
Approximate Entropy of Spiking Series Reveals Different Dynamical States in Cortical Assemblies


{kind=link}
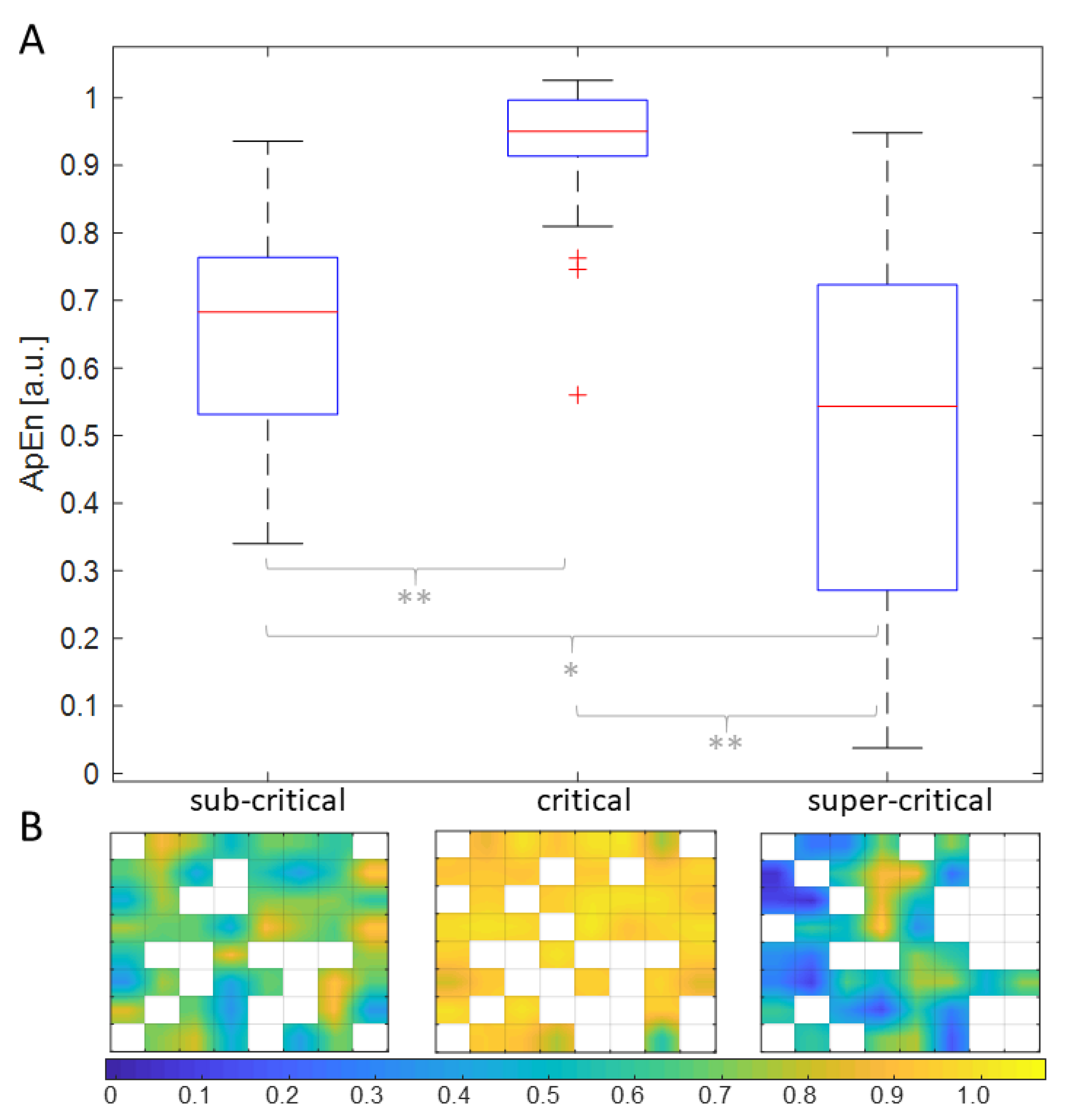{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
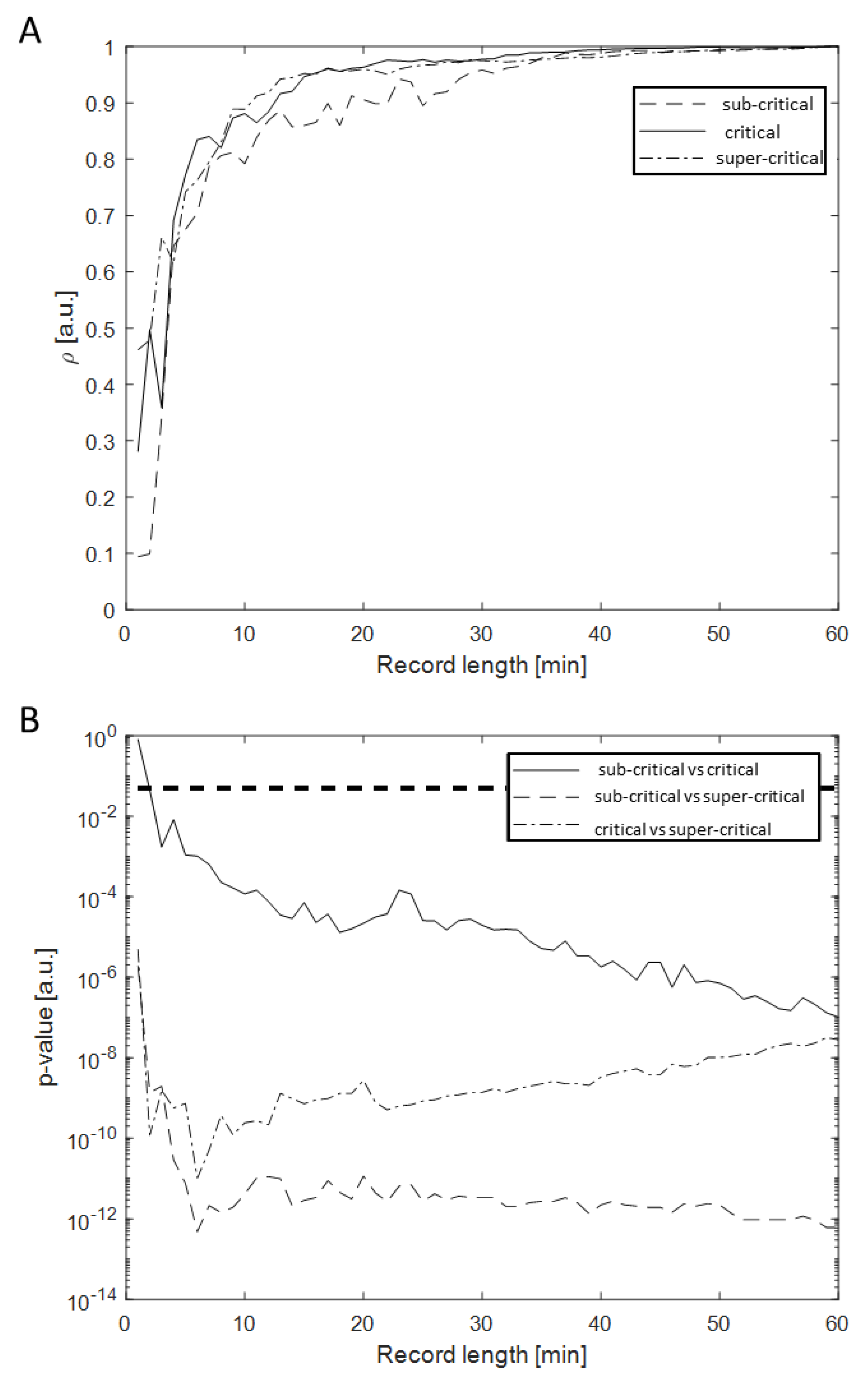{kind=link}
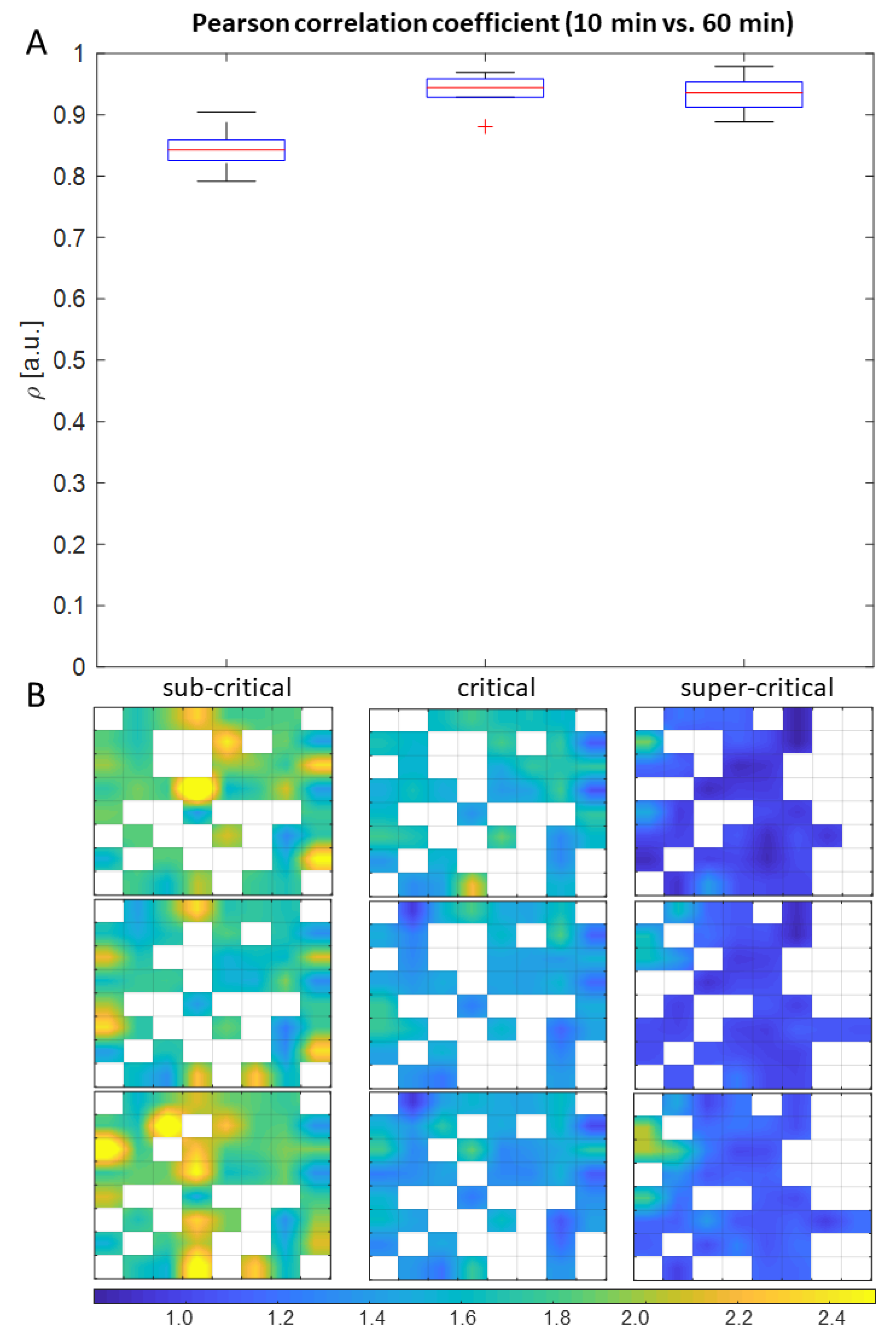{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Experimental Preparations
2.2. Extraction of Spiking Activity
2.3. Approximate Entropy
2.4. Statistical Analysis
3. Results
4. Discussion
4.1. Comparison with Literature
4.2. Interpretation of Our Results
5. Conclusions and Further Work
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ApEn | approximate entropy |
| ISI | inter-spike interval |
| MEA | micro-electrode array |
| SampEn | sample entropy |
| SOC | self-organized criticality |
| std | standard deviation |
Appendix A



References
- Abbott, L.F.; Dayan, P. Theoretical Neuroscience: Computational and Mathematical Modelling of Neural System; The MIT Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Bove, M.; Grattarola, M.; Verreschi, G. In vitro 2D networks of neurons characterized by processing the signals recorded with a planar microtransducer array. IEEE Trans. Biomed. Eng. 1997, 44, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Rodieck, R.W.; Kiang, N.Y.; Gerstein, G.L. Some quantitative methods for the study of spontaneous activity of single neurons. Biophys. J. 1962, 2, 351–368. [Google Scholar] [CrossRef] [Green Version]
- Aertsen, A.M.; Gerstein, G.L.; Habib, M.K.; Palm, G. Dynamics of neuronal firing correlation: Modulation of effective connectivity. J. Neurophysiol. 1989, 61, 900–917. [Google Scholar] [CrossRef]
