
Expression Patterns of GATA3 in Classical Hodgkin Lymphoma: A Clinico-Pathological Study

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.3. Statistical Analysis
3. Results
3.1. GATA3 Expression Patterns
3.2. Correlation with Clinical and Laboratory Data
3.3. Survival Analysis
3.4. EBV Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cirillo, M.; Reinke, S.; Klapper, W.; Borchmann, S. The translational science of hodgkin lymphoma. Br. J. Haematol. 2019, 184, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Piris, M.A.; Medeiros, L.J.; Chang, K.C. Hodgkin lymphoma: A review of pathological features and recent advances in pathogenesis. Pathology 2020, 52, 154–165. [Google Scholar] [CrossRef]
- Weniger, M.A.; Küppers, R. Molecular biology of Hodgkin lymphoma. Leukemia 2021, 35, 968–981. [Google Scholar] [CrossRef] [PubMed]
- Alaggio, R.; Amador, C.; Anagnostopoulos, I.; Attygalle, A.D.; Araujo, I.B.O.; Berti, E.; Bhagat, G.; Borges, A.M.; Boyer, D.; Calaminici, M.; et al. The 5th edition of the World Health Organization Classification of Haematolymphoid Tumours: Lymphoid Neoplasms. Leukemia 2022, 36, 1720–1748. [Google Scholar] [CrossRef]
- Campo, E.; Jaffe, E.S.; Cook, J.R.; Quintanilla-Martinez, L.; Swerdlow, S.H.; Anderson, K.C.; Brousset, P.; Cerroni, L.; de Leval, L.; Dirnhofer, S.; et al. The International Consensus Classification of Mature Lymphoid Neoplasms: A report from the Clinical Advisory Committee. Blood 2022, 140, 1229–1253. [Google Scholar] [CrossRef] [PubMed]
- Georgoulis, V.; Papoudou-Bai, A.; Makis, A.; Kanavaros, P.; Hatzimichael, E. Unraveling the Immune Microenvironment in Classic Hodgkin Lymphoma: Prognostic and Therapeutic Implications. Biology 2023, 12, 862. [Google Scholar] [CrossRef]
