
Small Molecules Targeting INSM1 for the Treatment of High-Risk Neuroblastoma
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Characteristics of NB
1.2. Identification of Biomarker Critical for Aggressive NB
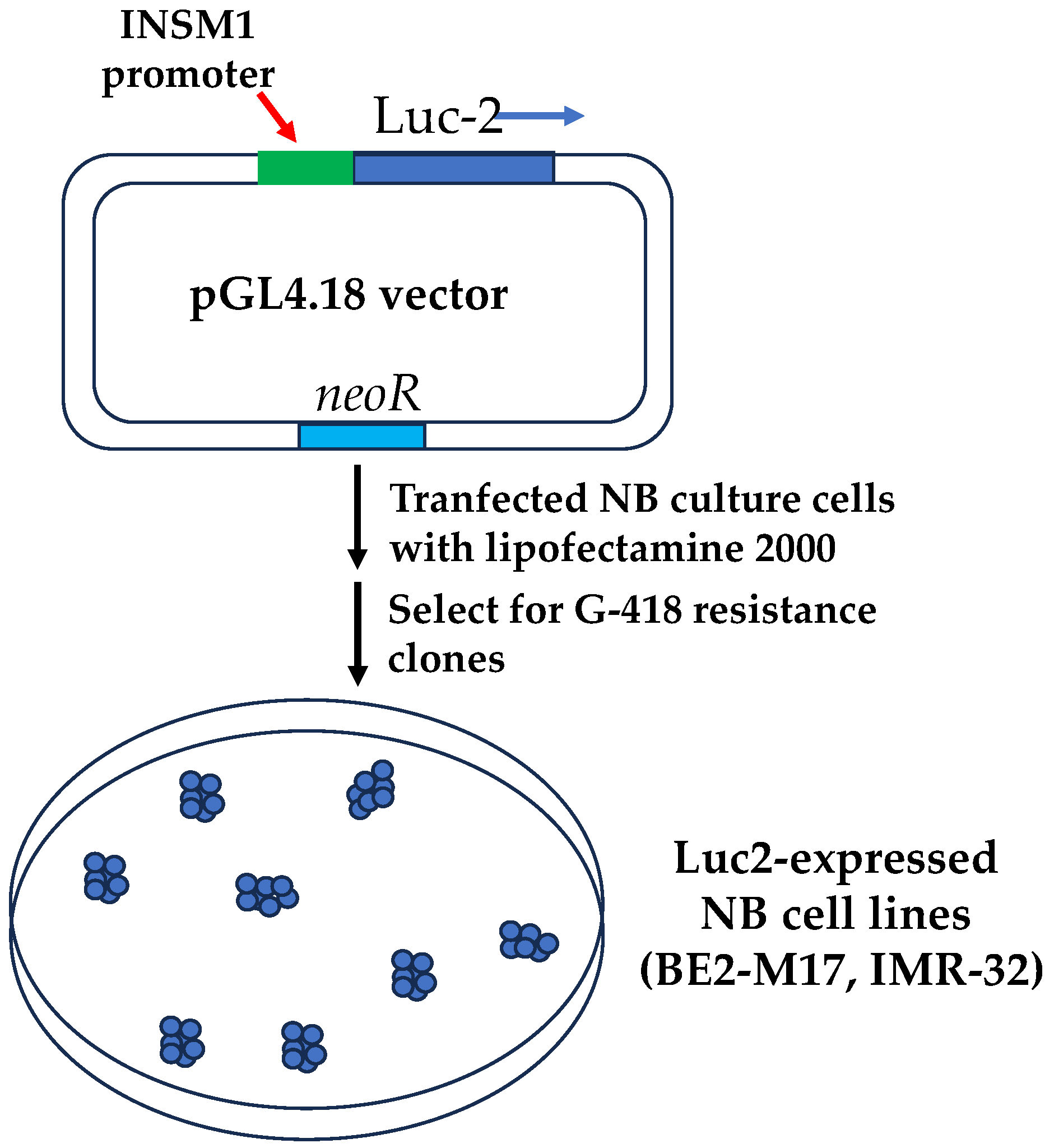
1.3. Development of a Bioassay Targeting INSM1 in NB
1.4. Small Molecules Target INSM1 Expression and NB Tumor Growth
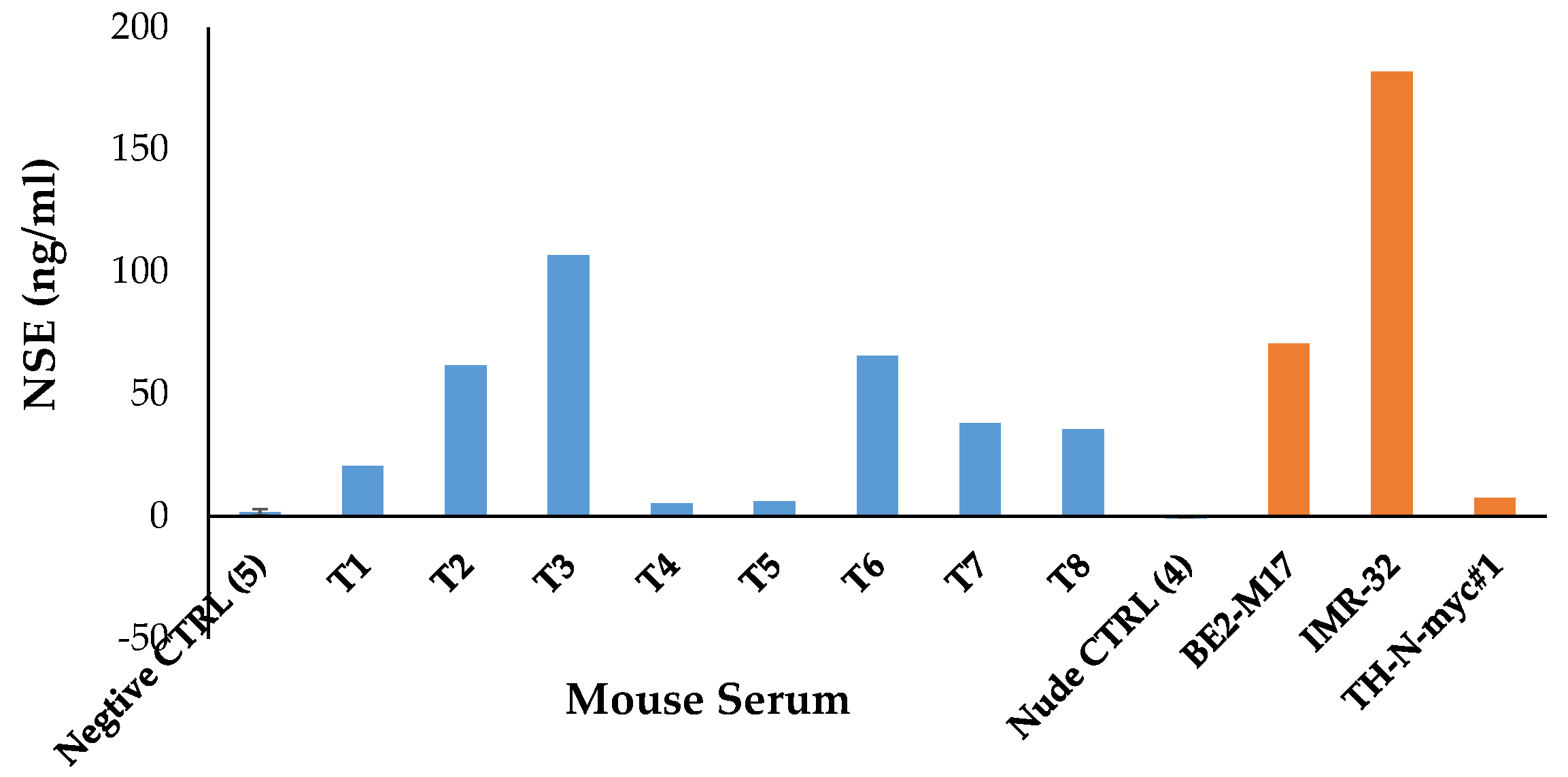
1.5. Animal Model for NB Therapy
2. Discussion
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harris, G.W. Development and present status of neuroendocrinology. Dtsch. Med. Wochenschr. 1965, 90, 61–65. [Google Scholar] [CrossRef]
- Heitz, P.U.; Roth, J.; Zuber, C.; Komminoth, P. Markers for Neural and Endocrine Cells in Pathology; VCH Publishers, Inc.: New York, NY, USA, 1991. [Google Scholar]
- Goto, Y.; DeSilva, M.G.; Toscani, A.; Prabhakar, B.S.; Notkins, A.L.; Lan, M.S. A novel human insulinoma-associated cDNA, IA-1, encodes a protein with zinc-finger DNA-binding motifs. J. Biol. Chem. 1992, 267, 15252–15257. [Google Scholar] [CrossRef]
- Breslin, M.B.; Zhu, M.; Lan, M.S. NeuroD1/E47 regulates the E-box element of a novel zinc-finger transcription factor, IA-1, in developing nervous system. J. Biol. Chem. 2003, 278, 38991–38997. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, J.N.; Guo, Z.; Baus, R.M.; Werner, H.; Rehrauer, W.M.; Lloyd, R.V. INSM1: A Novel Immunohistochemical and Molecular Marker for Neuroendocrine and Neuroepithelial Neoplasms. Am. J. Clin. Pathol. 2015, 144, 579–591. [Google Scholar] [CrossRef] [Green Version]
