
Yata Virus (Family Rhabdoviridae, Genus Ephemerovirus) Isolation from Mosquitoes from Uganda, the First Reported Isolation since 1969
 , , and
, , and 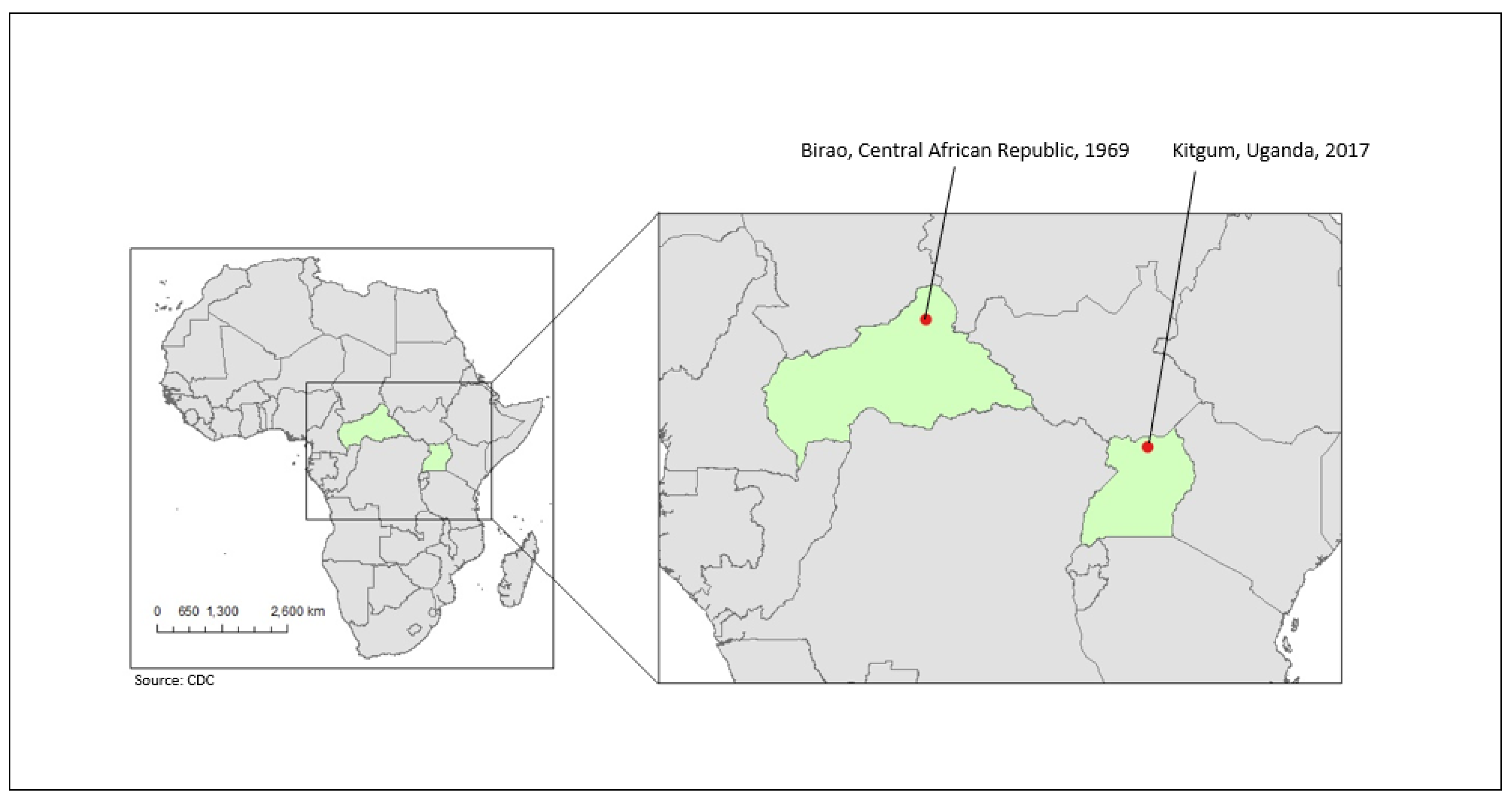{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Trituration and Infection Status Determination
2.2. RNA Sequencing
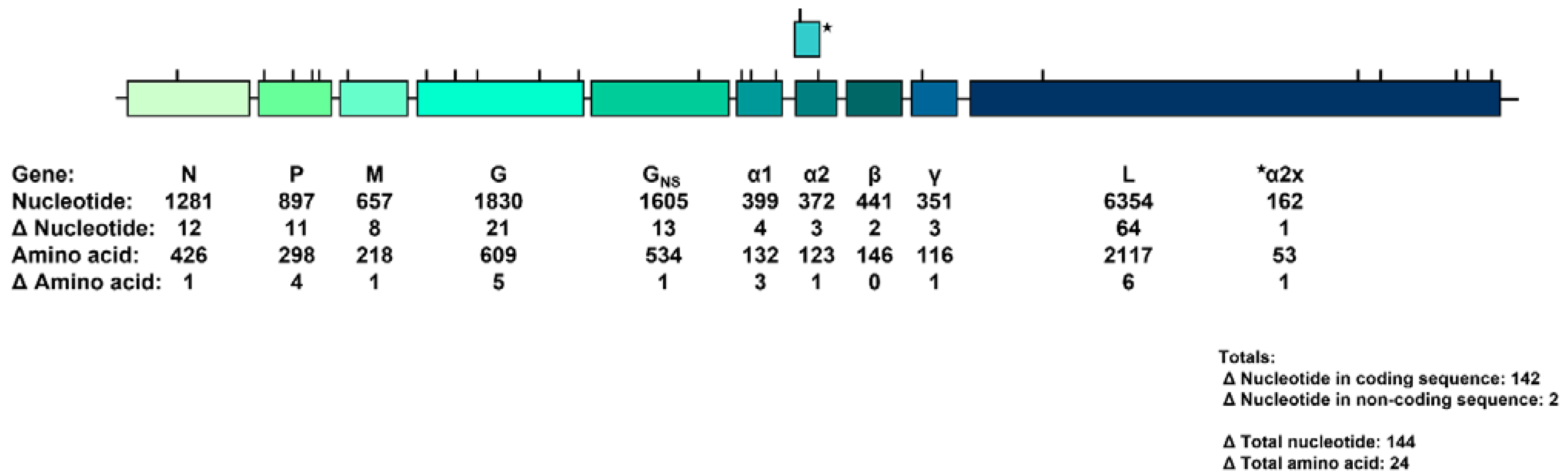
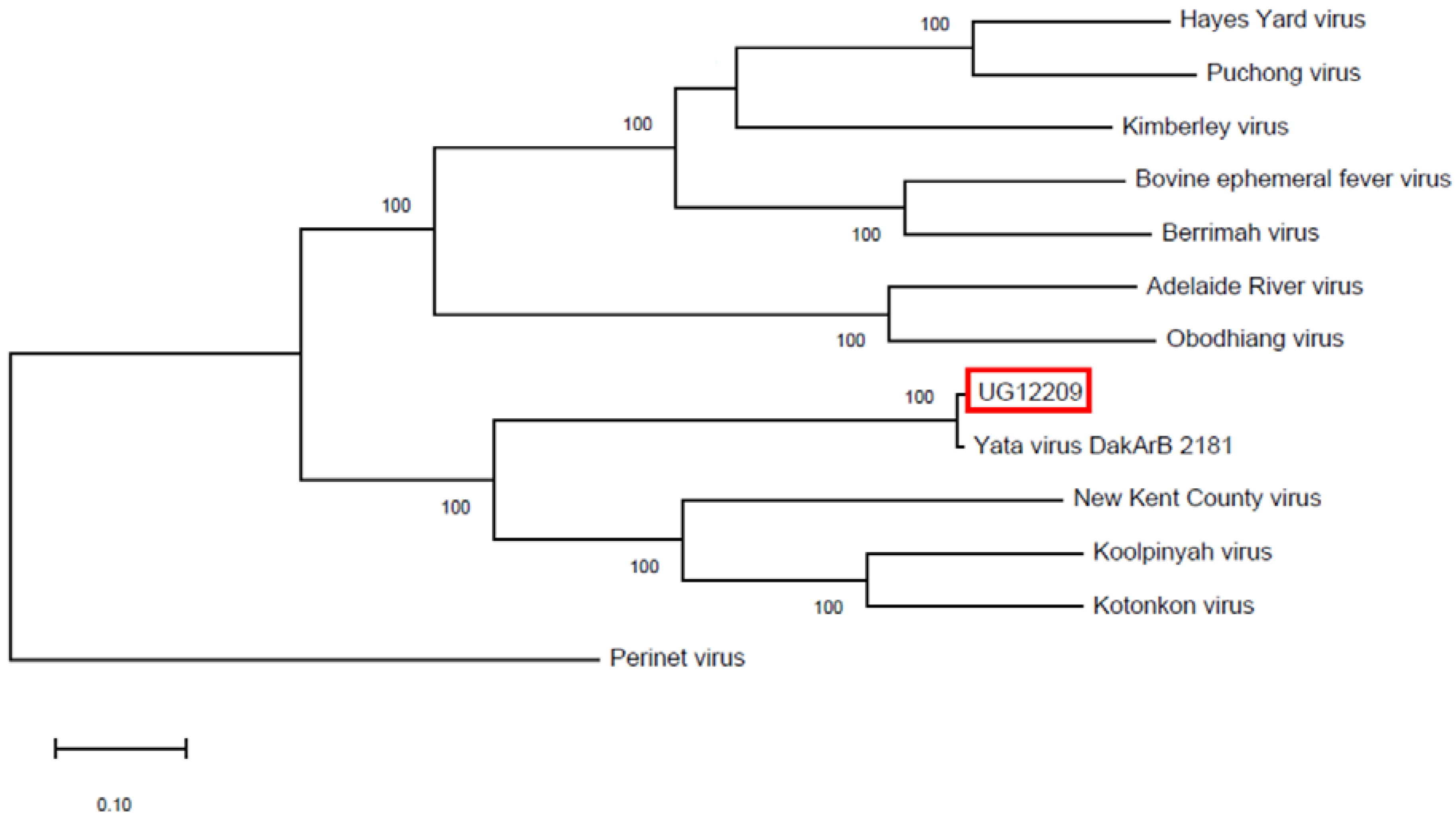
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, P.J.; Freitas-Astúa, J.; Bejerman, N.; Blasdell, K.R.; Breyta, R.; Dietzgen, R.G.; Fooks, A.R.; Kondo, H.; Kurath, G.; Kuzmin, I.V.; et al. ICTV Virus Taxonomy Profile: Rhabdoviridae 2022. J. Gen. Virol. 2022, 103, 001689. [Google Scholar] [CrossRef]
- Walker, P.J.; Firth, C.; Widen, S.G.; Blasdell, K.R.; Guzman, H.; Wood, T.G.; Paradkar, P.N.; Holmes, E.C.; Tesh, R.B.; Vasilakis, N. Evolution of genome size and complexity in the rhabdoviridae. PLoS Pathog. 2015, 11, e1004664. [Google Scholar] [CrossRef] [Green Version]
- Dietzgen, R.G.; Kondo, H.; Goodin, M.M.; Kurath, G.; Vasilakis, N. The family Rhabdoviridae: Mono- and bipartite negative-sense RNA viruses with diverse genome organization and common evolutionary origins. Virus Res. 2017, 227, 158–170. [Google Scholar] [CrossRef] [Green Version]
- Dietzgen, R.G.; Calisher, C.H.; Kurath, G.; Kuzmin, I.V.; Rodriguez, L.L.; Stone, D.M.; Tesh, R.B.; Tordo, N.; Walker, P.J.; Wetzel, T.; et al. Family Rhabdoviridae. In Virus Taxonomy; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: Oxford, UK, 2011; pp. 686–714. [Google Scholar]