- Jiwa, N.M.; Kanavaros, P.; De Bruin, P.C.; van der Valk, P.; Horstman, A.; Vos, W.; Mullink, H.; Walboomers, J.M.; Meijer, C.J. Presence of Epstein-Barr virus harbouring small and intermediate-sized cells in Hodgkin’s disease. Is there a relationship with Reed-Sternberg cells? J. Pathol. 1993, 170, 129–136. [Google Scholar] [CrossRef]
- Herrera, A.F.L.M.L.; Castellino, S.M.; Li, H.; Rutherford, S.C.; Evens, A.M.; Davison, K.; Punnett, A.; Hodgson, D.C.; Parsons, S.K.; Ahmed, S.; et al. SWOG S1826, a randomized study of nivolumab(N)-AVD versus brentuximab vedotin(BV)-AVD in advanced stage (AS) classic Hodgkin lymphoma (HL). J. Clin. Oncol. 2023, 41, LBA4. [Google Scholar] [CrossRef]
- Khazaeli Najafabadi, M.; Mirzaeian, E.; Memar Montazerin, S.; Tavangar, A.R.; Tabary, M.; Tavangar, S.M. Role of GATA3 in tumor diagnosis: A review. Pathol. Res. Pract. 2021, 226, 153611. [Google Scholar] [CrossRef]
- Geng, X.; Wang, C.; Gao, X.; Chowdhury, P.; Weiss, J.; Villegas, J.A.; Saed, B.; Perera, T.; Hu, Y.; Reneau, J.; et al. GATA-3 is a proto-oncogene in T-cell lymphoproliferative neoplasms. Blood Cancer J. 2022, 12, 149. [Google Scholar] [CrossRef] [PubMed]
- Küppers, R.; Klein, U.; Schwering, I.; Distler, V.; Bräuninger, A.; Cattoretti, G.; Tu, Y.; Stolovitzky, G.A.; Califano, A.; Hansmann, M.L.; et al. Identification of Hodgkin and Reed-Sternberg cell-specific genes by gene expression profiling. J. Clin. Investig. 2003, 111, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Stanelle, J.; Döring, C.; Hansmann, M.L.; Küppers, R. Mechanisms of aberrant GATA3 expression in classical Hodgkin lymphoma and its consequences for the cytokine profile of Hodgkin and Reed/Sternberg cells. Blood 2010, 116, 4202–4211. [Google Scholar] [CrossRef] [PubMed]