- Rooper, L.M.; Sharma, R.; Li, Q.K.; Illei, P.; Westra, W.H. INSM1 Demonstrates Superior Performance to the individual and combined use of Synaptophysin, Chromogranin, and CD56 for Diagnosing Neuroendocrine Tumors of the Thoracic Cavity. Am. J. Surg. Pathol. 2017, 41, 1561–1569. [Google Scholar] [CrossRef]
- Tanigawa, M.; Nakayama, M.; Taira, T.; Hattori, S.; Mihara, Y.; Kondo, R.; Kusano, H.; Nakamura, K.; Abe, Y.; Ishida, Y.; et al. Insulinoma-associated protein 1 (INSM1) is a useful marker for pancreatic neuroendocrine tumor. Med. Mol. Morphol. 2018, 51, 32–40. [Google Scholar] [CrossRef]
- Wildner, H.; Gierl, M.S.; Strehle, M.; Pla, P.; Birchmeier, C. Insm1 (IA-1) is a crucial component of the transcriptional network that controls differentiation of the sympatho-adrenal lineage. Development 2008, 135, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Wang, G.; Yu, S.; Sun, J.; He, L.; Zhao, H.; Ma, Y.; Wang, F.; Wang, X.; Wang, R.; et al. Transcriptomic analysis identifies a tumor subtype mRNA classifier for invasive non-functioning pituitary neuroendocrine tumor diagnostics. Theranostics 2021, 11, 132–146. [Google Scholar] [CrossRef]
