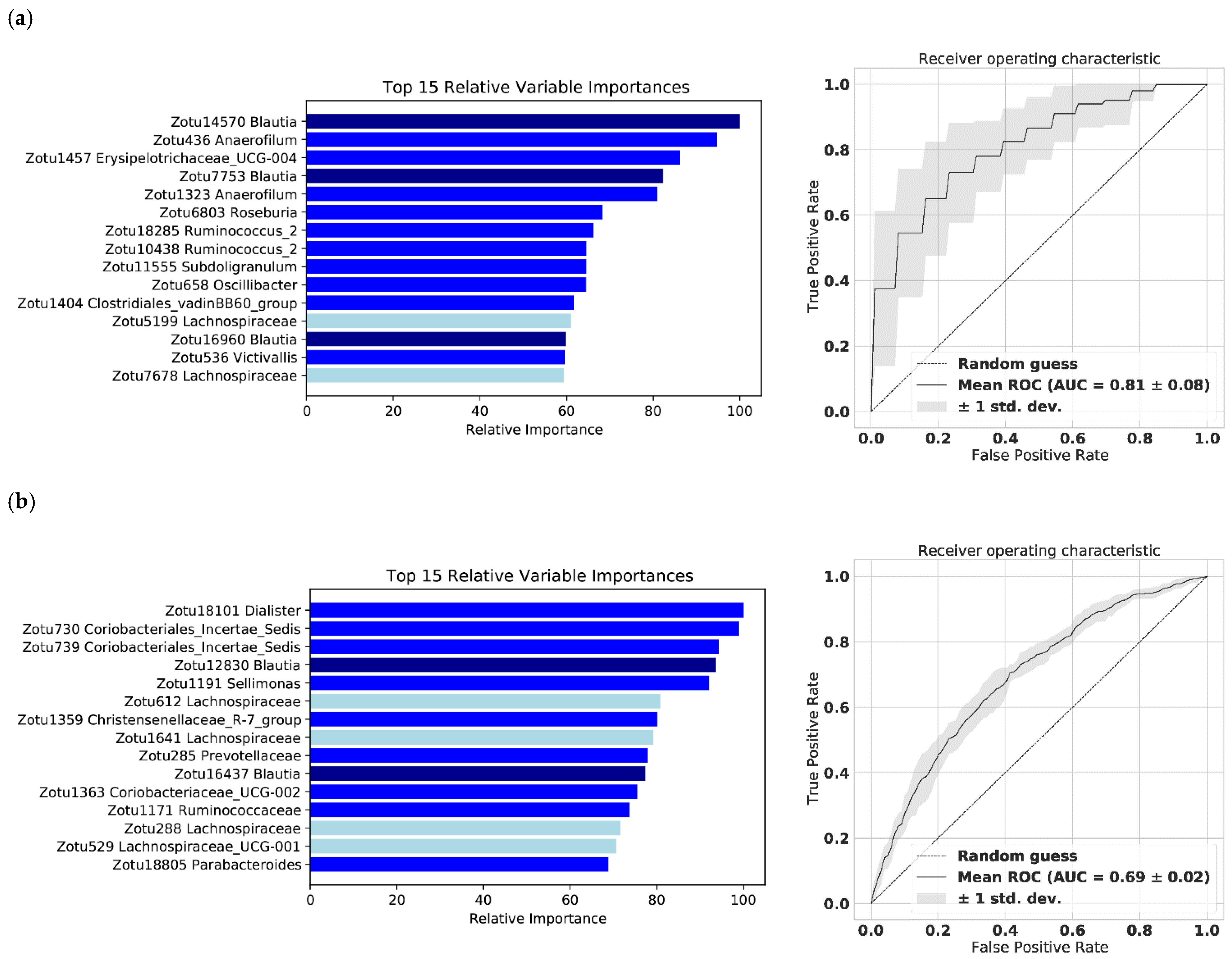
2.4. The Gut Microbiome Predicts Subjectively and Objectively Monitored Physical Activity—A Machine Learning Model
2.5. Parameters Related to Physical Fitness Associate with Variance of the Gut Microbiome
2.6. Functionality of the Gut Microbiome in Relation to Physical Activity and Related Parameters
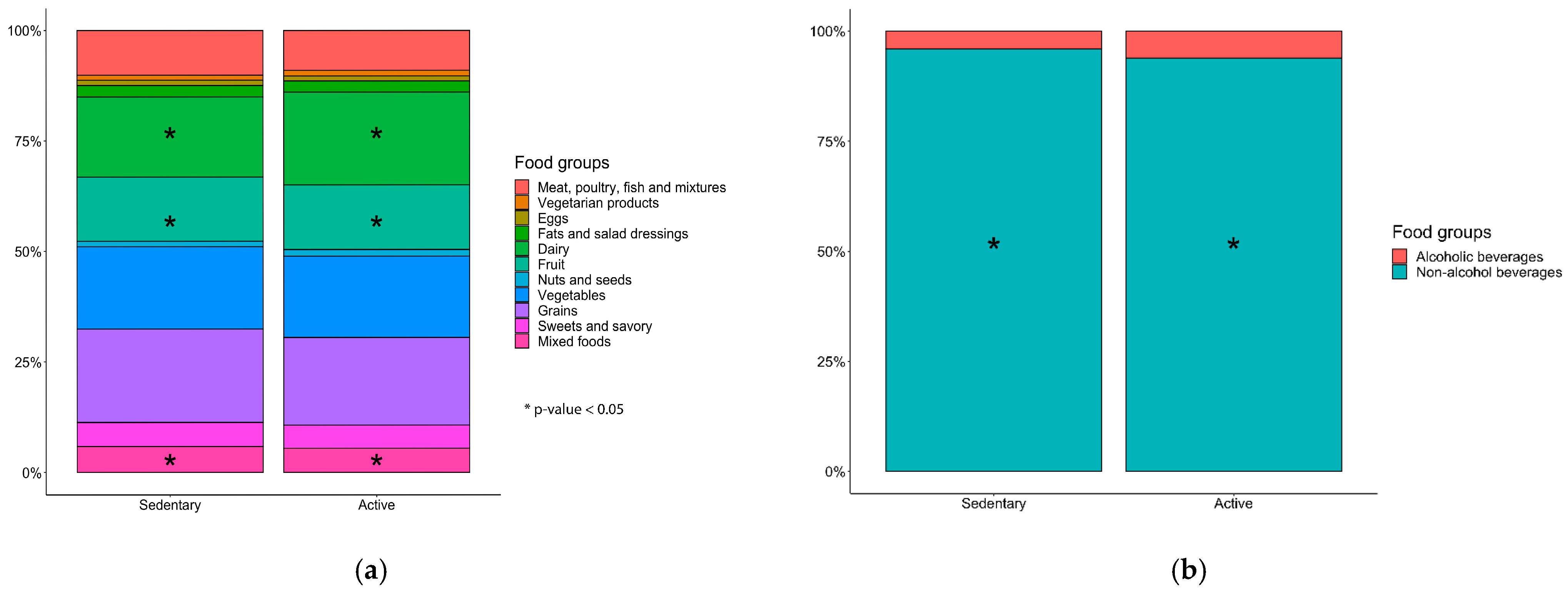
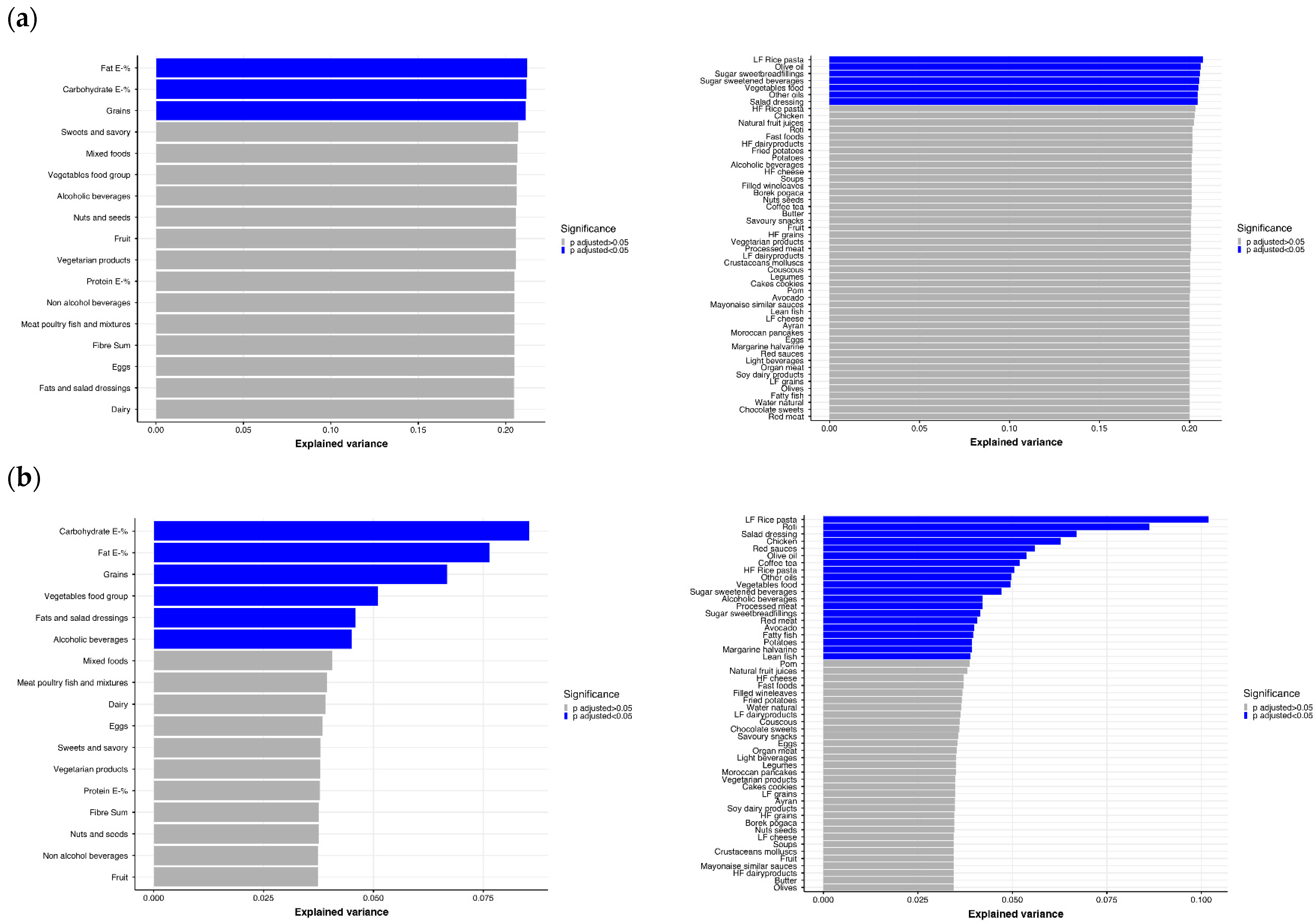
2.7. Diet and Specific Food Groups Characterize the Composition of the Gut Microbiome
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Body Composition, Function, and Biochemistry
4.3. Physical Activity
4.3.1. Subjective Physical Activity Monitor
4.3.2. Objective Physical Activity Monitor
4.4. Dietary Intake and Food Groups
4.5. Fecal Gut Microbiome Composition and Functionality
4.5.1. Profiling of Fecal Microbiota Composition
4.5.2. Processing of 16s rRNA Gene Reads and ASV Generation
4.5.3. Characteristics of Gut Microbiota Composition
4.5.4. Microbial Data Preparation
4.6. Statistical and Bioinformatics Analysis
4.6.1. Statistical Analysis of Clinical Outcomes
4.6.2. Statistical Analysis of Gut Microbiota Composition
4.6.3. Machine Learning
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Owen, N.; Healy, G.N.; Dempsey, P.C.; Salmon, J.; Timperio, A.; Clark, B.K.; Goode, A.D.; Koorts, H.; Ridgers, N.D.; Hadgraft, N.T.; et al. Sedentary Behavior and Public Health: Integrating the Evidence and Identifying Potential Solutions. Annu. Rev. Public Health 2020, 41, 265–287. [Google Scholar] [CrossRef] [Green Version]
- Guthold, R.; Stevens, G.A.; Riley, L.M.; Bull, F.C. Worldwide trends in insufficient physical activity from 2001 to 2016: A pooled analysis of 358 population-based surveys with 1·9 million participants. Lancet Glob. Health 2018, 6, e1077–e1086. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Global Action Plan on Physical Activity 2018–2030: More Active People for a Healthier World; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Young, D.R.; Hivert, M.-F.; Alhassan, S.; Camhi, S.M.; Ferguson, J.F.; Katzmarzyk, P.T.; Lewis, C.E.; Owen, N.; Perry, C.K.; Siddique, J.; et al. Sedentary Behavior and Cardiovascular Morbidity and Mortality: A Science Advisory From the American Heart Association. Circulation 2016, 134, e262–e279. [Google Scholar] [CrossRef]
- Wilmot, E.G.; Edwardson, C.L.; Achana, F.A.; Davies, M.J.; Gorely, T.; Gray, L.J.; Khunti, K.; Yates, T.; Biddle, S.J.H. Sedentary time in adults and the association with diabetes, cardiovascular disease and death: Systematic review and meta-analysis. Diabetologia 2012, 55, 2895–2905. [Google Scholar] [CrossRef] [PubMed]
- Patterson, R.; McNamara, E.; Tainio, M.; de Sá, T.H.; Smith, A.D.; Sharp, S.J.; Edwards, P.; Woodcock, J.; Brage, S.; Wijndaele, K. Sedentary behaviour and risk of all-cause, cardiovascular and cancer mortality, and incident type 2 diabetes: A systematic review and dose response meta-analysis. Eur. J. Epidemiol. 2018, 33, 811–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekelund, U.; Tarp, J.; Steene-Johannessen, J.; Hansen, B.H.; Jefferis, B.; Fagerland, M.W.; Whincup, P.; Diaz, K.M.; Hooker, S.P.; Chernofsky, A.; et al. Dose-response associations between accelerometry measured physical activity and sedentary time and all cause mortality: Systematic review and harmonised meta-analysis. BMJ 2019, 366, l4570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.