
Central Autonomic Mechanisms Involved in the Control of Laryngeal Activity and Vocalization
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
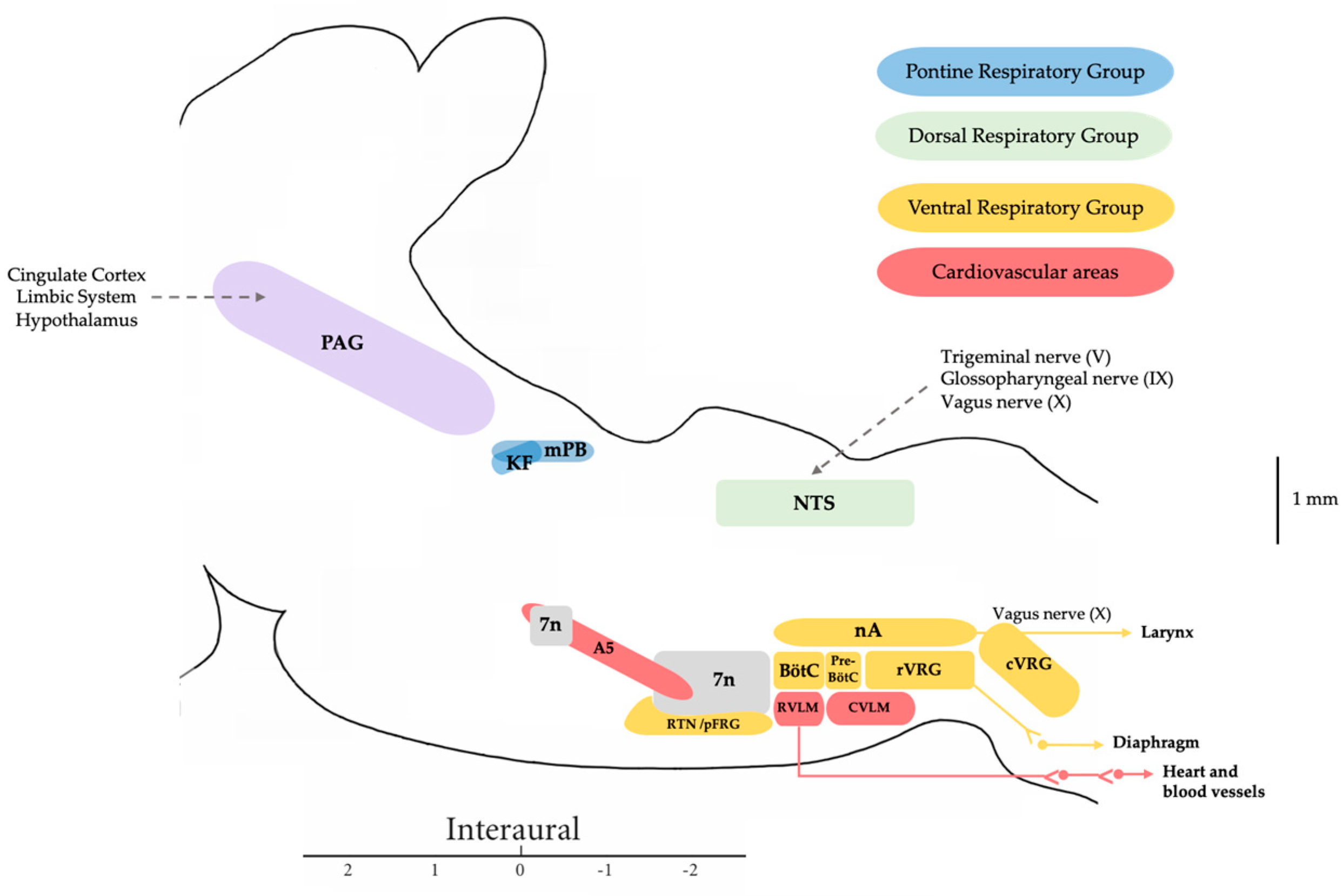
2. Central Autonomic Network Involved in the Control of Respiration and Vocal Emission
2.1. Periaqueductal Grey Matter
2.2. Pontine Nuclei
2.3. Medullary Nuclei
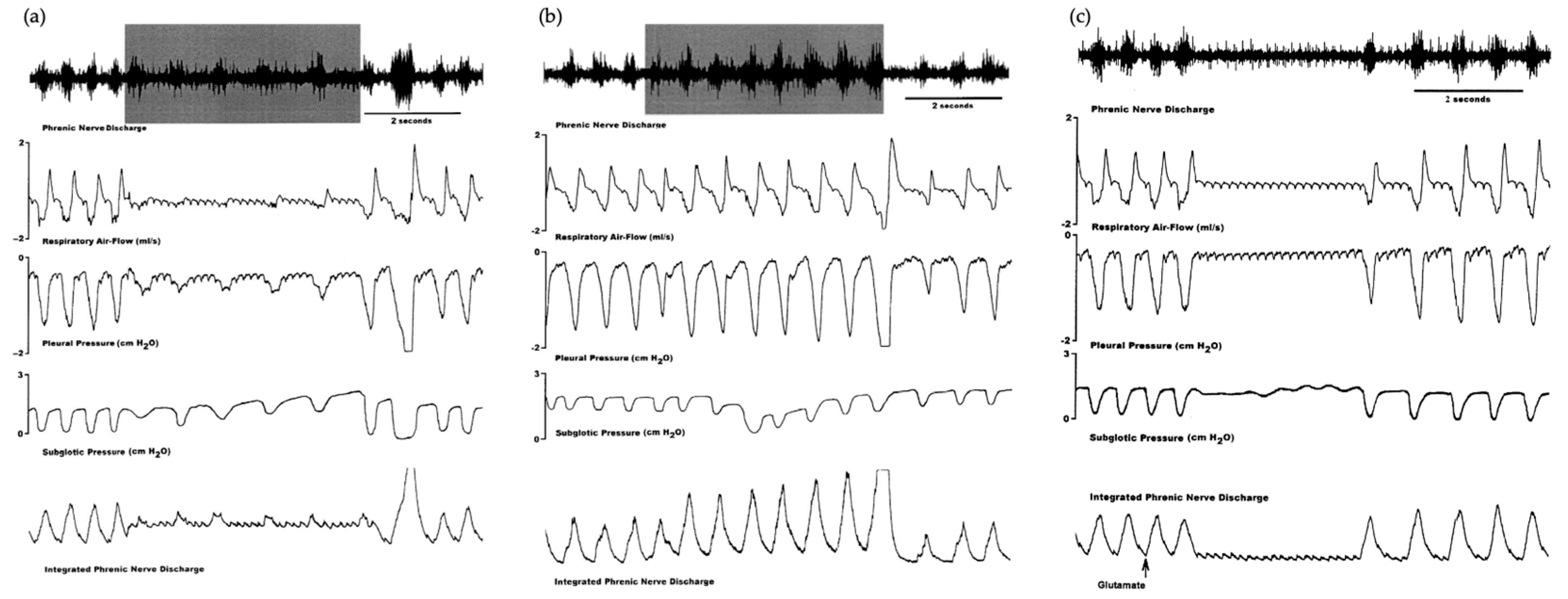
2.3.1. Pre-Bötzinger and Bötzinger Complexes
2.3.2. Nucleus Ambiguus
2.3.3. Nucleus Retroambiguus
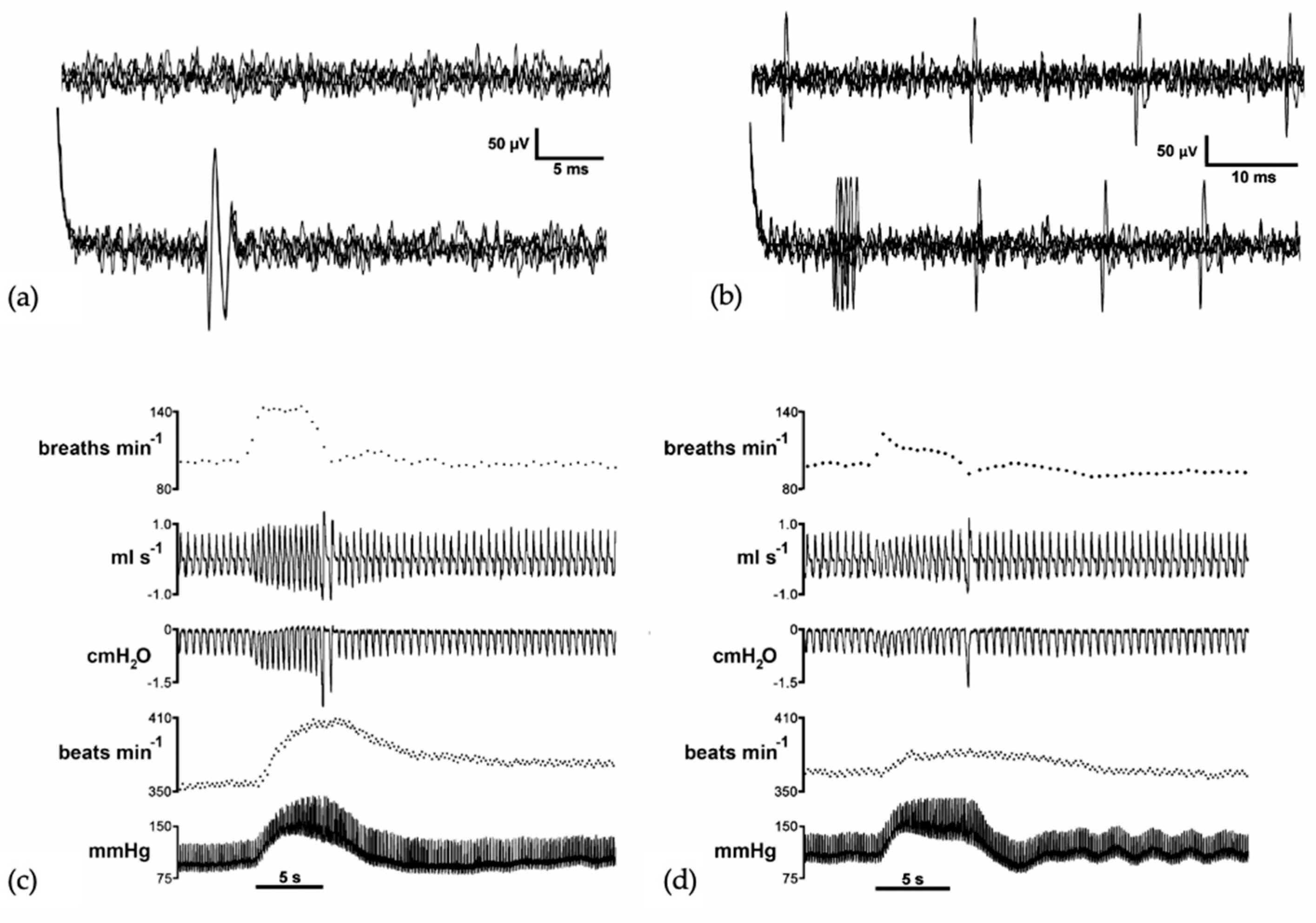
3. Vocalization in Apes: Connectivity between the PAG and the Laryngeal Motor Cortex
4. Clinical Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BötC | Bötzinger complex |
| cVRG | caudal ventral respiratory group |
| dlPAG | dorsolateral periaqueductal grey matter |
| DRG | dorsal respiratory group |
| KF | Kölliker–Fuse nucleus |
| LMC | laryngeal motor cortex |
| lPAG | lateral periaqueductal grey matter |
| lPB | lateral parabrachial |
| mPB-KF | medial parabrachial region and Kölliker–Fuse nucleus |
| mPB | medial parabrachial |
| nA | nucleus ambiguus |
| nRA | nucleus retroambiguus |
| NTS | nucleus tractus solitarius |
| PAG | periaqueductal grey matter |
| PB | parabrachial |
| PBc | parabrachial complex |
| Pre-BötC | pre-Bötzinger complex |
| PRG | pontine respiratory group |
| rVRG | rostral ventral respiratory group |
| vlPAG | ventrolateral periaqueductal grey matter |
| VRG | ventral respiratory group |
References
- Albersheim-Carter, J.; Blubaum, A.; Ballagh, I.H.; Missaghi, K.; Siuda, E.R.; McMurray, G.; Bass, A.H.; Dubuc, R.; Kelley, D.B.; Schmidt, M.F.; et al. Testing the evolutionary conservation of vocal motoneurons in vertebrates. Respir. Physiol. Neurobiol. 2016, 224, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Inagi, K.; Schultz, E.; Ford, C.N. An Anatomic Study of the Rat Larynx: Establishing the Rat Model for Neuromuscular Function. Otolaryngol. Head Neck Surg. 1998, 118, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.C.N.; Ferreira, M.V.N.; Serra, L.S.M.; Kuckelhaus, S.A.S.; da Silva, E.M.; Sampaio, A.L.L. Methodological Approaches for Vocal Folds Experiments in Laryngology: A Scoping Review. J. Voice 2022, in press. [Google Scholar] [CrossRef]
- Uludag, M.; Aygun, N.; Kartal, K.; Besler, E.; Isgor, A. Innervation of the human posterior cricoarytenoid muscle by the external branch of the superior laryngeal nerve. Head Neck 2017, 39, 2200–2207. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.; Minutello, K.; Murcek, B.W. Anatomy, Head and Neck, Larynx Recurrent Laryngeal Nerve; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Jürgens, U. The neural control of vocalization in mammals: A review. J. Voice 2009, 23, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Simonyan, K. The laryngeal motor cortex: Its organization and connectivity. Curr. Opin. Neurobiol. 2014, 28, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Tschida, K.; Michael, V.; Takatoh, J.; Han, B.X.; Zhao, S.; Sakurai, K.; Mooney, R.; Wang, F. A Specialized Neural Circuit Gates Social Vocalizations in the Mouse. Neuron 2019, 103, 459–472.e4. [Google Scholar] [CrossRef]
- Del Negro, C.A.; Funk, G.D.; Feldman, J.L. Breathing matters. Nat. Rev. Neurosci. 2018, 19, 351–367. [Google Scholar] [CrossRef]
