A Zebrafish Acromegaly Model Elevates DNA Damage and Impairs DNA Repair Pathways


Abstract
:1. Introduction
2. Materials and Methods
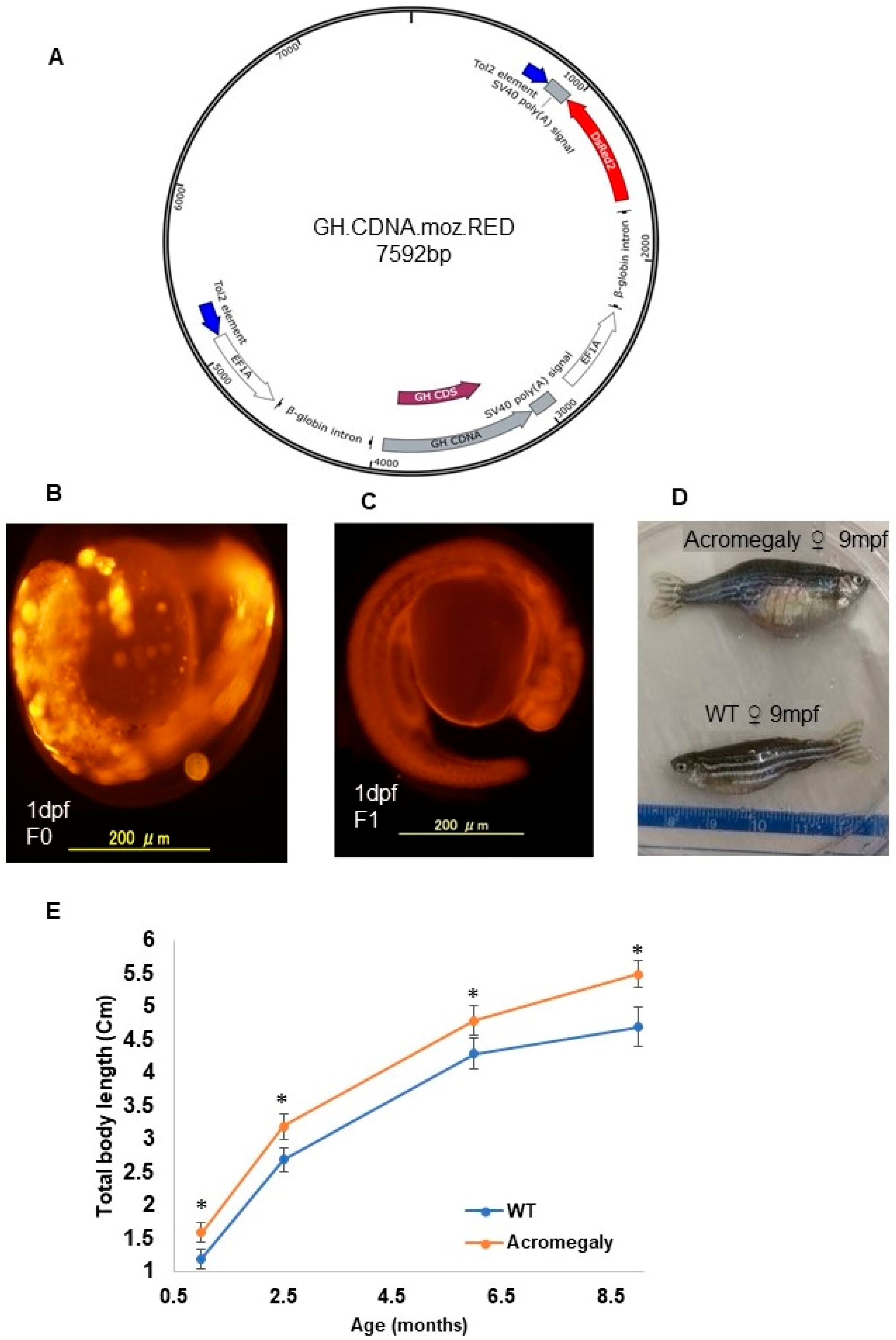
2.1. Transgene Construction
2.2. Production of the Acromegaly Zebrafish First Generation (F0)
2.3. Production of the Acromegaly Zebrafish Second Generation (F1)
2.4. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.5. Quantitative PCR (qPCR)
2.6. RNA-Seq and Data Analysis
RNA Isolation and Library Construction
2.7. Mapping and Identification of Differential Gene Expression
2.8. Expression2Kinases (X2K)
2.9. Heatmaps
2.10. Gene Set Enrichment Analysis (GSEA)
2.11. Isolation of Live Muscle and Brain Cells by Fluorescence-Activated Cell Sorting (FACS)
2.12. Immunocytochemistry of FACS-Isolated Live Cells
2.13. Whole-Mount Immunostaining
2.14. Acridine Orange Staining
2.15. Data Availability
3. Results
3.1. Tilapia GH is Physiologically Active in the Acromegaly Zebrafish Model
3.2. The Acromegaly Zebrafish Have Elevated DNA Damage
3.3. Impairment of DNA Repair Pathways in Acromegaly Zebrafish Liver and Induction of Apoptosis in Acromegaly Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chanson, P.; Salenave, S. Acromegaly. Orphanet J. Rare Dis. 2008, 3, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jallad, R.S.; Musolino, N.R.; Salgado, L.R.; Bronstein, M.D. Treatment of acromegaly: Is there still a place for radiotherapy? Pituitary 2007, 10, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, B.Å.; Edén, S.; Ernest, I.; Odén, A.; Sjögren, B. Epidemiology and Long-term Survival in Acromegaly: A Study of 166 Cases Diagnosed between 1955 and 1984. J. Intern. Med. 1988, 223, 327–335. [Google Scholar] [CrossRef]
- Velloso, C. Regulation of muscle mass by growth hormone and IGF-I. Br. J. Pharmacol. 2008, 154, 557–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, Y.; Sohmiya, M.; Nishiki, M. Regulation of human growth hormone secretion and its disorders. Intern. Med. 2002, 41, 7–13. [Google Scholar]
- Huang, E.; Zhu, G.; Jiang, W.; Yang, K.; Gao, Y.; Luo, Q.; Gao, J.L.; Kim, S.H.; Liu, X.; Li, M. Growth hormone synergizes with BMP9 in osteogenic differentiation by activating the JAK/STAT/IGF1 pathway in murine multilineage cells. J. Bone Miner. Res. 2012, 27, 1566–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poburski, D.; Leovsky, C.; Boerner, J.B.; Szimmtenings, L.; Ristow, M.; Glei, M.; Thierbach, R. Insulin-IGF signaling affects cell transformation in the BALB/c 3T3 cell model. Sci. Rep. 2016, 6, 37120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartke, A. Minireview: Role of the growth hormone/insulin-like growth factor system in mammalian aging. Endocrinology 2005, 146, 3718–3723. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Aguirre, J.; Balasubramanian, P.; Guevara-Aguirre, M.; Wei, M.; Madia, F.; Cheng, C.-W.; Hwang, D.; Martin-Montalvo, A.; Saavedra, J.; Ingles, S. Growth hormone receptor deficiency is associated with a major reduction in pro-aging signaling, cancer, and diabetes in humans. Sci. Transl. Med. 2011, 3, 70ra13. [Google Scholar] [CrossRef] [PubMed]
