The Effect of Bee Venom Peptides Melittin, Tertiapin, and Apamin on the Human Erythrocytes Ghosts: A Preliminary Study

 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Results
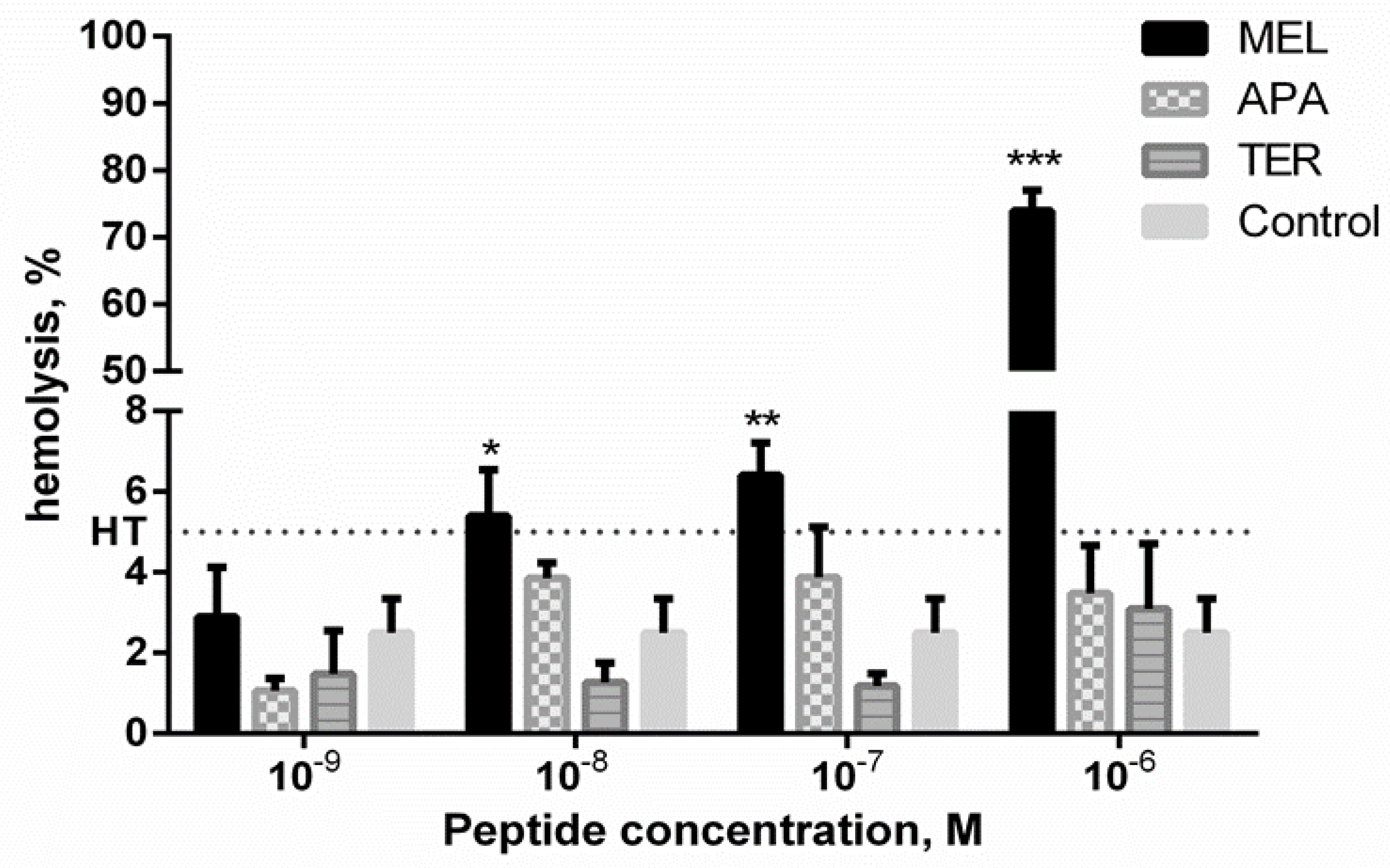
2.1. Hemolytic Activity of Apitoxins
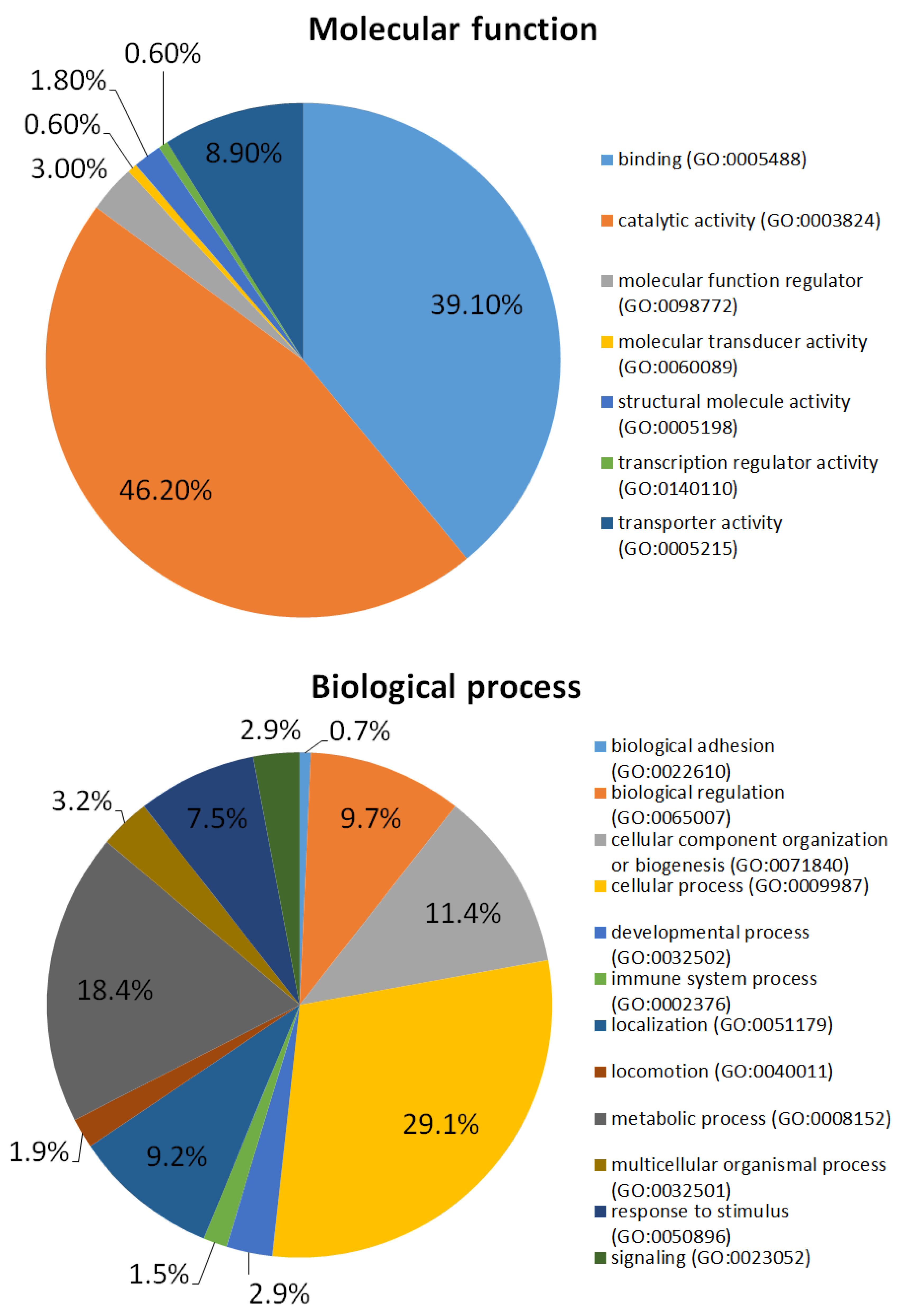
2.2. Red Blood Cells (RBCs) Ghosts Proteome
2.3. The Influence of Different Concentrations of Bee Venom Peptides on the Human Erythrocyte Membrane Proteome
2.4. The Influence of Specific Bee Venom Peptides on the Erythrocyte Membrane Proteome
3. Discussion
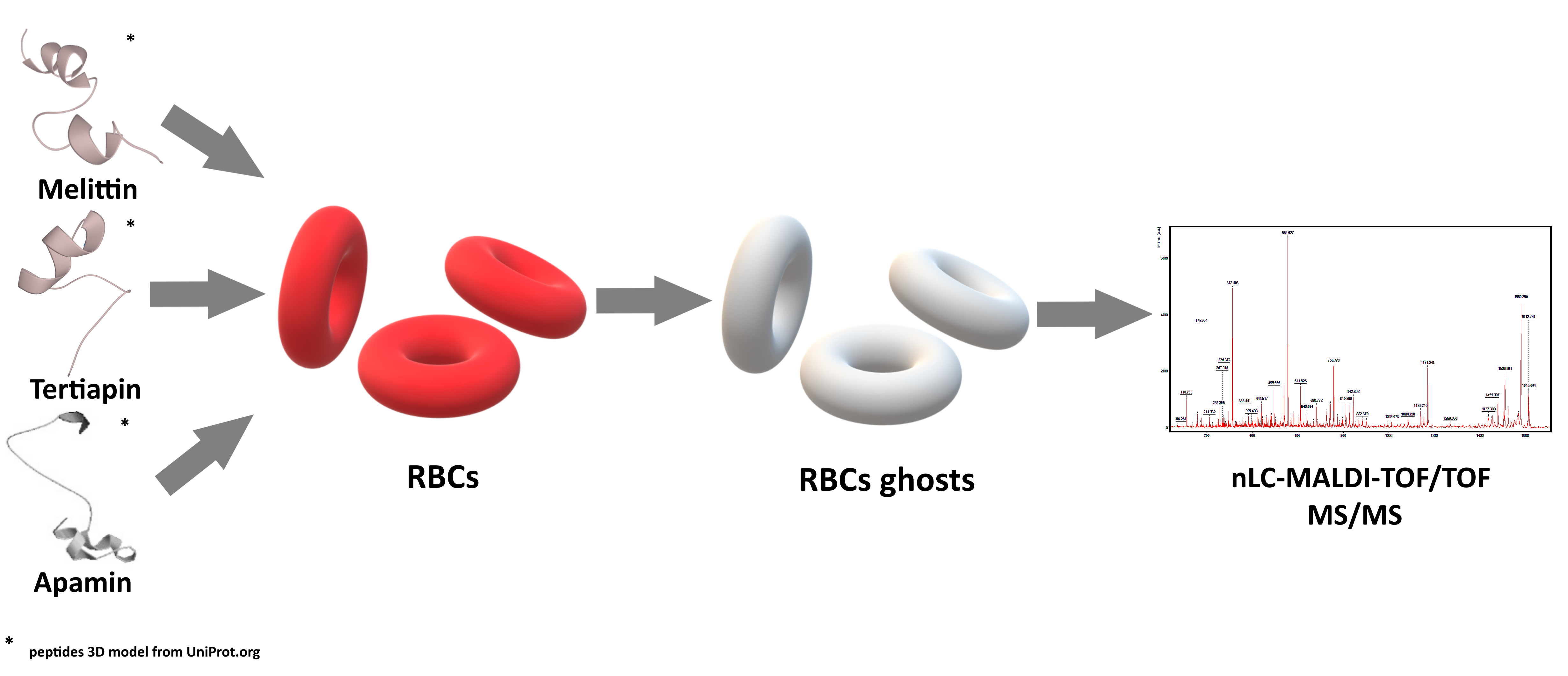
4. Materials and Methods
4.1. Reagents
4.2. Samples Preparation
4.2.1. Human Erythrocyte
4.2.2. Hemolysis Assay
4.2.3. Erythrocytes Shape Evaluation
4.2.4. Erythrocytes Ghosts
4.3. nLC-MALDI-TOF/TOF MS/MS Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bobrowska-Hägerstrand, M.; Wróbel, A.; Mrówczyńska, L.; Söderström, T.; Hägerstrand, H. Modulation of MRP1-like efflux activity in human erythrocytes caused by membrane perturbing agents. Mol. Membr. Biol. 2003, 20, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Pasini, E.M.; Mann, M.; Thomas, A.W. Red blood cell proteomics. Transfus. Clin. Biol. 2010, 17, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Fye, H.K.S.; Mrosso, P.; Bruce, L.; Thézénas, M.L.; Davis, S.; Fischer, R.; Rwegasira, G.L.; Makani, J.; Kessler, B.M. A robust mass spectrometry method for rapid profiling of erythrocyte ghost membrane proteomes. Clin. Proteomics 2018, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- Al-Thani, A.M.; Voss, S.C.; Al-Menhali, A.S.; Barcaru, A.; Horvatovich, P.; Al Jaber, H.; Nikolovski, Z.; Latiff, A.; Georgakopoulos, C.; Merenkov, Z.; et al. Whole blood storage in CPDA1 blood bags alters erythrocyte membrane proteome. Oxid. Med. Cell Longev. 2018, 2018, 6375379. [Google Scholar] [CrossRef]
- Barcellini, W.; Bianchi, P.; Fermo, E.; Imperiali, F.G.; Marcello, A.P.; Vercellati, C.; Zaninoni, A.; Zanella, A. Hereditary red cell membrane defects: Diagnostic and clinical aspects. Blood Transfus. 2011, 9, 274–277. [Google Scholar]