- De Troyer, A.; Kirkwood, P.A.; Wilson, T.A. Respiratory action of the intercostal muscles. Physiol. Rev. 2005, 85, 717–756. [Google Scholar] [CrossRef]
- Torres-Tamayo, N.; García-Martínez, D.; Lois Zlolniski, S.; Torres-Sánchez, I.; García-Río, F.; Bastir, M. 3D analysis of sexual dimorphism in size, shape and breathing kinematics of human lungs. J. Anat. 2018, 232, 227–237. [Google Scholar] [CrossRef]
- Welch, J.F.; Kipp, S.; Sheel, A.W. Respiratory muscles during exercise: Mechanics, energetics, and fatigue. Curr. Opin. Physiol. 2019, 10, 102–109. [Google Scholar] [CrossRef]
- Mortola, J.P. Lung viscoelasticity: Implications on breathing and forced expiration. Clin. Pulm. Med. 2013, 20, 144–148. [Google Scholar] [CrossRef]
- Wertheimer, E. Recherches expérimentales sur les centres respiratoires de la moelle épinière. J. Anat. Physiol. Norm. Pathol. Homme Animaux 1886, 22, 458–507. [Google Scholar]
- Hollinshead, W.H.; Keswani, N.H. Localization of the phrenic nucleus in the spinal cord of man. Anat. Rec. 1956, 125, 683–699. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Capelli, P.; Bouvier, J.; Goulding, M.; Arber, S.; Fortin, G. A V0 core neuronal circuit for inspiration. Nat. Commun. 2017, 8, 544. [Google Scholar] [CrossRef]
- Dutschmann, M.; Jones, S.E.; Subramanian, H.H.; Stanic, D.; Bautista, T.G. The physiological significance of postinspiration in respiratory control. Prog. Brain Res. 2014, 212, 113–130. [Google Scholar] [CrossRef] [PubMed]
- Dhingra, R.R.; Dick, T.E.; Furuya, W.I.; Galán, R.F.; Dutschmann, M. Volumetric mapping of the functional neuroanatomy of the respiratory network in the perfused brainstem preparation of rats. J. Physiol. 2020, 598, 2061–2079. [Google Scholar] [CrossRef] [PubMed]
- Riede, T.; Thomson, S.L.; Titze, I.R.; Goller, F. The evolution of the syrinx: An acoustic theory. PLoS Biol. 2019, 17, e2006507. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, Y.; Yokota, S.; Miwakeichi, F.; Pokorski, M.; Aoyama, R.; Fukuda, K.; Yoshida, H.; Toyama, Y.; Nakamura, M.; Okada, Y. Structural and functional identification of two distinct inspiratory neuronal populations at the level of the phrenic nucleus in the rat cervical spinal cord. Brain Struct. Funct. 2019, 224, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Tokuda, I.T.; Miyachi, S.; Dunn, J.C.; Herbst, C.T.; Ishimura, K.; Kaneko, A.; Kinoshita, Y.; Koda, H.; Saers, J.P.P.; et al. Evolutionary loss of complexity in human vocal anatomy as an adaptation for speech. Science 2022, 377, 760–763. [Google Scholar] [CrossRef]
- Esposito, A.; Demeurisse, G.; Alberti, B.; Fabbro, F. Complete mutism after midbrain periaqueductal gray lesion. Neuroreport 1999, 10, 681–685. [Google Scholar] [CrossRef]
- Jürgens, U. The role of the periaqueductal grey in vocal behaviour. Behav. Brain Res. 1994, 62, 107–117. [Google Scholar] [CrossRef]
- Kittelberger, J.M.; Land, B.R.; Bass, A.H. Midbrain periaqueductal gray and vocal patterning in a teleost fish. J. Neurophysiol. 2006, 96, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Farley, G.R.; Barlow, S.M.; Netsell, R. Factors influencing neural activity in parabrachial regions during cat vocalizations. Exp. Brain Res. 1992, 89, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.L.; Jürgens, U. Effects of chemical stimulation in the periaqueductal gray on vocalization in the squirrel monkey. Brain Res. Bull. 1993, 32, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Casares, A.; López-González, M.V.; Peinado-Aragonés, C.A.; González-Barón, S.; Dawid-Milner, M.S. Parabrachial complex glutamate receptors modulate the cardiorespiratory response evoked from hypothalamic defense area. Auton. Neurosci. Basic Clin. 2012, 169, 124–134. [Google Scholar] [CrossRef] [PubMed]