- Walker, P.J.; Dietzgen, R.G.; Joubert, D.A.; Blasdell, K.R. Rhabdovirus accessory genes. Virus Res. 2011, 162, 110–125. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Novella, I.S.; Dietzgen, R.G.; Padhi, A.; Rupprecht, C.E. The rhabdoviruses: Biodiversity, phylogenetics, and evolution. Infect. Genet. Evol. 2009, 9, 541–553. [Google Scholar] [CrossRef]
- Blasdell, K.R.; Davis, S.S.; Voysey, R.; Bulach, D.M.; Middleton, D.; Williams, S.; Harmsen, M.B.; Weir, R.P.; Crameri, S.; Walsh, S.J.; et al. Hayes Yard virus: A novel ephemerovirus isolated from a bull with severe clinical signs of bovine ephemeral fever is most closely related to Puchong virus. Vet Res. 2020, 51, 58. [Google Scholar] [CrossRef]
- Dacheux, L.; Dommergues, L.; Chouanibou, Y.; Doméon, L.; Schuler, C.; Bonas, S.; Luo, D.; Maufrais, C.; Cetre-Sossah, C.; Cardinale, E.; et al. Co-circulation and characterization of novel African arboviruses (genus Ephemerovirus) in cattle, Mayotte island, Indian Ocean, 2017. Transbound Emerg. Dis. 2019, 66, 2601–2604. [Google Scholar] [CrossRef] [Green Version]
- Rapport Annuel de l’Institut Pasteur de Bangui, 1969, pp. 51–52.
- Calisher, C.H.; Karabatsos, N.; Zeller, H.; Digoutte, J.P.; Tesh, R.B.; Shope, R.E.; Travassos da Rosa, A.P.; St George, T.D. Antigenic relationships among rhabdoviruses from vertebrates and hematophagous arthropods. Intervirology 1989, 30, 241–257. [Google Scholar] [CrossRef]