- Brown, E.N.; Kass, R.E.; Mitra, P.P. Multiple neural spike train data analysis: State-of-the-art and future challenges. Nat. Neurosci. 2004, 7, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Cocchi, L.; Gollo, L.L.; Zalesky, A.; Breakspear, M. Criticality in the brain: A synthesis of neurobiology, models and cognition. Prog. Neurobiol. 2017, 158, 132–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Destexhe, A.; Touboul, J.D. Is There Sufficient Evidence for Criticality in Cortical Systems? eNeuro 2021, 8, ENEURO.0551-20.2021. [Google Scholar] [CrossRef] [PubMed]
- Beggs, J.M.; Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 2003, 23, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Beggs, J.M. The criticality hypothesis: How local cortical networks might optimize information processing. Philos. Trans. R. Soc. A 2008, 366, 329–343. [Google Scholar] [CrossRef]
- Cover, T.M.; Thomas, J.A. Elements of Information Theory, 2nd ed.; Wiley-Interscience: Hoboken, NJ, USA, 2006. [Google Scholar]
- Pincus, S.M. Approximate entropy as a measure of system complexity. Proc. Natl. Acad. Sci. USA 1991, 88, 2297–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.J.; Hu, S.J.; Han, S.; Liu, G.P.; Xie, Y.; Xu, J.X. Relation between responsiveness to neurotransmitters and complexity of epileptiform activity in hippocampal CA1 neurons. Epilepsia 2002, 43, 1330–1336. [Google Scholar] [CrossRef] [Green Version]
- Akay, M.; Sekine, N. Investigating the complexity of respiratory patterns during recovery from severe hypoxia. J. Neural Eng. 2004, 1, 16–20. [Google Scholar] [CrossRef]
- Richman, J.S.; Moorman, J.R. Physiological time-series analysis using approximate entropy and sample entropy. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H2039–H2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapucu, F.E.; Mikkonen, J.E.; Tanskanen, J.M.; Hyttinen, J.A. Quantification and automatized adaptive detection of in vivo and in vitro neuronal bursts based on signal complexity. In Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25–29 August 2015; pp. 4729–4732. [Google Scholar]