- Mohandas, N.; Gallagher, P.G. Red cell membrane: Past, present, and future. Blood 2008, 112, 3939–3948. [Google Scholar] [CrossRef] [Green Version]
- Basa, B.; Wendimagegnehu, B.; Tilahun, A.; Teshale, A. Review on medicinal value of honeybee products: Apitherapy. Adv. Biol. Res. 2016, 10, 236–247. [Google Scholar]
- Woźniak, M.; Mrówczyńska, L.; Waśkiewicz, A.; Rogoziński, T.; Ratajczak, I. The role of seasonality on the chemical composition, antioxidant activity and cytotoxicity of Polish propolis in human erythrocytes. Brazilian J. Pharmacogn. 2019, 29, 301–308. [Google Scholar] [CrossRef]
- Woźniak, M.; Mrówczyńska, L.; Waśkiewicz, A.; Rogoziński, T.; Ratajczak, I. Phenolic profile and antioxidant activity of propolis extracts from Poland. Nat. Prod. Commun. 2019, 14, 1–7. [Google Scholar] [CrossRef]
- Lubawy, J.; Urbański, A.; Mrówczyńska, L.; Matuszewska, E.; Światły-Błaszkiewicz, A.; Matysiak, J.; Rosinski, G. The influence of bee venom melittin on the functioning of the immune system and the contractile activity of the insect heart—A preliminary study. Toxins 2019, 11, 494. [Google Scholar] [CrossRef] [Green Version]
- Raghuraman, H.; Chattopadhyay, A. Melittin: A membrane-active peptide with diverse functions. Biosci. Rep. 2007, 27, 189–223. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Nomura, F.; Yokoyama, Y.; Tanaka-Takiguchi, Y.; Homma, M.; Takiguchi, K. Multiple membrane interactions and versatile vesicle deformations elicited by melittin. Toxins 2013, 5, 637–664. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.; Bae, H. Anti-inflammatory applications of melittin, a major component of bee venom: Detailed mechanism of action and adverse effects. Molecules 2016, 21, 616. [Google Scholar] [CrossRef]
- Cornara1, L.; Biagi, M.; Xiao, J.; Burlando, B. Therapeutic properties of bioactive compounds from different honeybee products. Front. Pharmacol. 2017, 8, 412. [Google Scholar] [CrossRef]