- López-González, M.V.; Díaz-Casares, A.; González-García, M.; Peinado-Aragonés, C.A.; Barbancho, M.A.; Carrillo de Albornoz, M.; Dawid-Milner, M.S. Glutamate receptors of the A5 region modulate cardiovascular responses evoked from the dorsomedial hypothalamic nucleus and perifornical area. J. Physiol. Biochem. 2018, 74, 325–334. [Google Scholar] [CrossRef] [PubMed]
- González-García, M.; Carrillo-Franco, L.; Peinado-Aragonés, C.A.; Díaz-Casares, A.; Gago, B.; López-González, M.V.; Dawid-Milner, M.S. Impact of the glutamatergic neurotransmission within the A5 region on the cardiorespiratory response evoked from the midbrain dlPAG. Pflügers Arch. Eur. J. Physiol. 2023, 475, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Casares, A.; López-González, M.V.; Peinado-Aragonés, C.A.; Lara, J.P.; González-Barón, S.; Dawid-Milner, M.S. Role of the parabrachial complex in the cardiorespiratory response evoked from hypothalamic defense area stimulation in the anesthetized rat. Brain Res. 2009, 1279, 58–70. [Google Scholar] [CrossRef] [PubMed]
- López-González, M.V.; Díaz-Casares, A.; Peinado-Aragonés, C.A.; Lara, J.P.; Barbancho, M.A.; Dawid-Milner, M.S. Neurons of the A5 region are required for the tachycardia evoked by electrical stimulation of the hypothalamic defence area in anaesthetized rats. Exp. Physiol. 2013, 98, 1279–1294. [Google Scholar] [CrossRef]
- López-González, M.V.; González-García, M.; Peinado-Aragonés, C.A.; Barbancho, M.Á.; Díaz-Casares, A.; Dawid-Milner, M.S. Pontine A5 region modulation of the cardiorespiratory response evoked from the midbrain dorsolateral periaqueductal grey. J. Physiol. Biochem. 2020, 76, 561–572. [Google Scholar] [CrossRef]
- Lara, J.P.; Dawid-Milner, M.S.; López, M.V.; Montes, C.; Spyer, K.M.; González-Barón, S. Laryngeal effects of stimulation of rostral and ventral pons in the anaesthetized rat. Brain Res. 2002, 934, 97–106. [Google Scholar] [CrossRef]
- Dutschmann, M.; Dick, T.E. Pontine mechanisms of respiratory control. Compr. Physiol. 2012, 2, 2443–2469. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.F.; Feldman, J.L. Efferent projections of excitatory and inhibitory preBötzinger Complex neurons. J. Comp. Neurol. 2018, 526, 1389–1402. [Google Scholar] [CrossRef]
- Anderson, T.M.; Garcia, A.J., 3rd; Baertsch, N.A.; Pollak, J.; Bloom, J.C.; Wei, A.D.; Rai, K.G.; Ramirez, J.M. A novel excitatory network for the control of breathing. Nature 2016, 536, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Dampney, R.A. Central neural control of the cardiovascular system: Current perspectives. Adv. Physiol. Educ. 2016, 40, 283–296. [Google Scholar] [CrossRef]
- Carrive, P. The periaqueductal gray and defensive behavior: Functional representation and neuronal organization. Behav. Brain Res. 1993, 58, 27–47. [Google Scholar] [CrossRef]