-M.; Shiroma, E.J.; Lobelo, F.; Puska, P.; Blair, S.N.; Katzmarzyk, P.T.; Lancet Physical Activity Series Working Group. Effect of physical inactivity on major non-communicable diseases worldwide: An analysis of burden of disease and life expectancy. Lancet 2012, 380, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Arnett, D.K.; Blumenthal, R.S.; Albert, M.A.; Buroker, A.B.; Goldberger, Z.D.; Hahn, E.J.; Himmelfarb, C.D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W.; et al. 2019 ACC/AHA Guideline on the Primary Prevention of Cardiovascular Disease: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2019, 140, e596–e646. [Google Scholar] [CrossRef]
- Cosentino, F.; Grant, P.J.; Aboyans, V.; Bailey, C.J.; Ceriello, A.; Delgado, V.; Federici, M.; Filippatos, G.; Grobbee, D.E.; Hansen, T.B.; et al. 2019 ESC Guidelines on diabetes, pre-diabetes, and cardiovascular diseases developed in collaboration with the EASD. Eur. Heart J. 2020, 41, 255–323. [Google Scholar] [CrossRef] [Green Version]
- Yumuk, V.; Tsigos, C.; Fried, M.; Schindler, K.; Busetto, L.; Micic, D.; Toplak, H. European Guidelines for Obesity Management in Adults. Obes. Facts 2015, 8, 402–424. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 852. [Google Scholar] [CrossRef] [Green Version]
- Manor, O.; Dai, C.L.; Kornilov, S.A.; Smith, B.; Price, N.D.; Lovejoy, J.C.; Gibbons, S.M.; Magis, A.T. Health and disease markers correlate with gut microbiome composition across thousands of people. Nat. Commun. 2020, 11, 5206. [Google Scholar] [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Mailing, L.J.; Allen, J.M.; Buford, T.W.; Fields, C.J.; Woods, J.A. Exercise and the Gut Microbiome: A Review of the Evidence, Potential Mechanisms, and Implications for Human Health. Exerc. Sport Sci. Rev. 2019, 47, 75–85. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.J.; Vangay, P.; Al-Ghalith, G.A.; Hillmann, B.M.; Ward, T.L.; Shields-Cutler, R.R.; Kim, A.D.; Shmagel, A.K.; Syed, A.N.; Walter, J.; et al. Daily Sampling Reveals Personalized Diet-Microbiome Associations in Humans. Cell Host Microbe 2019, 25, 789–802.e5. [Google Scholar] [CrossRef] [PubMed]
- Barton, W.; Penney, N.C.; Cronin, O.; Garcia-Perez, I.; Molloy, M.G.; Holmes, E.; Shanahan, F.; Cotter, P.D.; O’Sullivan, O. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut 2018, 67, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Grosicki, G.J.; Durk, R.P.; Bagley, J.R. Rapid gut microbiome changes in a world-class ultramarathon runner. Physiol. Rep. 2019, 7, e14313. [Google Scholar] [CrossRef] [Green Version]
- Munukka, E.; Ahtiainen, J.P.; Puigbó, P.; Jalkanen, S.; Pahkala, K.; Keskitalo, A.; Kujala, U.M.; Pietilä, S.; Hollmén, M.; Elo, L.; et al. Six-week endurance exercise alters gut metagenome that is not reflected in systemic metabolism in over-weight women. Front. Microbiol. 2018, 9, 2323. [Google Scholar] [CrossRef] [PubMed]
- Scheiman, J.; Luber, J.M.; Chavkin, T.A.; MacDonald, T.; Tung, A.; Pham, L.-D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T.; et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef]