- Drici, M.D.; Diochot, S.; Terrenoire, C.; Romey, G.; Lazdunski, M. The bee venom peptide tertiapin underlines the role of I(KACh) in acetylcholine-induced atrioventricular blocks. Br. J. Pharmacol. 2000, 131, 569–577. [Google Scholar] [CrossRef] [Green Version]
- Habermann, E.; Fischer, K. Bee venom neurotoxin (apamin): Iodine labeling and characterization of binding sites. Eur. J. Biochem. 1979, 94, 355–364. [Google Scholar] [CrossRef]
- Stackman, R.W.; Hammond, R.S.; Linardatos, E.; Gerlach, A.; Maylie, J.; Adelman, J.P.; Tzounopoulos, T. Small conductance Ca2+-activated K+ channels modulate synaptic plasticity and memory encoding. J. Neurosci. 2002, 22, 10163–10171. [Google Scholar] [CrossRef] [Green Version]
- Deschaux, O.; Bizot, J.C. Apamin produces selective improvements of learning in rats. Neurosci. Lett. 2005, 386, 5–8. [Google Scholar] [CrossRef]
- Kallarackal, A.J.; Simard, J.M.; Bailey, A.M. The effect of apamin, a small conductance calcium activated potassium (SK) channel blocker, on a mouse model of neurofibromatosis 1. Behav. Brain Res. 2013, 237, 71–75. [Google Scholar] [CrossRef]
- Alvarez-Fischer, D.; Noelker, C.; Vulinović, F.; Grünewald, A.; Chevarin, C.; Klein, C.; Oertel, W.H.; Hirsch, E.C.; Michel, P.P.; Hartmann, A. Bee venom and its component apamin as neuroprotective agents in a Parkinson disease mouse model. PLoS ONE 2013, 8, e61700. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.I.; Yang, E.J.; Lee, M.S.; Kim, Y.S.; Huh, Y.; Cho, I.H.; Kang, S.; Koh, H.K. Bee venom reduces neuroinflammation in the MPTP-induced model of Parkinson’s disease. Int. J. Neurosci. 2011, 121, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.C.; Pitt, B. Compositions and Methods for Preserving Red Blood Cells. U.S. Patent No WO2008089337 A1, 24 July 2008. [Google Scholar]
- Pesciotta, E.N.; Sriswasdi, S.; Tang, H.Y.; Mason, P.J.; Bessler, M.; Speicher, D.W. A label-free proteome analysis strategy for identifying quantitative changes in erythrocyte membranes induced by red cell disorders. J. Proteomics 2012, 76, 194–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryk, A.H.; Wiśniewski, J.R. Quantitative analysis of human red blood cell proteome. J. Proteome