- Urasaki, A.; Morvan, G.; Kawakami, K. Functional Dissection of the Tol2 Transposable Element Identified the Minimal cis-Sequence and a Highly Repetitive Sequence in the Subterminal Region Essential for Transposition. Genetics 2006, 174, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K. Tol2: A versatile gene transfer vector in vertebrates. Genome Biol. 2007, 8, S7. [Google Scholar] [CrossRef] [PubMed]
- Suster, M.L.; Abe, G.; Schouw, A.; Kawakami, K. Transposon-mediated BAC transgenesis in zebrafish. Nat. Protoc. 2011, 6, 1998–2021. [Google Scholar] [CrossRef] [PubMed]
- Smedley, D.; Haider, S.; Ballester, B.; Holland, R.; London, D.; Thorisson, G.; Kasprzyk, A. BioMart–biological queries made easy. BMC Genom. 2009, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Xu, H.; Gordonov, S.; Lim, M.P.; Perkins, M.H.; Ma’ayan, A. Expression2Kinases: MRNA profiling linked to multiple upstream regulatory layers. Bioinformatics 2011, 28, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milanowska, K.; Krwawicz, J.; Papaj, G.; Kosiński, J.; Poleszak, K.; Lesiak, J.; Osińska, E.; Rother, K.; Bujnicki, J.M. REPAIRtoire—A database of DNA repair pathways. Nucleic Acids Res. 2010, 39, D788–D792. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xu, H.; Lam, S.H.; Gong, Z. Selection of reliable biomarkers from PCR array analyses using relative distance computational model: Methodology and proof-of-concept study. PLoS ONE 2013, 8, e83954. [Google Scholar] [CrossRef] [PubMed]
- Froehlich, J.M.; Seiliez, I.; Gabillard, J.-C.; Biga, P.R. Preparation of Primary Myogenic Precursor Cell/Myoblast Cultures from Basal Vertebrate Lineages. J. Vis. Exp. 2014, e51354. [Google Scholar] [CrossRef] [PubMed]
- Morales, R.; Herrera, M.T.; Arenal, A.; Cruz, A.; Hernández, O.; Pimentel, R.; Guillén, I.; Martínez, R.; Estrada, M.P. Tilapia chromosomal growth hormone gene expression accelerates growth in transgenic zebrafish (Danio rerio). Electron. J. Biotechnol. 2001, 4, 5–6. [Google Scholar]
- Ross, R.; Tsagarakis, S.; Grossman, A.; Nhagafoong, L.; Touzel, R.; Rees, L.; Besser, G. GH feedback occurs through modulation of hypothalamic somatostatin under cholinergic control: Studies with pyridostigmine and GHRH. Clin. Endocrinol. 1987, 27, 727–733. [Google Scholar] [CrossRef]
- Rojas-Gil, A.P.; Ziros, P.G.; Diaz, L.; Kletsas, D.; Basdra, E.K.; Alexandrides, T.K.; Zadik, Z.; Frank, S.J.; Papathanassopoulou, V.; Beratis, N.G. Growth hormone/JAK-STAT axis signal-transduction defect. FEBS J. 2006, 273, 3454–3466. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, A.; Gómez-González, B. Genome instability: A mechanistic view of its causes and consequences. Nat. Rev. Genet. 2008, 9, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Lachmann, A.; Ma’ayan, A. KEA: Kinase enrichment analysis. Bioinformatics 2009, 25, 684–686. [Google Scholar] [CrossRef] [PubMed]
- Kaina, B. DNA damage-triggered apoptosis: Critical role of DNA repair, double-strand breaks, cell proliferation and signaling. Biochem. Pharmacol. 2003, 66, 1547–1554. [Google Scholar] [CrossRef]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353. [Google Scholar] [CrossRef] [PubMed]