- Zhang, W.; Hayward, L.F.; Davenport, P.W. Respiratory responses elicited by rostral versus caudal dorsal periaqueductal gray stimulation in rats. Auton. Neurosci. 2007, 134, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, H.H. Descending control of the respiratory neuronal network by the midbrain periaqueductal grey in the rat in vivo. J. Physiol. 2013, 591, 109–122. [Google Scholar] [CrossRef]
- Faull, O.K.; Subramanian, H.H.; Ezra, M.; Pattinson, K.T.S. The midbrain periaqueductal gray as an integrative and interoceptive neural structure for breathing. Neurosci. Biobehav. Rev. 2019, 98, 135–144. [Google Scholar] [CrossRef]
- Linnman, C.; Moulton, E.A.; Barmettler, G.; Becerra, L.; Borsook, D. Neuroimaging of the periaqueductal gray: State of the field. Neuroimage 2012, 60, 505–522. [Google Scholar] [CrossRef]
- Tavares, I.; Lima, D. From neuroanatomy to gene therapy: Searching for new ways to manipulate the supraspinal endogenous pain modulatory system. J. Anat. 2007, 211, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Trevizan-Baú, P.; Dhingra, R.R.; Furuya, W.I.; Stanić, D.; Mazzone, S.B.; Dutschmann, M. Forebrain projection neurons target functionally diverse respiratory control areas in the midbrain, pons, and medulla oblongata. J. Comp. Neurol. 2021, 529, 2243–2264. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.; Jiang, J.; Saito, K.; Toth, B.A.; Dickey, J.E.; Rodeghiero, S.R.; Deng, Y.; Deng, G.; Xue, B.; Zhu, Z.; et al. Neuroanatomical organization and functional roles of PVN MC4R pathways in physiological and behavioral regulations. Mol. Metab. 2022, 55, 101401. [Google Scholar] [CrossRef] [PubMed]
- Keay, K.A.; Feil, K.; Gordon, B.D.; Herbert, H.; Bandler, R. Spinal afferents to functionally distinct periaqueductal gray columns in the rat: An anterograde and retrograde tracing study. J. Comp. Neurol. 1997, 385, 207–229. [Google Scholar] [CrossRef]
- Kawai, Y. Differential Ascending Projections from the Male Rat Caudal Nucleus of the Tractus Solitarius: An Interface between Local Microcircuits and Global Macrocircuits. Front. Neuroanat. 2018, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.E.; Yang, T.Z. The efferent projections from the reticular formation and the locus coeruleus studied by anterograde and retrograde axonal transport in the rat. Comp. Neurol. 1985, 242, 56–92. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, R.; Corsetti, G.; Rodella, L.; Tredici, G.; Gioia, M. Supraspinal connections and termination patterns of the parabrachial complex determined by the biocytin anterograde tract-tracing technique in the rat. J. Anat. 1998, 193, 417–430. [Google Scholar] [CrossRef] [PubMed]
- McGovern, A.E.; Driessen, A.K.; Simmons, D.G.; Powell, J.; Davis-Poynter, N.; Farrell, M.J.; Mazzone, S.B. Distinct brainstem and forebrain circuits receiving tracheal sensory neuron inputs revealed using a novel conditional anterograde transsynaptic viral tracing system. J. Neurosci. 2015, 35, 7041–7055. [Google Scholar] [CrossRef]
- Frontera, J.L.; Baba Aissa, H.; Sala, R.W.; Mailhes-Hamon, C.; Georgescu, I.A.; Léna, C.; Popa, D. Bidirectional control of fear memories by cerebellar neurons projecting to the ventrolateral periaqueductal grey. Nat Commun. 2020, 11, 5207. [Google Scholar] [CrossRef]
- Subramanian, H.H.; Balnave, R.J.; Holstege, G. Microstimulation in Different Parts of the Periaqueductal Gray Generates Different Types of Vocalizations in the Cat. J. Voice 2021, 35, 804.e9–804.e25. [Google Scholar] [CrossRef]