Res. 2017, 16, 2752–2761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lux, S.E. Anatomy of the red cell membrane skeleton: Unanswered questions. Blood 2016, 127, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Mankelow, T.J.; Satchwell, T.J.; Burton, N.M. Refined views of multi-protein complexes in the erythrocyte membrane. Blood Cells Mol. Dis. 2012, 49, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pasini, E.M.; Kirkegaard, M.; Mortensen, P.; Lutz, H.U.; Thomas, A.W.; Mann, M. In-depth analysis of the membrane and cytosolic proteome of red blood cells. Blood 2006, 108, 791–801. [Google Scholar] [CrossRef]
- Hur, J.; Kim, K.; Lee, S.; Park, H.; Park, Y. Melittin-induced alterations in morphology and deformability of human red blood cells using quantitative phase imaging techniques. Sci. Rep. 2017, 7, 9306. [Google Scholar] [CrossRef] [Green Version]
- Gajski, G.; Garaj-Vrhovac, V. Melittin: A lytic peptide with anticancer properties. Environ. Toxicol. Pharmacol. 2013, 36, 697–705. [Google Scholar] [CrossRef]
- Woods, N.; Niwasabutra, K.; Acevedo, R.; Igoli, J.; Altwaijry, N.A.; Tusiimire, J.; Gray, A.I.; Watson, D.G.; Ferro, V.A. Natural vaccine adjuvants and immunopotentiators derived from plants, fungi, marine organisms, and insects. In Immunopotentiators in Modern Vaccines, 2nd ed.; Academic Press: London, UK, 2016; pp. 211–230. [Google Scholar]
- Kitamura, H.; Yamada, M.; Yokoyama, M.; Akita, H.; Matsushita, K.; Kurachi, Y. Tertiapin potently and selectively blocks muscarinic K+ channels in rabbit cardiac myocytes. J. Pharmacol. Exp. Ther. 2000, 293, 196–205. [Google Scholar]
- Palma, M.S. Insect venom peptides. In Handbook of Biologically Active Peptides; Academic Press: Cambridge, MA, USA, 2006; pp. 389–396. [Google Scholar]
- Miroshnikov, A.I.; Boĭkov, V.A.; Snezhkova, L.G.; Severin, S.E.; Shvets, V.I. Interaction of tertiapin, a neurotoxin from bee venom, with calmodulin. Bioorganicheskaia khimiia 1983, 9, 26–32. [Google Scholar] [PubMed]