- Kapucu, F.E.; Valkki, I.; Christophe, F.; Tanskanen, J.M.A.; Johansson, J.; Mikkonen, T.; Hyttinen, J.A.K. On electrophysiological signal complexity during biological neuronal network development and maturation. In Proceedings of the 2017 39th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Jeju, Korea, 11–15 July 2017; pp. 3333–3338. [Google Scholar]
- Chen, L.; Luo, W.; Deng, Y.; Wang, Z.; Zeng, S. Characterizing the complexity of spontaneous electrical signals in cultured neuronal networks using approximate entropy. IEEE Trans. Inf. Technol. Biomed. 2009, 13, 405–410. [Google Scholar] [CrossRef]
- Dorval, A.D.; Russo, G.S.; Hashimoto, T.; Xu, W.; Grill, W.M.; Vitek, J.L. Deep brain stimulation reduces neuronal entropy in the MPTP-primate model of Parkinson’s disease. J. Neurophysiol. 2008, 100, 2807–2818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafreniere-Roula, M.; Darbin, O.; Hutchison, W.D.; Wichmann, T.; Lozano, A.M.; Dostrovsky, J.O. Apomorphine reduces subthalamic neuronal entropy in parkinsonian patients. Exp. Neurol. 2010, 225, 455–458. [Google Scholar] [CrossRef] [Green Version]
- Darbin, O.; Jin, X.; Von Wrangel, C.; Schwabe, K.; Nambu, A.; Naritoku, D.K.; Krauss, J.K.; Alam, M. Neuronal Entropy-Rate Feature of Entopeduncular Nucleus in Rat Model of Parkinson’s Disease. Int. J. Neural Syst. 2016, 26, 1550038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ermini, L.; Mesin, L.; Massobrio, P. Approximate Entropy of Spiking Series of a Neuronal Network in either Subcritical or Critical state. In Proceedings of the 2018 IEEE Workshop on Complexity in Engineering (COMPENG), Florence, Italy, 10–12 October 2018. [Google Scholar]