- Hartmann, K.; Brecht, M. A Functionally and Anatomically Bipartite Vocal Pattern Generator in the Rat Brain Stem. iScience 2020, 23, 101804. [Google Scholar] [CrossRef]
- Ludlow, C.L. Central nervous system control of interactions between vocalization and respiration in mammals. Head Neck 2011, 33, 121–125. [Google Scholar] [CrossRef]
- Subramanian, H.H.; Huang, Z.G.; Silburn, P.A.; Balnave, R.J.; Holstege, G. The physiological motor patterns produced by neurons in the nucleus retroambiguus in the rat and their modulation by vagal, peripheral chemosensory, and nociceptive stimulation. J. Comp. Neurol. 2018, 526, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Dutschmann, M.; Bautista, T.G.; Trevizan-Baú, P.; Dhingra, R.R.; Furuya, W.I. The pontine Kölliker-Fuse nucleus gates facial, hypoglossal, and vagal upper airway related motor activity. Respir. Physiol. Neurobiol. 2021, 284, 103563. [Google Scholar] [CrossRef]
- Ezure, K.; Tanaka, I. Distribution and medullary projection of respiratory neurons in the dorsolateral pons of the rat. Neuroscience 2006, 141, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Geerling, J.C.; Yokota, S.; Rukhadze, I.; Roe, D.; Chamberlin, N.L. Kölliker-Fuse GABAergic and glutamatergic neurons project to distinct targets. J. Comp. Neurol. 2017, 525, 1844–1860. [Google Scholar] [CrossRef]
- Núñez-Abades, P.A.; Portillo, F.; Pásaro, R. Characterisation of afferent projections to the nucleus ambiguus of the rat by means of fluorescent double labelling. J. Anat. 1990, 172, 1–15. [Google Scholar] [PubMed]
- Song, G.; Wang, H.; Xu, H.; Poon, C.S. Kölliker–Fuse neurons send collateral projections to multiple hypoxia-activated and nonactivated structures in rat brainstem and spinal cord. Brain Struct. Funct. 2012, 217, 835–858. [Google Scholar] [CrossRef]
- Dutschmann, M.; Herbert, H. The Kölliker-Fuse nucleus gates the postinspiratory phase of the respiratory cycle to control inspiratory off-switch and upper airway resistance in rat. Eur. J. Neurosci. 2006, 24, 1071–1084. [Google Scholar] [CrossRef]
- Yang, C.F.; Kim, E.J.; Callaway, E.M.; Feldman, J.L. Monosynaptic Projections to Excitatory and Inhibitory preBötzinger Complex Neurons. Front. Neuroanat. 2020, 14, 58. [Google Scholar] [CrossRef]
- Varga, A.G.; Maletz, S.N.; Bateman, J.T.; Reid, B.T.; Levitt, E.S. Neurochemistry of the Kölliker-fuse nucleus from a respiratory perspective. J. Neurochem. 2021, 156, 16–37. [Google Scholar] [CrossRef]
- Hirsch, J.A.; Bishop, B. Respiratory sinus arrhythmia in humans: How breathing pattern modulates heart rate. Am. J. Physiol. 1981, 241, 620–629. [Google Scholar] [CrossRef]
- Elstad, M.; O’Callaghan, E.L.; Smith, A.J.; Ben-Tal, A.; Ramchandra, R. Cardiorespiratory interactions in humans and animals: Rhythms for life. Am. J. Physiol. 2018, 315, 6–17. [Google Scholar] [CrossRef]
- Farmer, D.G.S.; Dutschmann, M.; Paton, J.F.R.; Pickering, A.E.; McAllen, R.M. Brainstem sources of cardiac vagal tone and respiratory sinus arrhythmia. J. Physiol. 2016, 594, 7249–7265. [Google Scholar] [CrossRef] [PubMed]