- Nelson, G.A.; Andrews, M.L.; Karnovsky, M.J. Control of erythrocyte shape by calmodulin. J. Cell Biol. 1983, 96, 730–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehdi, S.; Angelastro, M.R.; Wiseman, J.S.; Bey, P. Inhibition of the proteolysis of rat erythrocyte membrane proteins by a synthetic inhibitor of calpain. Biochem. Biophys. Res. Commun. 1988, 157, 1117–1123. [Google Scholar] [CrossRef]
- Lou, X.; De Waal, B.F.M.; Milroy, L.G.; Van Dongen, J.L.J. A sample preparation method for recovering suppressed analyte ions in MALDI TOF MS. J. Mass Spectrom. 2015, 50, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Albrethsen, J. Reproducibility in Protein Profiling by MALDI-TOF Mass Spectrometry. Clin. Chem. 2007, 53, 852–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proulx, É.; Power, S.K.; Oliver, D.K.; Sargin, D.; McLaurin, J.; Lambe, E.K. Apamin improves prefrontal nicotinic impairment in mouse model of Alzheimer’s disease. Cereb. Cortex 2019, 30, 563–574. [Google Scholar] [CrossRef]
- Kshatri, A.S.; Gonzalez-Hernandez, A.; Giraldez, T. Physiological roles and therapeutic potential of Ca2+ activated potassium channels in the nervous system. Front. Mol. Neurosci. 2018, 11, 258. [Google Scholar] [CrossRef]
- Huang, Y.; Qiao, F.; Abagyan, R.; Hazard, S.; Tomlinson, S. Defining the CD59-C9 binding interaction. J. Biol. Chem. 2006, 281, 27398–27404. [Google Scholar] [CrossRef] [Green Version]
- Freund, A.; Zhong, F.L.; Venteicher, A.S.; Meng, Z.; Veenstra, T.D.; Frydman, J.; Artandi, S.E. Proteostatic control of telomerase function through TRiC-mediated folding of TCAB1. Cell 2014, 159, 1389–1403. [Google Scholar] [CrossRef] [Green Version]
- Holguin, M.H.; Fredrick, L.R.; Bernshaw, N.J.; Wilcox, L.A.; Parker, C.J. Isolation and characterization of a membrane protein from normal human erythrocytes that inhibits reactive lysis of the erythrocytes of paroxysmal nocturnal hemoglobinuria. J. Clin. Investig. 1989, 84, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Lublin, D.M.; Atkinson, J.P. Decay-accelerating factor: Biochemistry, molecular biology, and function. Ann. Rev. Immunol. 1989, 7, 35–58. [Google Scholar] [CrossRef] [PubMed]