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, Y.; Ni, Y.; Cheung, C.K.Y.; Lam, K.S.L.; Wang, Y.; Xia, Z.; Ye, D.; Guo, J.; Tse, M.A.; et al. Gut Microbiome Fermentation Determines the Efficacy of Exercise for Diabetes Prevention. Cell Metab. 2020, 31, 77–91.e5. [Google Scholar] [CrossRef] [PubMed]
- Motiani, K.K.; Collado, M.C.; Eskelinen, J.-J.; Virtanen, K.A.; Löyttyniemi, E.; Salminen, S.; Nuutila, P.; Kalliokoski, K.K.; Hannukainen, J.C. Exercise Training Modulates Gut Microbiota Profile and Improves Endotoxemia. Med. Sci. Sports. Exerc. 2020, 52, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Clauss, M.; Gérard, P.; Mosca, A.; Leclerc, M. Interplay Between Exercise and Gut Microbiome in the Context of Human Health and Performance. Front. Nutr. 2021, 8, 637010. [Google Scholar] [CrossRef] [PubMed]
- Lampinen, E.-K.; Eloranta, A.-M.; Haapala, E.A.; Lindi, V.; Väistö, J.; Lintu, N.; Karjalainen, P.; Kukkonen-Harjula, K.; Laaksonen, D.; Lakka, T.A. Physical activity, sedentary behaviour, and socioeconomic status among Finnish girls and boys aged 6–8 years. Eur. J. Sport Sci. 2017, 17, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Alley, S.J.; Schoeppe, S.; Rebar, A.L.; Hayman, M.; Vandelanotte, C. Age differences in physical activity intentions and implementation intention preferences. J. Behav. Med. 2018, 41, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Loyen, A.; Nicolaou, M.; Snijder, M.B.; Peters, R.J.G.; Stronks, K.; Langøien, L.J.; van der Ploeg, H.P.; Brug, J.; Lakerveld, J. Objectively measured sedentary time among five ethnic groups in Amsterdam: The HELIUS study. PLoS ONE 2017, 12, e0182077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturkenboom, S.M.; Dekker, L.H.; Lamkaddem, M.; Schaap, L.A.; de Vries, J.H.; Stronks, K.; Nicolaou, M. Acculturation and dietary patterns among residents of Surinamese origin in The Netherlands: The HELIUS dietary pattern study. Public Health Nutr. 2016, 19, 682–692. [Google Scholar] [CrossRef] [Green Version]
- Deschasaux, M.; Bouter, K.E.; Prodan, A.; Levin, E.; Groen, A.K.; Herrema, H.; Tremaroli, V.; Bakker, G.J.; Attaye, I.; Pinto-Sietsma, S.-J.; et al. Depicting the composition of gut microbiota in a population with varied ethnic origins but shared geography. Nat. Med. 2018, 24, 1526–1531. [Google Scholar] [CrossRef]
- Vaccaro, J.A.; Huffman, F.G. Sex and Race/Ethnicity Differences in Following Dietary and Exercise Recommendations for U.S. Representative Sample of Adults With Type 2 Diabetes. Am. J. Mens Health 2017, 11, 380–391. [Google Scholar] [CrossRef] [Green Version]
- Snijder, M.B.; Galenkamp, H.; Prins, M.; Derks, E.M.; Peters, R.J.G.; Zwinderman, A.H.; Stronks, K. Cohort profile: The Healthy Life in an Urban Setting (HELIUS) study in Amsterdam, The Netherlands. BMJ Open 2017, 7, e017873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.-Y.; Xia, G.-H.; Lu, J.-Q.; Chen, M.-X.; Zhen, X.; Wang, S.; You, C.; Nie, J.; Zhou, H.-W.; Yin, J. Impaired renal function and dysbiosis of gut microbiota contribute to increased trimethylamine-N-oxide in chronic kidney disease patients. Sci. Rep. 2017, 7, 1445. [Google Scholar] [CrossRef] [PubMed]
- Bressa, C.; Bailén-Andrino, M.; Pérez-Santiago, J.; González-Soltero, R.; Pérez, M.; Montalvo-Lominchar, M.G.; Maté-Muñoz, J.L.; Domínguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar] [CrossRef] [Green Version]