- Atayar, C.; Poppema, S.; Blokzijl, T.; Harms, G.; Boot, M.; van den Berg, A. Expression of the T-cell transcription factors, GATA-3 and T-bet, in the neoplastic cells of Hodgkin lymphomas. Am. J. Pathol. 2005, 166, 127–134. [Google Scholar] [CrossRef]
- Kezlarian, B.; Alhyari, M.; Venkataraman, G.; Karner, K.; Inamdar, K.V.; Menon, M.P. GATA3 Immunohistochemical Staining in Hodgkin Lymphoma: Diagnostic Utility in Differentiating Classic Hodgkin Lymphoma from Nodular Lymphocyte Predominant Hodgkin Lymphoma and Other Mimicking Entities. Appl. Immunohistochem. Mol. Morphol. 2019, 27, 180–184. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, H.K.; Park, G.; Min, S.K.; Cha, H.J.; Lee, H.; Choi, S.J.; Na, H.Y.; Choe, J.Y.; Kim, J.E. Comparative pathologic analysis of mediastinal B-cell lymphomas: Selective expression of p63 but no GATA3 optimally differentiates primary mediastinal large B-cell lymphoma from classic Hodgkin lymphoma. Diagn. Pathol. 2019, 14, 133. [Google Scholar] [CrossRef] [PubMed]
- Siemer, D.; Kurth, J.; Lang, S.; Lehnerdt, G.; Stanelle, J.; Küppers, R. EBV transformation overrides gene expression patterns of B cell differentiation stages. Mol. Immunol. 2008, 45, 3133–3141. [Google Scholar] [CrossRef]
- Kyriazopoulou, L.; Karpathiou, G.; Hatzimichael, E.; Peoc’h, M.; Papoudou-Bai, A.; Kanavaros, P. Autophagy and cellular senescence in classical Hodgkin lymphoma. Pathol. Res. Pract. 2022, 236, 153964. [Google Scholar] [CrossRef]
- Papoudou-Bai, A.; Bai, M.; Doukas, M.; Stefanaki, K.; Spyridonos, P.; Karatzias, G.; Barbouti, A.; Galani, V.; Kanavaros, P. Immunohistological characterization of thymic dendritic cells. In Vivo 2012, 26, 985–992. [Google Scholar]