- McAllen, R.M.; Spyer, K.M. The location of cardiac vagal preganglionic motoneurones in the medulla of the cat. J. Physiol. 1976, 258, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Dergacheva, O.; Griffioen, K.J.; Neff, R.A.; Mendelowitz, D. Respiratory modulation of premotor cardiac vagal neurons in the brainstem. Respir. Physiol. Neurobiol. 2010, 174, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Navarrete-Opazo, A.A.; Cook-Snyder, D.R.; Miller, J.R.; Callison, J.J.; McCarthy, N.; Palkovic, B.; Stuth, E.A.E.; Zuperku, E.J.; Stucke, A.G. Endogenous glutamatergic inputs to the parabrachial nucleus/kölliker-fuse complex determine respiratory rate. Respir. Physiol. Neurobiol. 2020, 277, 103401. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Yu, Y.; Poon, C.S. Cytoarchitecture of pneumotaxic integration of respiratory and nonrespiratory information in the rat. J. Neurosci. 2006, 26, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Zuperku, E.J.; Stucke, A.G.; Hopp, F.A.; Stuth, E.A.E. Characteristics of breathing rate control mediated by a subregion within the pontine parabrachial complex. J. Neurophysiol. 2017, 117, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Lüthe, L.; Häusler, U.; Jürgens, U. Neuronal activity in the medulla oblongata during vocalization. A single-unit recording study in the squirrel monkey. Behav. Brain Res. 2000, 116, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Bautista, T.G.; Pitts, T.E.; Pilowsky, P.M.; Morris, K.F. The Brainstem Respiratory Network. In Neuronal Networks in Brain Function, CNS Disorders, and Therapeutics; Academic Press: Cambridge, MA, USA, 2014; pp. 235–245. ISBN 9780124158047. [Google Scholar] [CrossRef]
- Feher, J. Control of ventilation. In Quantitative Human Physiology an Introduction; Academic Press: Cambridge, MA, USA, 2017; pp. 672–681. ISBN 9780128008836. [Google Scholar] [CrossRef]
- Marchenko, V.; Koizumi, H.; Mosher, B.; Koshiya, N.; Tariq, M.F.; Bezdudnaya, T.G.; Zhang, R.; Molkov, Y.I.; Rybak, I.A.; Smith, J.C. Perturbations of respiratory rhythm and pattern by disrupting synaptic inhibition within pre-Bötzinger and Bötzinger complexes. ENeuro 2016, 3, e0011-16.2016. [Google Scholar] [CrossRef] [PubMed]
- Holstege, G. Descending motor pathways and the spinal motor system: Limbic and non-limbic components. Prog. Brain Res. 1991, 87, 307–421. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.C.; Abdala, A.P.; Koizumi, H.; Rybak, I.A.; Paton, J.F. Spatial and functional architecture of the mammalian brain stem respiratory network: A hierarchy of three oscillatory mechanisms. J. Neurophysiol. 2007, 98, 3370–3387. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.F.; Peever, J.H.; Duffin, J. Bötzinger-complex expiratory neurons monosynaptically inhibit phrenic motoneurons in the decerebrate rat. Exp. Brain Res. 1998, 122, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Ausborn, J.; Koizumi, H.; Barnett, W.H.; John, T.T.; Zhang, R.; Molkov, Y.I.; Smith, J.C.; Rybak, I.A. Organization of the core respiratory network: Insights from optogenetic and modeling studies. PLOS Comput. Biol. 2018, 14, e1006148. [Google Scholar] [CrossRef]
- Richter, D.W.; Smith, J.C. Respiratory rhythm generation in vivo. Physiology 2014, 29, 58–71. [Google Scholar] [CrossRef]
- Pascual-Font, A.; Hernández-Morato, I.; McHanwell, S.; Vázquez, T.; Maranillo, E.; Sañudo, J.; Valderrama-Canales, F.J. The central projections of the laryngeal nerves in the rat. J. Anat. 2011, 219, 217–228. [Google Scholar] [CrossRef]