- Slomiany, M.G.; Grass, G.D.; Robertson, A.D.; Yang, X.Y.; Maria, B.L.; Beeson, C.; Toole, B.P. Hyaluronan, CD44, and emmprin regulate lactate efflux and membrane localization of monocarboxylate transporters in human breast carcinoma cells. Cancer Res. 2009, 69, 1293–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirk, P.; Wilson, M.C.; Heddle, C.; Brown, M.H.; Barclay, A.N.; Halestrap, A.P. CD147 is tightly associated with lactate transporters MCT1 and MCT4 and facilitates their cell surface expression. EMBO J. 2000, 19, 3896–3904. [Google Scholar] [CrossRef]
- Muramatsu, T. Basigin (CD147), a multifunctional transmembrane glycoprotein with various binding partners. J. Biochem. 2016, 159, 481–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasiewicz, B.; Sierakowska, A.; Jankowski, W.; Hoffmann, M.; Piorońska, W.; Górnicka, A.; Bielawska, A.; Bielawski, K.; Mrówczyńska, L. Antioxidant and cytotoxic activity of new di- and polyamine caffeine analogues. Free Radic. Res. 2018, 52, 724–736. [Google Scholar] [CrossRef]
- Bessis, M.; Weed, R.I.; Leblond, P.F. Red cell shape. In Physiology, Pathology, Utrastructure; Springer: New York, NY, USA; Berlin/Heidelberg, Germany, 1973; 180p. [Google Scholar]
- Dodge, J.T.; Mitchell, C.; Hanahan, D.J. The preparation and chemical characteristics of hemoglobin-free ghosts of human erythrocytes. Arch. Biochem. Biophys. 1963, 100, 119–130. [Google Scholar] [CrossRef]
- Swiatly, A.; Horala, A.; Matysiak, J.; Hajduk, J.; Nowak-Markwitz, E.; Kokot, Z. Understanding Ovarian Cancer: iTRAQ-Based Proteomics for Biomarker Discovery. Int. J. Mol. Sci. 2018, 19, 2240. [Google Scholar] [CrossRef] [Green Version]
- Matysiak, J.; Hajduk, J.; Mayer, F.; Hebeler, R.; Kokot, Z. Hyphenated LC-MALDI-ToF/ToF and LC-ESI-QToF approach in proteomic characterization of honeybee venom. J. Pharm. Biomed. Anal. 2016, 121, 69–76. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Proteins not Detected in the RBCs Ghosts Samples Treated with Apamin | Proteins Identified only in Rbcs Ghosts Samples Treated with Apamin | ||||