- Whisner, C.M.; Maldonado, J.; Dente, B.; Krajmalnik-Brown, R.; Bruening, M. Diet, physical activity and screen time but not body mass index are associated with the gut microbiome of a diverse cohort of college students living in university housing: A cross-sectional study. BMC Microbiol. 2018, 18, 210. [Google Scholar] [CrossRef]
- Aya, V.; Flórez, A.; Perez, L.; Ramírez, J.D. Association between physical activity and changes in intestinal microbiota composition: A systematic review. PLoS ONE 2021, 16, e0247039. [Google Scholar] [CrossRef]
- Estaki, M.; Pither, J.; Baumeister, P.; Little, J.P.; Gill, S.K.; Ghosh, S.; Ahmadi-Vand, Z.; Marsden, K.R.; Gibson, D.L. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome 2016, 4, 42. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.M.; Mailing, L.J.; Niemiro, G.M.; Moore, R.; Cook, M.D.; White, B.A.; Holscher, H.D.; Woods, J.A. Exercise Alters Gut Microbiota Composition and Function in Lean and Obese Humans. Med. Sci. Sports Exerc. 2018, 50, 747–757. [Google Scholar] [CrossRef]
- Koopen, A.M.; de Clercq, N.C.; Warmbrunn, M.V.; Herrema, H.; Davids, M.; de Groot, P.F.; Kootte, R.S.; Bouter, K.E.C.; Nieuwdorp, M.; Groen, A.K.; et al. Plasma Metabolites Related to Peripheral and Hepatic Insulin Sensitivity Are Not Directly Linked to Gut Microbiota Composition. Nutrients 2020, 12, 2308. [Google Scholar] [CrossRef]
- Nay, K.; Jollet, M.; Goustard, B.; Baati, N.; Vernus, B.; Pontones, M.; Lefeuvre-Orfila, L.; Bendavid, C.; Rué, O.; Mariadassou, M.; et al. Gut bacteria are critical for optimal muscle function: A potential link with glucose homeostasis. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E158–E171. [Google Scholar] [CrossRef]
- van der Kooi, A.-L.L.F.; Snijder, M.B.; Peters, R.J.G.; van Valkengoed, I.G.M. The Association of Handgrip Strength and Type 2 Diabetes Mellitus in Six Ethnic Groups: An Analysis of the HELIUS Study. PLoS ONE 2015, 10, e0137739. [Google Scholar] [CrossRef]
- Studenski, S.; Perera, S.; Patel, K.; Rosano, C.; Faulkner, K.; Inzitari, M.; Brach, J.; Chandler, J.; Cawthon, P.; Connor, E.B.; et al. Gait speed and survival in older adults. JAMA 2011, 305, 50–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, C.-H.; Tien, C.-P.; Li, C.-I.; L’Heureux, A.; Liu, C.-S.; Lin, C.-H.; Lin, C.-C.; Lai, S.-W.; Lai, M.-M.; Lin, W.-Y. Mid-upper arm circumference, calf circumference and mortality in Chinese long-term care facility residents: A prospective cohort study. BMJ Open 2018, 8, e020485. [Google Scholar] [CrossRef] [Green Version]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.B.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef]
- Fielding, R.A.; Reeves, A.R.; Jasuja, R.; Liu, C.; Barrett, B.B.; Lustgarten, M.S. Muscle strength is increased in mice that are colonized with microbiota from high-functioning older adults. Exp. Gerontol. 2019, 127, 110722. [Google Scholar] [CrossRef] [PubMed]
- Zhernakova, A.; Kurilshikov, A.; Bonder, M.J.; Tigchelaar, E.F.; Schirmer, M.; Vatanen, T.; Mujagic, Z.; Vila, A.V.; Falony, G.; Vieira-Silva, S.; et al. Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science 2016, 352, 565–569. [Google Scholar] [CrossRef] [Green Version]