- Tesh, R.B.; Travassos Da Rosa, A.P.; Travassos Da Rosa, J.S. Antigenic relationship among rhabdoviruses infecting terrestrial vertebrates. J. Gen. Virol. 1983, 64 Pt 1, 169–176. [Google Scholar] [CrossRef]
- Blasdell, K.R.; Widen, S.G.; Diviney, S.M.; Firth, C.; Wood, T.G.; Guzman, H.; Holmes, E.C.; Tesh, R.B.; Vasilakis, N.; Walker, P.J. Koolpinyah and Yata viruses: Two newly recognised ephemeroviruses from tropical regions of Australia and Africa. Vet. Microbiol. 2014, 174, 547–553. [Google Scholar] [CrossRef]
- Mossel, E.C.; Crabtree, M.B.; Mutebi, J.P.; Lutwama, J.J.; Borland, E.M.; Powers, A.M.; Miller, B.R. Arboviruses Isolated From Mosquitoes Collected in Uganda, 2008–2012. J. Med. Entomol. 2017, 54, 1403–1409. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Novella, I.S.; Presloid, J.B.; Zhou, T.; Smith-Tsurkan, S.D.; Ebendick-Corpus, B.E.; Dutta, R.N.; Lust, K.L.; Wilke, C.O. Genomic evolution of vesicular stomatitis virus strains with differences in adaptability. J. Virol. 2010, 84, 4960–4968. [Google Scholar] [CrossRef] [Green Version]
- Novella, I.S.; Presloid, J.B.; Smith, S.D.; Wilke, C.O. Specific and nonspecific host adaptation during arboviral experimental evolution. J. Mol. Microbiol. Biotechnol. 2011, 21, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Trinidad, L.; Blasdell, K.R.; Joubert, D.A.; Davis, S.S.; Melville, L.; Kirkland, P.D.; Coulibaly, F.; Holmes, E.C.; Walker, P.J. Evolution of bovine ephemeral fever virus in the Australian episystem. J. Virol. 2014, 88, 1525–1535. [Google Scholar] [CrossRef] [Green Version]