|---|---|---|---|---|---|
| No. | Accession | Protein | No. | Accession | Protein |
| 1. | SBP1_HUMAN | Selenium-binding protein 1 | 1. | BASI_HUMAN | Basigin |
| 2. | TGM2_HUMAN | Protein-glutamine gamma-glutamyltransferase 2 | 2. | S29A1_HUMAN | Equilibrative nucleoside transporter 1 |
| 3. | ESTD_HUMAN | S-formylglutathione hydrolase | 3. | CD44_HUMAN | CD44 antigen |
| 4. | F10A1_HUMAN | Hsc70-interacting protein | 4. | KAP0_HUMAN | cAMP-dependent protein kinase type I-alpha regulatory subunit |
| 5. | LDHB_HUMAN | L-lactate dehydrogenase B chain | 5. | ABCB6_HUMAN | ATP-binding cassette sub-family B member 6, mitochondrial |
| 6. | BLMH_HUMAN | Bleomycin hydrolase | 6. | CD99_HUMAN | CD99 antigen |
| 7. | NSF1C_HUMAN | NSFL1 cofactor p47 | 7. | BCAM_HUMAN | Basal cell adhesion molecule |
| 8. | S10A8_HUMAN | Protein S100-A8 | 8. | CD59_HUMAN | CD59 glycoprotein |
| 9. | RADI_HUMAN | Radixin | |||
| 10. | NDKA_HUMAN | Nucleoside diphosphate kinase A | |||
| 11. | PA1B3_HUMAN | Platelet-activating factor acetylhydrolase IB subunit gamma | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Światły-Błaszkiewicz, A.; Mrówczyńska, L.; Matuszewska, E.; Lubawy, J.; Urbański, A.; Kokot, Z.J.; Rosiński, G.; Matysiak, J. The Effect of Bee Venom Peptides Melittin, Tertiapin, and Apamin on the Human Erythrocytes Ghosts: A Preliminary Study. Metabolites 2020, 10, 191. https://doi.org/10.3390/metabo10050191
Światły-Błaszkiewicz A, Mrówczyńska L, Matuszewska E, Lubawy J, Urbański A, Kokot ZJ, Rosiński G, Matysiak J. The Effect of Bee Venom Peptides Melittin, Tertiapin, and Apamin on the Human Erythrocytes Ghosts: A Preliminary Study. Metabolites. 2020; 10(5):191. https://doi.org/10.3390/metabo10050191
Chicago/Turabian StyleŚwiatły-Błaszkiewicz, Agata, Lucyna Mrówczyńska, Eliza Matuszewska, Jan Lubawy, Arkadiusz Urbański, Zenon J. Kokot, Grzegorz Rosiński, and Jan Matysiak. 2020. "The Effect of Bee Venom Peptides Melittin, Tertiapin, and Apamin on the Human Erythrocytes Ghosts: A Preliminary Study" Metabolites 10, no. 5: 191. https://doi.org/10.3390/metabo10050191