- Hernández-Morato, I.; Pascual-Font, A.; Ramírez, C.; Matarranz-Echeverría, J.; McHanwell, S.; Vázquez, T.; Sañudo, J.R.; Valderrama-Canales, F.J. Somatotopy of the neurons innervating the cricothyroid, posterior cricoarytenoid, and thyroarytenoid muscles of the rat’s larynx. Anat. Rec. 2013, 296, 470–479. [Google Scholar] [CrossRef]
- Hernandez-Morato, I.; Pitman, M.J.; Sharma, S. Muscle specific nucleus ambiguus neurons isolation and culturing. J. Neurosci. Methods 2016, 273, 33–39. [Google Scholar] [CrossRef]
- Merrill, E.G. The lateral respiratory neurones of the medulla: Their associations with nucleus ambiguus, nucleus retroambigualis, the spinal accessory nucleus and the spinal cord. Brain Res. 1970, 24, 11–28. [Google Scholar] [CrossRef]
- Subramanian, H.H.; Holstege, G. The nucleus retroambiguus control of respiration. J. Neurosci. 2009, 29, 3824–3832. [Google Scholar] [CrossRef]
- Krohn, F.; Novello, M.; van der Giessen, R.S.; De Zeeuw, C.I.; Pel, J.J.M.; Bosman, L.W.J. The integrated brain network that controls respiration. Elife 2023, 12, e83654. [Google Scholar] [CrossRef]
- Buttry, J.L.; Goshgarian, H.G. WGA-Alexa transsynaptic labeling in the phrenic motor system of adult rats: Intrapleural injection versus intradiaphragmatic injection. J. Neurosci. Methods 2015, 241, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Jensen, V.N.; Alilain, W.J.; Crone, S.A. Role of Propriospinal Neurons in Control of Respiratory Muscles and Recovery of Breathing Following Injury. Front. Syst. Neurosci. 2020, 13, 84. [Google Scholar] [CrossRef]
- Zheng, Y.; Riche, D.; Rekling, J.C.; Foutz, A.S.; Denavit-Saubié, M. Brainstem neurons projecting to the rostral ventral respiratory group in the medulla oblongata of the rat revealed by co-application of NMDA and biocytin. Brain Res. 1998, 782, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Boers, J.; Kirkwood, P.A.; de Weerd, H.; Holstege, G. Ultrastructural evidence for direct excitatory retroambiguus projections to cutaneous trunci and abdominal external oblique muscle motoneurons in the cat. Brain Res. Bull. 2006, 68, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Holstege, G.; Subramanian, H.H. Two different motor systems are needed to generate human speech. J. Comp. Neurol. 2016, 524, 1558–1577. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Stanić, D.; Dutschmann, M. Dorsal and ventral aspects of the most caudal medullary reticular formation have differential roles in modulation and formation of the respiratory motor pattern in rat. Brain Struct. Funct. 2016, 221, 4353–4368. [Google Scholar] [CrossRef]
- Rosin, D.L.; Chang, D.A.; Guyenet, P.G. Afferent and efferent connections of the rat retrotrapezoid nucleus. J. Comp. Neurol. 2006, 499, 64–89. [Google Scholar] [CrossRef]
- Gerrits, P.O.; Holstege, G. Pontine and medullary projections to the nucleus retroambiguus: A wheat germ agglutinin-horseradish peroxidase and autoradiographic tracing study in the cat. J. Comp. Neurol. 1996, 373, 173–185. [Google Scholar] [CrossRef]
- Holstege, G. The somatic motor system. Prog. Brain Res. 1996, 107, 9–26. [Google Scholar] [CrossRef]
- Holstege, G. The emotional motor system. Eur. J. Morphol. 1992, 30, 67–79. [Google Scholar]