- Gard, G.P.; Melville, L.F.; Calisher, C.H.; Karabatsos, N. Koolpinyah: A virus related to kotonkan from cattle in northern Australia. Intervirology 1992, 34, 142–145. [Google Scholar]
- Kemp, G.E.; Lee, V.H.; Moore, D.L.; Shope, R.E.; Causey, O.R.; Murphy, F.A. Kotonkan, a new rhabdovirus related to Mokola virus of the rabies serogroup. Am. J. Epidemiol. 1973, 98, 43–49. [Google Scholar] [CrossRef]
- Tomori, O.; Fagbami, A.; Kemp, G. Kotonkan virus: Experiment infection of white Fulani calves. Bull. Epizoot. Dis. Afr. 1974, 22, 195–200. [Google Scholar]
- Ba, Y.; Diallo, D.; Dia, I.; Diallo, M. Comportement trophique des vecteurs du virus de la fièvre de la vallée du Rift au Sénégal: Implications dans l’épidémiologie de la maladie. Bull. Soc. Pathol. Exot. 2006, 99, 283–289. [Google Scholar]
- Chandler, J.A.; Boreham, P.F.; Highton, R.B.; Hill, M.N. Feeding habits of mosquitoes in the Kisumu area and their possible relationship to disease transmission. East Afr. Med. J. 1975, 52, 413–417. [Google Scholar]
- Chandler, J.A.; Boreham, P.F.; Highton, R.B.; Hill, M.N. A study of the host selection patterns of the mosquitoes of the Kisumu area of Kenya. Trans. R. Soc. Trop. Med. Hyg. 1975, 69, 415–425. [Google Scholar] [CrossRef]
- Crabtree, M.B.; Kading, R.C.; Mutebi, J.P.; Lutwama, J.J.; Miller, B.R. Identification of host blood from engorged mosquitoes collected in western Uganda using cytochrome oxidase I gene sequences. J. Wildl. Dis. 2013, 49, 611–626. [Google Scholar] [CrossRef]