- van Etten, S.; Crielaard, L.; Muilwijk, M.; van Valkengoed, I.; Snijder, M.B.; Stronks, K.; Nicolaou, M. Lifestyle clusters related to type 2 diabetes and diabetes risk in a multi-ethnic population: The HELIUS study. Prev. Med. 2020, 137, 106141. [Google Scholar] [CrossRef]
- Jang, L.-G.; Choi, G.; Kim, S.-W.; Kim, B.-Y.; Lee, S.; Park, H. The combination of sport and sport-specific diet is associated with characteristics of gut microbiota: An observational study. J. Int. Soc. Sports Nutr. 2019, 16, 21. [Google Scholar] [CrossRef] [Green Version]
- Röytiö, H.; Mokkala, K.; Vahlberg, T.; Laitinen, K. Dietary intake of fat and fibre according to reference values relates to higher gut microbiota richness in overweight pregnant women. Br. J. Nutr. 2017, 118, 343–352. [Google Scholar] [CrossRef]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-induced extinctions in the gut microbiota compound over generations. Nature 2016, 529, 212–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Nguyen, L.H.; Song, M.; Wang, D.D.; Franzosa, E.A.; Cao, Y.; Joshi, A.; Drew, D.A.; Mehta, R.; Ivey, K.L.; et al. Dietary fiber intake, the gut microbiome, and chronic systemic inflammation in a cohort of adult men. Genome Med. 2021, 13, 102. [Google Scholar] [CrossRef]
- Deehan, E.C.; Yang, C.; Perez-Muñoz, M.E.; Nguyen, N.K.; Cheng, C.C.; Triador, L.; Zhang, Z.; Bakal, J.A.; Walter, J. Precision Microbiome Modulation with Discrete Dietary Fiber Structures Directs Short-Chain Fatty Acid Production. Cell Host Microbe 2020, 27, 389–404.e6. [Google Scholar] [CrossRef]
- Verhaar, B.J.H.; Collard, D.; Prodan, A.; Levels, J.H.M.; Zwinderman, A.H.; Bäckhed, F.; Vogt, L.; Peters, M.J.L.; Muller, M.; Nieuwdorp, M.; et al. Associations between gut microbiota, faecal short-chain fatty acids, and blood pressure across ethnic groups: The HELIUS study. Eur. Heart J. 2020, 41, 4259–4267. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.P.N.; Mannerås-Holm, L.; Puschmann, R.; Wu, H.; Troise, A.D.; Nijsse, B.; Boeren, S.; Bäckhed, F.; Fiedler, D.; DeVos, W.M. Conversion of dietary inositol into propionate and acetate by commensal Anaerostipes associates with host health. Nat. Commun. 2021, 12, 4798. [Google Scholar] [CrossRef]
- Cicero, N.; Albergamo, A.; Salvo, A.; Bua, G.D.; Bartolomeo, G.; Mangano, V.; Rotondo, A.; Di Stefano, V.; Di Bella, G.; Dugo, G. Chemical characterization of a variety of cold-pressed gourmet oils available on the Brazilian market. Food Res. Int. 2018, 109, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Koistinen, V.M.; Mattila, O.; Katina, K.; Poutanen, K.; Aura, A.-M.; Hanhineva, K. Metabolic profiling of sourdough fermented wheat and rye bread. Sci. Rep. 2018, 8, 5684. [Google Scholar] [CrossRef]
- Bento-Silva, A.; Koistinen, V.M.; Mena, P.; Bronze, M.R.; Hanhineva, K.; Sahlstrøm, S.; Kitrytė, V.; Moco, S.; Aura, A.-M. Factors affecting intake, metabolism and health benefits of phenolic acids: Do we understand individual variability? Eur. J. Nutr. 2019, 59, 1275–1293. [Google Scholar] [CrossRef] [Green Version]
- Mokkala, K.; Houttu, N.; Cansev, T.; Laitinen, K. Interactions of dietary fat with the gut microbiota: Evaluation of mechanisms and metabolic consequences. Clin. Nutr. 2020, 39, 994–1018. [Google Scholar] [CrossRef]