- Bayram, F.; Bitgen, N.; Donmez-Altuntas, H.; Cakir, I.; Hamurcu, Z.; Sahin, F.; Simsek, Y.; Baskol, G. Increased genome instability and oxidative DNA damage and their association with IGF-1 levels in patients with active acromegaly. Growth Horm. IGF Res. 2014, 24, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Hamurcu, Z.; Cakir, I.; Donmez-Altuntas, H.; Bitgen, N.; Karaca, Z.; Elbuken, G.; Bayram, F. Micronucleus evaluation in mitogen-stimulated lymphocytes of patients with acromegaly. Metabolism 2011, 60, 1620–1626. [Google Scholar] [CrossRef] [PubMed]
- Lemon, J.; Rollo, C.; Boreham, D. Elevated DNA damage in a mouse model of oxidative stress: Impacts of ionizing radiation and a protective dietary supplement. Mutagenesis 2008, 23, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Pauliny, A.; Devlin, R.H.; Johnsson, J.I.; Blomqvist, D. Rapid growth accelerates telomere attrition in a transgenic fish. BMC Evol. Biol. 2015, 15, 159. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, H.; Handayaningsih, A.-E.; Iguchi, G.; Cho, Y.; Takahashi, M.; Yamamoto, M.; Suda, K.; Kasahara, K.; Hakuno, F.; Yamanouchi, K. Enhanced oxidative stress in GH-transgenic rat and acromegaly in humans. Growth Horm. IGF Res. 2012, 22, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.E. Growth Hormone Deficiency: Physiology and Clinical Management; Springer: Berlin, Germany, 2016. [Google Scholar]
- Oldfield, E.H.; Jane, J.A., Jr.; Thorner, M.O.; Pledger, C.L.; Sheehan, J.P.; Vance, M.L. Correlation between GH and IGF-1 during treatment for acromegaly. J. Neurosurg. 2016, 126, 1959–1966. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-de-Los-Monteros, A.; Sosa-Eroza, E.; Gonzalez, B.; Mendoza, V.; Mercado, M. Prevalence, clinical and biochemical spectrum and treatment outcome of acromegaly with normal basal GH at diagnosis. J. Clin. Endocrinol. Metab. 2018, 103, 3919–3924. [Google Scholar] [CrossRef] [PubMed]
- Casanueva, F.F. Growth Hormone in Health and Disease; Academic Press: Cambridge, MA, USA, 2016; Volume 138. [Google Scholar]
- Mosimann, C.; Kaufman, C.K.; Li, P.; Pugach, E.K.; Tamplin, O.J.; Zon, L.I. Ubiquitous transgene expression and Cre-based recombination driven by the ubiquitin promoter in zebrafish. Development 2011, 138, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Burket, C.T.; Montgomery, J.E.; Thummel, R.; Kassen, S.C.; LaFave, M.C.; Langenau, D.M.; Zon, L.I.; Hyde, D.R. Generation and characterization of transgenic zebrafish lines using different ubiquitous promoters. Transgenic Res. 2008, 17, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; McCormick, S.D. Prolactin and growth hormone in fish osmoregulation. Gen. Comp. Endocrinol. 2006, 147, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Vélez, E.J.; Lutfi, E.; Azizi, S.; Perelló, M.; Salmerón, C.; Riera-Codina, M.; Ibarz, A.; Fernández-Borràs, J.; Blasco, J.; Capilla, E.; et al. Understanding fish muscle growth regulation to optimize aquaculture production. Aquaculture 2017, 467, 28–40. [Google Scholar] [CrossRef]