- Papoudou-Bai, A.; Hatzimichael, E.; Kyriazopoulou, L.; Briasoulis, E.; Kanavaros, P. Rare variants in the spectrum of human herpesvirus 8/Epstein-Barr virus-copositive lymphoproliferations. Hum. Pathol. 2015, 46, 1566–1571. [Google Scholar] [CrossRef]
- Bargou, R.C.; Emmerich, F.; Krappmann, D.; Bommert, K.; Mapara, M.Y.; Arnold, W.; Royer, H.D.; Grinstein, E.; Greiner, A.; Scheidereit, C.; et al. Constitutive nuclear factor-kappaB-RelA activation is required for proliferation and survival of Hodgkin’s disease tumor cells. J. Clin. Investig. 1997, 100, 2961–2969. [Google Scholar] [CrossRef]
- Hinz, M.; Lemke, P.; Anagnostopoulos, I.; Hacker, C.; Krappmann, D.; Mathas, S.; Dörken, B.; Zenke, M.; Stein, H.; Scheidereit, C. Nuclear factor kappaB-dependent gene expression profiling of Hodgkin’s disease tumor cells, pathogenetic significance, and link to constitutive signal transducer and activator of transcription 5a activity. J. Exp. Med. 2002, 196, 605–617. [Google Scholar] [CrossRef]
- Jundt, F.; Acikgöz, O.; Kwon, S.H.; Schwarzer, R.; Anagnostopoulos, I.; Wiesner, B.; Mathas, S.; Hummel, M.; Stein, H.; Reichardt, H.M.; et al. Aberrant expression of Notch1 interferes with the B-lymphoid phenotype of neoplastic B cells in classical Hodgkin lymphoma. Leukemia 2008, 22, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
- Jundt, F.; Anagnostopoulos, I.; Förster, R.; Mathas, S.; Stein, H.; Dörken, B. Activated Notch1 signaling promotes tumor cell proliferation and survival in Hodgkin and anaplastic large cell lymphoma. Blood 2002, 99, 3398–3403. [Google Scholar] [CrossRef]
- Barth, T.F.; Martin-Subero, J.I.; Joos, S.; Menz, C.K.; Hasel, C.; Mechtersheimer, G.; Parwaresch, R.M.; Lichter, P.; Siebert, R.; Mooller, P. Gains of 2p involving the REL locus correlate with nuclear c-Rel protein accumulation in neoplastic cells of classical Hodgkin lymphoma. Blood 2003, 101, 3681–3686. [Google Scholar] [CrossRef]