- Abu-Ghazaleh, N.; Chua, W.J.; Gopalan, V. Intestinal microbiota and its association with colon cancer and red/processed meat consumption. J. Gastroenterol. Hepatol. 2021, 36, 75–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bel Lassen, P.; Attaye, I.; Adriouch, S.; Nicolaou, M.; Aron-Wisnewsky, J.; Nielsen, T.; Chakaroun, R.; Le Chatelier, E.; Forslund, S.; Belda, E.; et al. Protein Intake, Metabolic Status and the Gut Microbiota in Different Ethnicities: Results from Two Independent Cohorts. Nutrients 2021, 13, 3159. [Google Scholar] [CrossRef]
- Vujkovic-Cvijin, I.; Sklar, J.; Jiang, L.; Natarajan, L.; Knight, R.; Belkaid, Y. Host variables confound gut microbiota studies of human disease. Nature 2020, 587, 448–454. [Google Scholar] [CrossRef]
- Nicolaou, M.; Gademan, M.G.J.; Snijder, M.B.; Engelbert, R.H.H.; Dijkshoorn, H.; Terwee, C.B.; Stronks, K. Validation of the SQUASH Physical Activity Questionnaire in a Multi-Ethnic Population: The HELIUS Study. PLoS ONE 2016, 11, e0161066. [Google Scholar] [CrossRef]
- Laudadio, I.; Fulci, V.; Palone, F.; Stronati, L.; Cucchiara, S.; Carissimi, C. Quantitative Assessment of Shotgun Metagenomics and 16S rDNA Amplicon Sequencing in the Study of Human Gut Microbiome. OMICS 2018, 22, 248–254. [Google Scholar] [CrossRef]
- Durazzi, F.; Sala, C.; Castellani, G.; Manfreda, G.; Remondini, D.; De Cesare, A. Comparison between 16S rRNA and shotgun sequencing data for the taxonomic characterization of the gut microbiota. Sci. Rep. 2021, 11, 3030. [Google Scholar] [CrossRef]
- Newland, G.A.I.; Gibson, G.R.; Jackson, F.L.; Wijeyesekera, A. Assessment of stool collection and storage conditions for in vitro human gut model studies. J. Microbiol. Methods 2021, 185, 106230. [Google Scholar] [CrossRef] [PubMed]
- Wendel-Vos, S.; van den Berg, M.; Duijvestijn, M.; de Hollander, E. Beweegrichtlijnen en Wekelijks Sporter. RIVM-Briefrapport 2019-0237; Rijksinstituut voor Volksgezondheid en Milieu (RIVM): Bilthoven, The Netherlands, 2019. [Google Scholar]
- Bull, F.C.; Al-Ansari, S.S.; Biddle, S.; Borodulin, K.; Buman, M.P.; Cardon, G.; Carty, C.; Chaput, J.-P.; Chastin, S.; Chou, R.; et al. World Health Organization 2020 guidelines on physical activity and sedentary behaviour. Br. J. Sports Med. 2020, 54, 1451–1462. [Google Scholar] [CrossRef]
- Brage, S.; Brage, N.; Franks, P.W.; Ekelund, U.; Wareham, N.J. Reliability and validity of the combined heart rate and movement sensor Actiheart. Eur. J. Clin. Nutr. 2005, 59, 561–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beukers, M.H.; Dekker, L.H.; de Boer, E.J.; Perenboom, C.W.M.; Meijboom, S.; Nicolaou, M.; de Vries, J.H.M.; Brants, H.A.M. Development of the HELIUS food frequency questionnaires: Ethnic-specific questionnaires to assess the diet of a multiethnic population in The Netherlands. Eur. J. Clin. Nutr. 2015, 69, 579–584. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complete | SQUASH: Subjective Monitoring | ActiHeart: Objective Monitoring | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Complete | Sedentary | Active | p-Value | Complete | Sedentary | Active | p-Value | ||
| n | 1334 | 1309 | 441 | 868 | 162 | 100 | 62 | ||
| Age | 51.0 ± 10.8 | 51.9 ± 10.7 | 49.0 ± 10.5 | 53.4 ± 10.5 | <0.001 | 51.1 ± 7.4 | 51.8 ± 6.5 | 50.0 ± 8.5 | 0.125 |