- Reid, T.J.; Jin, Z.; Shen, W.; Reyes-Vidal, C.M.; Fernandez, J.C.; Bruce, J.N.; Kostadinov, J.; Post, K.D.; Freda, P.U. IGF-1 levels across the spectrum of normal to elevated in acromegaly: Relationship to insulin sensitivity, markers of cardiovascular risk and body composition. Pituitary 2015, 18, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Podlutsky, A.; Valcarcel-Ares, M.N.; Yancey, K.; Podlutskaya, V.; Nagykaldi, E.; Gautam, T.; Miller, R.A.; Sonntag, W.E.; Csiszar, A.; Ungvari, Z. The GH/IGF-1 axis in a critical period early in life determines cellular DNA repair capacity by altering transcriptional regulation of DNA repair-related genes: Implications for the developmental origins of cancer. Geroscience 2017, 39, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Katznelson, L.; Laws, E.R., Jr.; Melmed, S.; Molitch, M.E.; Murad, M.H.; Utz, A.; Wass, J.A. Acromegaly: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2014, 99, 3933–3951. [Google Scholar] [CrossRef] [PubMed]
- Charames, G.S.; Bapat, B. Genomic instability and cancer. Curr. Mol. Med. 2003, 3, 589–596. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinases | Substrate in Input | Substrates in Database | Combined Score |
|---|---|---|---|
| The main eight kinases regulate Upregulated genes in Acromegaly brain | |||
| HIPK2 | 9 | 43 | 32.7 |
| MAPK14 | 9 | 43 | 32.72258 |
| PRKDC | 16 | 461 | 15.36258 |
| ATR | 11 | 227 | 15.34714 |
| TAF1 | 6 | 63 | 14.71918 |
| CSNK2A1 | 5 | 42 | 13.43954 |
| CHEK2 | 17 | 521 | 13.2443 |
| CDK2 | 5 | 41 | 12.59273 |
| The main eight kinases regulate Upregulated genes in Acromegaly liver | |||
| MAPK1 | 34 | 312 | 76.13761 |
| CSNK2A1 | 38 | 521 | 57.07884 |
| GSK3B | 36 | 600 | 41.68791 |
| CHUK | 14 | 90 | 35.26104 |
| MAPK14 | 29 | 461 | 34.38588 |
| ATM | 18 | 177 | 34.03207 |
| CSNK2A2 | 22 | 267 | 33.89444 |
| AKT1 | 21 | 256 | 31.73479 |
| The main eight kinases regulate Upregulated genes in Acromegaly Kidney | |||
| MAPK1 | 17 | 312 | 36.51985 |
| CSNK2A1 | 21 | 521 | 33.22942 |
| CSNK2A2 | 15 | 267 | 31.16424 |
| MAPK8 | 14 | 275 | 26.28329 |
| ATM | 11 | 177 | 24.1775 |
| HIPK2 | 7 | 43 | 23.41801 |
| MAPK14 | 17 | 461 | 22.83626 |
| ATR | 7 | 63 | 18.45281 |
| The main eight kinases regulate Upregulated genes in Acromegaly larvae | |||
| MAPK1 | 28 | 312 | 53.17025 |
| AKT1 | 22 | 256 | 41.05688 |
| CSNK2A1 | 32 | 521 | 39.35772 |
| MAPK3 | 21 | 251 | 34.32238 |
| ATM | 17 | 177 | 30.62717 |
| HIPK2 | 10 | 43 | 28.47911 |
| ATR | 11 | 63 | 26.07581 |
| PRKCD | 14 | 146 | 24.01391 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elbialy, A.; Asakawa, S.; Watabe, S.; Kinoshita, S. A Zebrafish Acromegaly Model Elevates DNA Damage and Impairs DNA Repair Pathways. Biology 2018, 7, 47. https://doi.org/10.3390/biology7040047
Elbialy A, Asakawa S, Watabe S, Kinoshita S. A Zebrafish Acromegaly Model Elevates DNA Damage and Impairs DNA Repair Pathways. Biology. 2018; 7(4):47. https://doi.org/10.3390/biology7040047
Chicago/Turabian StyleElbialy, Abdalla, Shuichi Asakawa, Shugo Watabe, and Shigeharu Kinoshita. 2018. "A Zebrafish Acromegaly Model Elevates DNA Damage and Impairs DNA Repair Pathways" Biology 7, no. 4: 47. https://doi.org/10.3390/biology7040047