- Nam-Cha, S.H.; Montes-Moreno, S.; Salcedo, M.T.; Sanjuan, J.; Garcia, J.F.; Piris, M.A. Lymphocyte-rich classical Hodgkin’s lymphoma: Distinctive tumor and microenvironment markers. Mod. Pathol. 2009, 22, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Chronowski, G.M.; Wilder, R.B.; Tucker, S.L.; Ha, C.S.; Sarris, A.H.; Hagemeister, F.B.; Barista, I.; Hess, M.A.; Cabanillas, F.; Cox, J.D. An elevated serum beta-2-microglobulin level is an adverse prognostic factor for overall survival in patients with early-stage Hodgkin disease. Cancer 2002, 95, 2534–2538. [Google Scholar] [CrossRef] [PubMed]
- Höflinger, S.; Kesavan, K.; Fuxa, M.; Hutter, C.; Heavey, B.; Radtke, F.; Busslinger, M. Analysis of Notch1 function by in vitro T cell differentiation of Pax5 mutant lymphoid progenitors. J. Immunol. 2004, 173, 3935–3944. [Google Scholar] [CrossRef]
- Strobl, L.J.; Höfelmayr, H.; Stein, C.; Marschall, G.; Brielmeier, M.; Laux, G.; Bornkamm, G.W.; Zimber-Strobl, U. Both Epstein-Barr viral nuclear antigen 2 (EBNA2) and activated Notch1 transactivate genes by interacting with the cellular protein RBP-J. kappa. Immunobiology 1997, 198, 299–306. [Google Scholar] [CrossRef]
- Vassilakopoulos, T.P.; Nadali, G.; Angelopoulou, M.K.; Dimopoulou, M.N.; Siakantaris, M.P.; Kontopidou, F.N.; Karkantaris, C.; Kokoris, S.I.; Dimitriadou, E.M.; Calpadaki, C.; et al. beta(2)-microglobulin in Hodgkin’s lymphoma: Prognostic significance in patients treated with ABVD or equivalent regimens. J. Buon 2005, 10, 59–69. [Google Scholar]