| Seks (n, %) | |||||||||
| Men | 647 (48.5) | 633 (48.4) | 203 (46.0) | 431 (49.1) | 0.187 | 76 (46.9) | 47 (47.0) | 29 (46.8) | 0.975 |
| Women | 687 (51.5) | 673 (51.4) | 238 (54.0) | 435 (50.9) | 0.187 | 86 (53.1) | 53 (53.0) | 33 (53.2) | 0.975 |
| Ethnicity (n, %) | |||||||||
| AS | 170 (12.7) | 166 (12.7) | 75 (17.0) | 91 (10.5) | 0.001 | 13 (8.0) | 6 (6.0) | 7 (11.3) | 0.250 |
| SAS | 99 (7.4) | 92 (7.0) | 46 (10.4) | 46 (5.3) | 0.001 | 11 (6.8) | 5 (5.0) | 6 (9.7) | 0.228 |
| Maroccan | 346 (25.9) | 344 (26.3) | 110 (24.9) | 234 (27.0) | 0.434 | 43 (26.5) | 28 (28.0) | 15 (24.2) | 0.597 |
| Turkish | 286 (21.4) | 281 (21.5) | 109 (24.7) | 172 (19.8) | 0.041 | 32 (19.8) | 22 (22.0) | 10 (6.2) | 0.362 |
| Dutch origin | 434 (32.5) | 426 (32.5) | 101 (22.9) | 325 (37.4) | <0.001 | 63 (38.9) | 39 (39.0) | 24 (38.7) | 0.971 |
| Weight (kg) | 77.4 ± 15.0 | 77.4 ± 14.9 | 79.4 ± 16.5 | 76.4 ± 14.0 | 0.001 | 76.7 ± 14.9 | 76.8 ± 15.0 | 76.7 ± 14.9 | 0.971 |
| BMI (kg/m2) | 27.1 ± 4.8 | 27.3 ± 4.8 | 28.0 ± 5.2 | 26.7 ± 4.5 | <0.001 | 26.5 ± 4.6 | 26.6 ± 4.7 | 26.2 ± 4.4 | 0.577 |
| Fat mass (%) | 30.6 ± 9.2 | 30.6 ± 9.3 | 31.7 ± 9.5 | 30.0 ± 9.1 | 0.002 | 30.9 ± 7.8 | 31.4 ± 7.9 | 30.1 ± 7.6 | 0.314 |
| WC (cm) | 94.0 ± 12.4 | 94.0 ± 12.4 | 95.3 ± 13.6 | 93.3 ± 11.7 | 0.007 | 93.1 ± 11.5 | 93.9 ± 10.8 | 91.7 ± 12.5 | 0.235 |
| WHR | 0.92 ± 0.1 | 1.00 ± 2.1 | 0.91 ± 0.1 | 0.98 ± 1.7 | 0.292 | 0.92 ± 0.1 | 0.93 ± 0.1 | 0.91 ± 0.1 | 0.034 |
| TC (cm) | 58.8 ± 10.3 | 58.9 ± 10.4 | 59.8 ± 10.2 | 58.4 ± 10.4 | 0.022 | 58.2 ± 9.8 | 58.0 ± 8.4 | 58.6 ± 11.8 | 0.733 |
| CC (cm) | 37.5 ± 3.4 | 37.5 ± 3.4 | 37.9 ± 3.9 | 37.3 ± 3.2 | 0.001 | 37.6 ± 3.4 | 37.9 ± 3.6 | 37.2 ± 3.1 | 0.233 |
| Muscle strength (N) | 208.6 ± 76.9 | 208.5 ± 76.9 | 201.0 ± 77.8 | 212.3 ± 76.2 | 0.013 | 209.5 ± 69.1 | 208.0 ± 70.0 | 211.7 ± 68.3 | 0.745 |
| Creatinine (µmol/L) | 74 (74.6–76.5) | 74 (74.7–76.6) | 73 (72.7–76.5) | 75 (75.1–77.2) | 0.007 * | 74.0 (72.6–77.4) | 76.8 (71.1–76.7) | 72.5 (72.4–81.2) | 0.642 * |
| CK (µmol/L) | 119 (146.7–161.9) | 120 (147.2–162.7) | 112 (141.1–175.7) | 123 (145.5–160.9) | 0.141 * | 123.5 (132.8–167.0) | 117.0 (122.0–159.6) | 134.0 (131.2–197.9) | 0.305 * |
| Overall (n = 1309) | Sedentary (n = 441) | Active (n = 868) | p-Value | |
|---|---|---|---|---|
| Energy (kcal/d) | 2280.5 ± 975.3 | 2211.9 ± 926.0 | 2317.2 ± 996.2 | 0.056 |
| Carbohydrates (E-%) | 45.0 ± 9.4 | 45.2 ± 7.8 | 44.7 ± 8.3 | 0.414 |
| Protein (E-%) | 16.4 ± 4.7 | 16.5 ± 3.1 | 16.2 ± 3.3 | 0.917 |
| Fat (E-%) | 32.0 ± 8.5 | 31.3 ± 6.3 | 31.7 ± 6.5 | 0.912 |
| SFA (E-%) | 11.0 ± 6.4 | 10.8 ± 2.9 | 10.8 ± 3.2 | 0.503 |
| MUFA (E-%) | 12.6 ± 4.3 | 12.6 ± 3.2 | 12.5 ± 3.3 | 0.925 |
| PUFA (E-%) | 7.8 ± 4.0 | 7.4 ± 2.3 | 7.7 ± 2.3 | 0.343 |
| Alcohol (g/d) | 7.1 ± 13.0 | 5.0 ± 11.1 | 8.1 ± 13.8 | <0.001 |
| Fiber (g/d) | 24.7 ± 10.7 | 23.5 ± 9.6 | 25.1 ± 10.7 | 0.006 |
| Muscle Strength | CC | TC | Creatinine | CK | |||
|---|---|---|---|---|---|---|---|
| Richness | Model 1 | R2 | 0.251 | 0.171 | 0.166 | 0.171 | 0.169 |
| R2 adj. | 0.244 | 0.166 | 0.160 | 0.166 | 0.164 | ||
| β | 0.121 | 5.322 | 0.282 | 0.679 | 0.046 | ||
| p-value | 0.119 | 0.005 | 0.525 | 0.016 | 0.128 | ||
| Model 2 | R2 | 0.185 | 0.186 | 0.181 | 0.185 | 0.183 | |
| R2 adj. | 0.178 | 0.179 | 0.174 | 0.178 | 0.176 | ||