- Brofiga, M.; Pisano, M.; Tedesco, M.; Boccaccio, A.; Massobrio, P. Functional Inhibitory Connections Modulate the Electrophysiological Activity Patterns of Cortical-Hippocampal Ensembles. Cereb. Cortex 2021, bhab318. [Google Scholar] [CrossRef] [PubMed]
- Maccione, A.; Gandolfo, M.; Massobrio, P.; Novellino, A.; Martinoia, S.; Chiappalone, M. A novel algorithm for precise identification of spikes in extracellularly recorded neuronal signals. J. Neurosci. Methods 2009, 177, 241–249. [Google Scholar] [CrossRef]
- Chiarion, G.; Mesin, L. Resolution of Spike Overlapping by Biogeography-Based Optimization. Electronics 2021, 10, 1469. [Google Scholar] [CrossRef]
- Wagenaar, D.A.; Nadasdy, Z.; Potter, S.M. Persistent dynamic attractors in activity patterns of cultured neuronal networks. Phys. Rev. E 2006, 73, 051907. [Google Scholar] [CrossRef] [Green Version]
- Mesin, L. Estimation of Complexity of Sampled Biomedical Continuous Time Signals using Approximate Entropy. Front. Physiol. 2018, 9, 710. [Google Scholar] [CrossRef]
- Pincus, S.M.; Goldberger, A.L. Physiological time-series analysis: What does regularity quantify? Am. J. Physiol. Heart Circ. Physiol. 1994, 266, H1643–H1656. [Google Scholar] [CrossRef]
- Schreiber, T.; Schmitz, A. Improved surrogate data for nonlinearity tests. Phys. Rev. Lett. 1996, 77, 635–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massobrio, P.; Pasquale, V.; Martinoia, S. Self-organized criticality in cortical assemblies occurs in concurrent scale-free and small-world networks. Sci. Rep. 2015, 5, 10578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timme, N.M.; Marshall, N.J.; Bennett, N.; Ripp, M.; Lautzenhiser, E.; Beggs, J.M. Criticality maximizes complexity in neural tissue. Front. Physiol. 