- Jürgens, U.; Hage, S.R. On the role of the reticular formation in vocal pattern generation. Behav. Brain Res. 2007, 182, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Ngan, E.; Liotti, M. A larynx area in the human motor cortex. Cereb. Cortex 2008, 18, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Ludlow, C.L. Central nervous system control of the laryngeal muscles in humans. Respir. Physiol. Neurobiol. 2005, 147, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Loucks, T.M.; Poletto, C.J.; Simonyan, K.; Reynolds, C.L.; Ludlow, C.L. Human brain activation during phonation and exhalation: Common volitional control for two upper airway functions. Neuroimage 2007, 36, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Jürgens, U.; Pratt, R. Role of the periaqueductal grey in vocal expression of emotion. Brain Res. 1979, 167, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Kirzinger, A.; Jürgens, U. Cortical lesion effects and vocalization in the squirrel monkey. Brain Res. 1982, 233, 299–315. [Google Scholar] [CrossRef] [PubMed]
- Schadt, C.R.; Cox, K.L.; Tramontana, M.G.; Byrne, D.W.; Davis, T.L.; Fang, J.Y.; Konrad, P.E.; Padaliya, B.; Mutter, R.W.; Gill, C.E.; et al. Depression and intelligence in patients with Parkinson’s disease and deep-brain stimulation. J. Natl. Med. Assoc. 2006, 98, 1121–1125. [Google Scholar] [PubMed]
- Peterson, J.R.; Watts, C.R.; Morris, J.A.; Shelton, J.M.; Cooper, B.G. Laryngeal aging and acoustic changes in male rat ultrasonic vocalizations. Dev. Psychobiol. 2013, 55, 818–828. [Google Scholar] [CrossRef]
- Drager, L.F.; Genta, P.R.; Pedrosa, R.P.; Nerbass, F.B.; Gonzaga, C.C.; Krieger, E.M.; Lorenzi-Filho, G. Characteristics and predictors of obstructive sleep apnea in patients with systemic hypertension. Am. J. Cardiol. 2010, 105, 1135–1139. [Google Scholar] [CrossRef]
- Wang, X.; Guo, R.; Zhao, W.; Pilowsky, P.M. Medullary mediation of the laryngeal adductor reflex: A possible role in sudden infant death syndrome. Respir. Physiol. Neurobiol. 2016, 226, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Mor, N.; Simonyan, K.; Blitzer, A. Central voice production and pathophysiology of spasmodic dysphonia. Laryngoscope 2018, 128, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Christopher, K.L.; Morris, M.J. Vocal cord dysfunction, paradoxic vocal fold motion, or laryngomalacia? Our understanding requires an interdisciplinary approach. Otolaryngol. Clin. N. Am. 2010, 43, 43–66. [Google Scholar] [CrossRef] [PubMed]
- Van Houtte, E.; Van Lierde, K.; Claeys, S. Pathophysiology and treatment of muscle tension dysphonia: A review of the current knowledge. J. Voice 2011, 25, 202–207. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-García, M.; Carrillo-Franco, L.; Morales-Luque, C.; Dawid-Milner, M.S.; López-González, M.V. Central Autonomic Mechanisms Involved in the Control of Laryngeal Activity and Vocalization. Biology 2024, 13, 118. https://doi.org/10.3390/biology13020118
González-García M, Carrillo-Franco L, Morales-Luque C, Dawid-Milner MS, López-González MV. Central Autonomic Mechanisms Involved in the Control of Laryngeal Activity and Vocalization. Biology. 2024; 13(2):118. https://doi.org/10.3390/biology13020118
Chicago/Turabian StyleGonzález-García, Marta, Laura Carrillo-Franco, Carmen Morales-Luque, Marc Stefan Dawid-Milner, and Manuel Víctor López-González. 2024. "Central Autonomic Mechanisms Involved in the Control of Laryngeal Activity and Vocalization" Biology 13, no. 2: 118. https://doi.org/10.3390/biology13020118