- Zander, T.; Wiedenmann, S.; Wolf, J. Prognostic factors in Hodgkin’s lymphoma. Ann. Oncol. 2002, 13 (Suppl. S1), 67–74. [Google Scholar] [CrossRef]
- Itoh, K.; Kinoshita, T.; Watanabe, T.; Yoshimura, K.; Okamoto, R.; Chou, T.; Ogura, M.; Hirano, M.; Asaoku, H.; Kurosawa, M.; et al. Prognostic analysis and a new risk model for Hodgkin lymphoma in Japan. Int. J. Hematol. 2010, 91, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Cuccaro, A.; Bartolomei, F.; Cupelli, E.; Galli, E.; Giachelia, M.; Hohaus, S. Prognostic factors in hodgkin lymphoma. Mediterr. J. Hematol. Infect. Dis. 2014, 6, e2014053. [Google Scholar] [CrossRef] [PubMed]




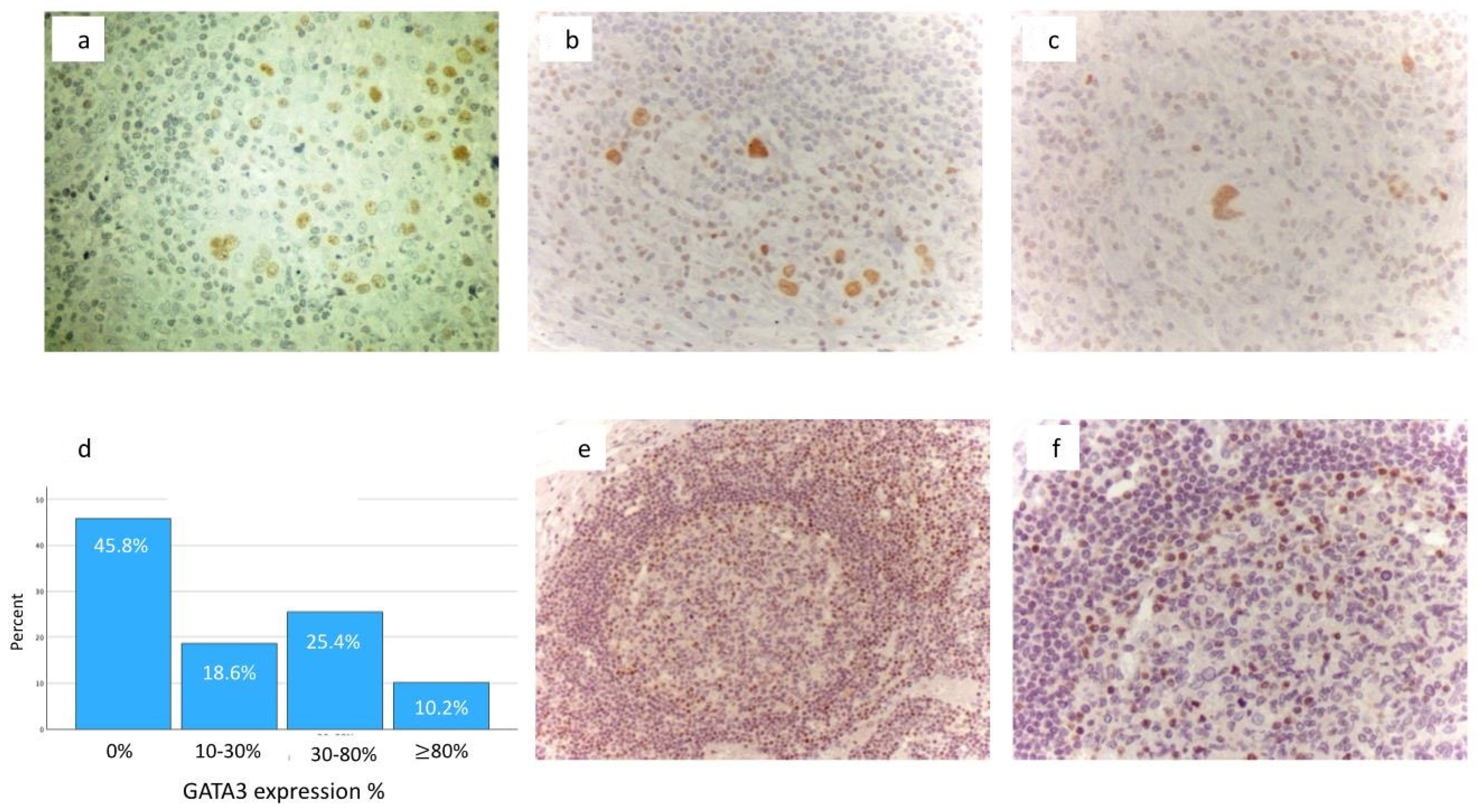{kind=link}
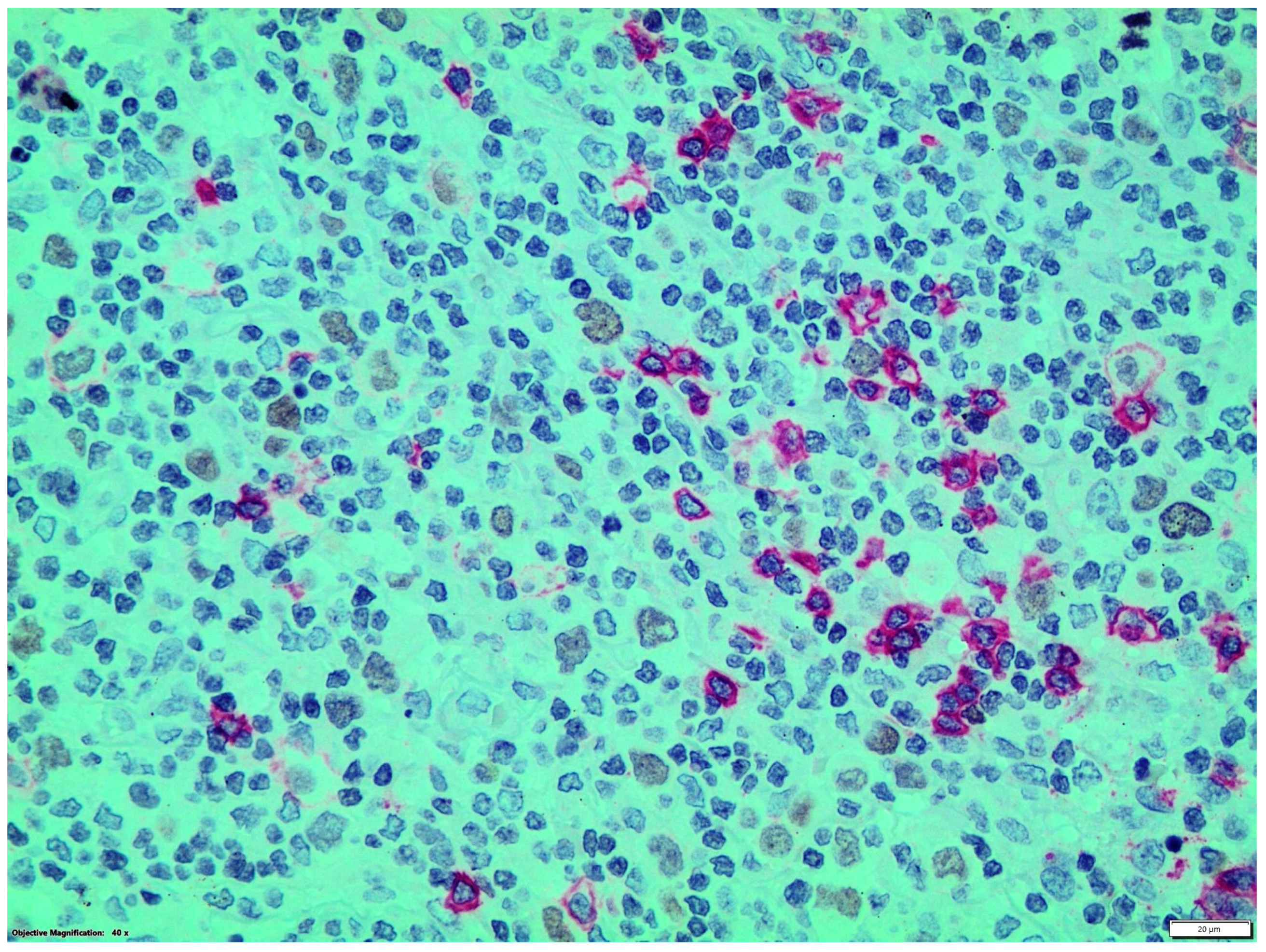{kind=link}
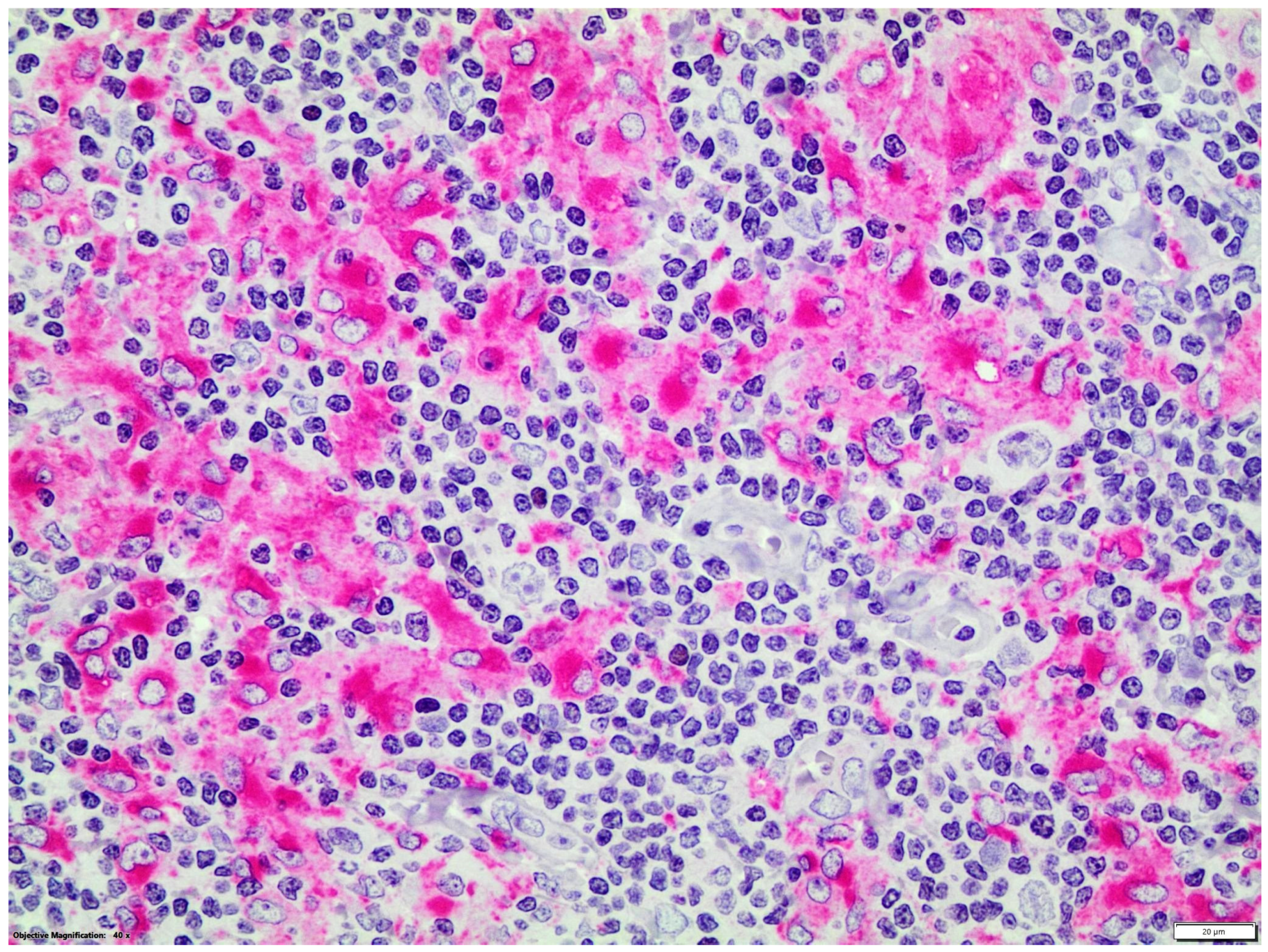{kind=link}
| Parameter | No GATA3 Expression | GATA3 Expression | p-Value (χ2 or Fisher Exact Test) |
|---|---|---|---|
| Age of diagnosis > 45 years old | 21/27 (77.8%) | 14/32 (43.8%) | 0.017 |
| Hemoglobin < 10.5 g/dL | 3/27(11.1%) | 8/32 (25%) | 0.2 |
| Lymphopenia (<0.6 × 109/L) | 2/27 (7.4%) | 3/32 (9.4%) | 1.0 |
| White Blood Cells > 15 × 109/L | 3/27 (11.1%) | 4/32 (12.5%) | 1.0 |
| Increased LDH | 12/26 (46.2%) | 18/31 (54.4%) | 0.36 |
| Albumin < 4 mg/dL | 14/26 (53.8%) | 18/31 (58.1%) | 0.74 |
| β2-microglobulin increased | 6/21 (28.6%) | 3/30 (10%) | 0.14 |
| Male | 16/27(59.3%) | 17/32 (53.1%) | 0.83 |
| B symptoms | 12/27 (44.4%) | 18/32 (56.3%) | 0.52 |
| Lugano stage advanced | 18/27 (66.7%) | 19/32 (59.4%) | 0.56 |
| Bulky disease | 4/27 (14.8%) | 7/32 (21.9%) | 0.52 |
| Positive interim PET/CT | 2/7 (28.7%) | 7/19 (36.8%) | 1.0 |
| Progressive disease/primary refractory | 5/25 (20%) | 13/31 (41.9%) | 0.14 |
| ESR > 50 mm/h | 12/26 (46.2%) | 13/31 (41.9%) | 0.75 |