2016, 7, 425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Barkhaus, P.E.; Zhang, X.; Rymer, W.Z. Characterizing the complexity of spontaneous motor unit patterns of amyotrophic lateral sclerosis using approximate entropy. J. Neural Eng. 2011, 8, 066010. [Google Scholar] [CrossRef] [PubMed]
- Pajevic, S.; Plenz, D. Efficient network reconstruction from dynamical cascades identifies small-world topology of neuronal avalanches. PLoS Comput. Biol. 2009, 5, e1000271. [Google Scholar] [CrossRef] [Green Version]
- Castiglioni, P.; Faes, L.; Valenza, G. Assessing Complexity in Physiological Systems through Biomedical Signals Analysis. Entropy 2020, 22, 1005. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, R.K.; Deb, S.; Dandapat, S. Analysis of physiological signals using state space correlation entropy. Healthc. Technol. Lett. 2017, 4, 30–33. [Google Scholar] [CrossRef]
- Jamin, A.; Humeau-Heurtier, A. (Multiscale) Cross-Entropy Methods: A Review. Entropy 2020, 22, 45. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ermini, L.; Massobrio, P.; Mesin, L. Approximate Entropy of Spiking Series Reveals Different Dynamical States in Cortical Assemblies. Electronics 2022, 11, 936. https://doi.org/10.3390/electronics11060936
Ermini L, Massobrio P, Mesin L. Approximate Entropy of Spiking Series Reveals Different Dynamical States in Cortical Assemblies. Electronics. 2022; 11(6):936. https://doi.org/10.3390/electronics11060936
Chicago/Turabian StyleErmini, Leonardo, Paolo Massobrio, and Luca Mesin. 2022. "Approximate Entropy of Spiking Series Reveals Different Dynamical States in Cortical Assemblies" Electronics 11, no. 6: 936. https://doi.org/10.3390/electronics11060936