| β | 0.154 | 5.106 | 0.216 | 0.682 | 0.040 | ||
| p-value | 0.038 | 0.005 | 0.609 | 0.012 | 0.174 | ||
| Shannon index | Model 1 | R2 | 0.207 | 0.209 | 0.207 | 0.213 | 0.205 |
| R2 adj. | 0.202 | 0.204 | 0.202 | 0.208 | 0.200 | ||
| β | 0.000 | 0.016 | 0.003 | 0.003 | 0.000 | ||
| p-value | 0.087 | 0.010 | 0.044 | 0.000 | 0.504 | ||
| Model 2 | R2 | 0.227 | 0.227 | 0.225 | 0.002 | 0.224 | |
| R2 adj. | 0.220 | 0.220 | 0.218 | 0.001 | 0.217 | ||
| β | 0.000 | 0.013 | 0.002 | 0.003 | 0.000 | ||
| p-value | 0.079 | 0.021 | 0.071 | 0.000 | 0.512 | ||
| Faith’s phylogenetic diversity | Model 1 | R2 | 0.237 | 0.241 | 0.237 | 0.243 | 0.236 |
| R2 adj. | 0.232 | 0.236 | 0.232 | 0.239 | 0.231 | ||
| β | 0.004 | 0.249 | 0.030 | 0.046 | 0.000 | ||
| p-value | 0.235 | 0.004 | 0.141 | 0.000 | 0.727 | ||
| Model 2 | R2 | 0.253 | 0.255 | 0.251 | 0.257 | 0.251 | |
| R2 adj. | 0.246 | 0.248 | 0.244 | 0.250 | 0.244 | ||
| β | 0.005 | 0.236 | 0.025 | 0.044 | 0.001 | ||
| p-value | 0.139 | 0.004 | 0.197 | 0.000 | 0.707 | ||
| Simpson index | Model 1 | R2 | 0.098 | 0.098 | 0.098 | 0.102 | 0.097 |
| R2 adj. | 0.093 | 0.092 | 0.093 | 0.097 | 0.091 | ||
| β | 0.000 | 0.001 | 0.000 | 0.000 | 0.000 | ||
| p-value | 0.660 | 0.244 | 0.085 | 0.006 | 0.832 | ||
| Model 2 | R2 | 0.104 | 0.103 | 0.103 | 0.106 | 0.102 | |
| R2 adj. | 0.095 | 0.095 | 0.095 | 0.098 | 0.094 | ||
| β | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | ||
| p-value | 0.719 | 0.322 | 0.119 | 0.018 | 0.835 | ||
| Inverse Simpson index | Model 1 | R2 | 0.180 | 0.180 | 0.179 | 0.184 | 0.177 |
| R2 adj. | 0.175 | 0.175 | 0.174 | 0.179 | 0.172 | ||
| β | 0.017 | 0.457 | 0.107 | 0.106 | 0.002 | ||
| p-value | 0.039 | 0.026 | 0.026 | 0.000 | 0.517 | ||
| Model 2 | R2 | 0.188 | 0.187 | 0.186 | 0.191 | 0.185 | |
| R2 adj. | 0.181 | 0.179 | 0.179 | 0.184 | 0.177 | ||
| β | 0.017 | 0.364 | 0.093 | 0.101 | 0.002 | ||
| p-value | 0.030 | 0.062 | 0.040 | 0.001 | 0.561 |
| PCR Cycle Step | Temperature (°C) | Time (min:sec) |
|---|---|---|
| Initial denaturation | 94 | 47:00 |
| Denaturation | 94 | 00:47 |
| Annealing | 52 | 00:60 |
| Elongation | 72 | 01:30 |
| Final elongation | 72 | 10:00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Houttu, V.; Boulund, U.; Nicolaou, M.; Holleboom, A.G.; Grefhorst, A.; Galenkamp, H.; van den Born, B.-J.; Zwinderman, K.; Nieuwdorp, M. Physical Activity and Dietary Composition Relate to Differences in Gut Microbial Patterns in a Multi-Ethnic Cohort—The HELIUS Study. Metabolites 2021, 11, 858. https://doi.org/10.3390/metabo11120858
Houttu V, Boulund U, Nicolaou M, Holleboom AG, Grefhorst A, Galenkamp H, van den Born B-J, Zwinderman K, Nieuwdorp M. Physical Activity and Dietary Composition Relate to Differences in Gut Microbial Patterns in a Multi-Ethnic Cohort—The HELIUS Study. Metabolites. 2021; 11(12):858. https://doi.org/10.3390/metabo11120858
Chicago/Turabian StyleHouttu, Veera, Ulrika Boulund, Mary Nicolaou, Adriaan Georgius Holleboom, Aldo Grefhorst, Henrike Galenkamp, Bert-Jan van den Born, Koos Zwinderman, and Max Nieuwdorp. 2021. "Physical Activity and Dietary Composition Relate to Differences in Gut Microbial Patterns in a Multi-Ethnic Cohort—The HELIUS Study" Metabolites 11, no. 12: 858. https://doi.org/10.3390/metabo11120858