| Extranodal disease | 10/27 (37%) | 8/32 (25%) | 0.31 |
| Parameter | No GATA3 Expression | GATA3 Expression >26% | p-Value (χ2 or Fisher Exact Test) |
|---|---|---|---|
| Age of diagnosis > 45 years old | 30/38 (78.9%) | 5/21 (23.8%) | <0.001 |
| Hemoglobin < 10.5 g/dL | 6/38(15.8%) | 5/21 (23.8%) | 1.0 |
| Lymphopenia (<0.6 × 109/L) | 3/38 (7.9%) | 2/21 (9.5%) | 1.0 |
| White Blood Cells > 15 × 109/L | 6/38 (15.8%) | 1/21 (5%) | 0.4 |
| Increased LDH | 16/37 (43.2%) | 14/20 (70%) | 0.098 |
| Albumin < 4 mg/dL | 21/37 (56.8%) | 11/20 (55%) | 1.0 |
| β2-microglobulin increased | 9/32 (28.1%) | 0/19 (0%) | 0.018 |
| Male | 21/38(55.3%) | 12/21 (57.1%) | 1.0 |
| B symptoms | 17/38 (44.7%) | 13/21 (61.9%) | 0.32 |
| Lugano stage advanced | 25/39 (64.1%) | 12/20 (60%) | 0.98 |
| Bulky disease | 6/38 (15.8%) | 5/21 (23.8%) | 0.68 |
| Positive interim PET/CT | 4/10 (40%) | 5/16 (31.3%) | 0.69 |
| Progressive disease/primary refractory | 8/35 (22.9%) | 10/21 (47.6%) | 0.1 |
| ESR > 50 mm/h | 17/37 (43.2%) | 9/20 (45%) | 1.0 |
| Extranodal disease | 12/38 (31.6%) | 6/21 (28.6%) | 1.0 |
| TBIL increased | 7/38 (18.4%) | 0/20 (0%) | 0.08 |
| Characteristic | 5 Year-OS (%) | p-Value |
|---|---|---|
| GATA 3 (−) | 79% | 0.99 |
| GATA 3 (+) | 77% | |
| GATA 3 < 26% | 79% | 0.97 |
| GATA 3 ≥ 26% | 60% | |
| Age of diagnosis ≤ 45 years old | 77% | 0.18 |
| Age of diagnosis > 45 years old | 72% | |
| Hb ≥ 10.5 g/dL | 79% | 0.13 |
| Hb < 10.5 g/dL | 53% | |
| Lymphopenia (≥0.6 × 109/L) | 75% | 0.66 |
| Lymphopenia (<0.6 × 109/L) | 75% | |
| Female | 83% | 0.4 |
| Male | 67% | |
| White Blood Cells ≤ 15 × 109/L | 73% | 0.58 |
| White Blood Cells > 15 × 109/L | 85% | |
| Normal LDH | 72% | 0.85 |
| Increased LDH | 76% | |