- Beier, J.C.; Odago, W.O.; Onyango, F.K.; Asiago, C.M.; Koech, D.K.; Roberts, C.R. Relative abundance and blood feeding behavior of nocturnally active culicine mosquitoes in western Kenya. J. Am. Mosq Control Assoc. 1990, 6, 207–212. [Google Scholar]
- Paterson, H.E.; Bronsden, P.; Levitt, J.; Worth, C.B. Some Culicine Mosquitoes (Diptera, Culicidae) at Ndumu, Republic of South Africa: A Study of Their Host Preferences and Host Range. Med. Proc. 1964, 10, 188–192. [Google Scholar]
- International Catalogue of Arboviruses: Including Certain Other Viruses of Vertebrates, 3rd ed.; American Society of Tropical medicine and Hygiene: San Antonio, TX, USA, 1985; p. 1147.



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perinet, L.C.; Mutebi, J.-P.; Powers, A.M.; Lutwama, J.J.; Mossel, E.C. Yata Virus (Family Rhabdoviridae, Genus Ephemerovirus) Isolation from Mosquitoes from Uganda, the First Reported Isolation since 1969. Diseases 2023, 11, 21. https://doi.org/10.3390/diseases11010021
Perinet LC, Mutebi J-P, Powers AM, Lutwama JJ, Mossel EC. Yata Virus (Family Rhabdoviridae, Genus Ephemerovirus) Isolation from Mosquitoes from Uganda, the First Reported Isolation since 1969. Diseases. 2023; 11(1):21. https://doi.org/10.3390/diseases11010021
Chicago/Turabian StylePerinet, Lara C., John-Paul Mutebi, Ann M. Powers, Julius J. Lutwama, and Eric C. Mossel. 2023. "Yata Virus (Family Rhabdoviridae, Genus Ephemerovirus) Isolation from Mosquitoes from Uganda, the First Reported Isolation since 1969" Diseases 11, no. 1: 21. https://doi.org/10.3390/diseases11010021