| Albumin ≥ 4 mg/dL | 82% | 0.12 |
| Albumin < 4 mg/dL | 67% | |
| β2-microglobulin normal | 83% | <0.001 |
| β2-microglobulin increased | 27% | |
| No B symptoms | 90% | 0.008 |
| B symptoms | 59% | |
| Lugano stage limited | 82% | 0.18 |
| Lugano stage advanced | 71% | |
| No Bulky disease | 73% | 0.56 |
| Bulky disease | 83% | |
| Negative interim PET/CT | 65% | 0.2 |
| Positive interim PET/CT | 47% | |
| No Progressive/primary refractory disease | 85% | 0.19 |
| Progressive disease/primary refractory | 61% | |
| ESR≤ 50 mm/h | 72% | 0.76 |
| ESR > 50 mm/h | 77% | |
| No Extranodal disease | 70% | 0.35 |
| Extranodal disease | 87% | |
| TBIL normal | 77% | 0.05 |
| TBIL elevated | 54% |
| Characteristic | Hazard Ratio | 95% Cl | p-Value |
|---|---|---|---|
| β2-microglobulin increased | 14.6 | 1.6–133.6 | 0.018 |
| B symptoms | 9.8 | 1.18–83.2 | 0.035 |
| TBIL increased | 1.2 | 0.21–7.29 | 0.81 |
| GATA3 ≥ 26% | 1.45 | 0.23–8.9 | 0.69 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papoudou-Bai, A.; Koumpis, E.; Karpathiou, G.; Hatzimichael, E.; Kanavaros, P. Expression Patterns of GATA3 in Classical Hodgkin Lymphoma: A Clinico-Pathological Study. Diseases 2024, 12, 51. https://doi.org/10.3390/diseases12030051
Papoudou-Bai A, Koumpis E, Karpathiou G, Hatzimichael E, Kanavaros P. Expression Patterns of GATA3 in Classical Hodgkin Lymphoma: A Clinico-Pathological Study. Diseases. 2024; 12(3):51. https://doi.org/10.3390/diseases12030051
Chicago/Turabian StylePapoudou-Bai, Alexandra, Epameinondas Koumpis, Georgia Karpathiou, Eleftheria Hatzimichael, and Panagiotis Kanavaros. 2024. "Expression Patterns of GATA3 in Classical Hodgkin Lymphoma: A Clinico-Pathological Study" Diseases 12, no. 3: 51. https://doi.org/10.3390/diseases12030051