- Doxtader, E.E.; Mukhopadhyay, S. Insulinoma-associated protein 1 is a sensitive and specific marker of neuroendocrine lung neoplasms in cytology specimens. Cancer Cytopathol. 2018, 126, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, M.; Ludtke, T.H.; Deuper, L.; Rudat, C.; Christoffels, V.M.; Kispert, A.; Trowe, M.O. TBX2 specifies and maintains inner hair and supporting cell fate in the Organ of Corti. Nat. Commun. 2022, 13, 7628. [Google Scholar] [CrossRef]
- Rosenbaum, J.N.; Duggan, A.; Garcia-Anoveros, J. Insm1 promotes the transition of olfactory progenitors from apical and proliferative to basal, terminally dividing and neuronogenic. Neural Dev. 2011, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Maris, J.M.; Hogarty, M.D.; Bagatell, R.; Cohn, S.L. Neuroblastoma. Lancet 2007, 369, 2106–2120. [Google Scholar] [CrossRef]
- Bresler, S.C.; Weiser, D.A.; Huwe, P.J.; Park, J.H.; Krytska, K.; Ryles, H.; Laudenslager, M.; Rappaport, E.F.; Wood, A.C.; McGrady, P.W.; et al. ALK mutations confer differential oncogenic activation and sensitivity to ALK inhibition therapy in neuroblastoma. Cancer Cell 2014, 26, 682–694. [Google Scholar] [CrossRef] [Green Version]
- van Limpt, V.; Schramm, A.; van Lakeman, A.; Sluis, P.; Chan, A.; van Noesel, M.; Baas, F.; Caron, H.; Eggert, A.; Versteeg, R. The Phox2B homeobox gene is mutated in sporadic neuroblastomas. Oncogene 2004, 23, 9280–9288. [Google Scholar] [CrossRef] [Green Version]
- Mosse, Y.; Greshock, J.; King, A.; Khazi, D.; Weber, B.L.; Maris, J.M. Identification and high-resolution mapping of a constitutional 11q deletion in an infant with multifocal neuroblastoma. Lancet Oncol. 2003, 4, 769–771. [Google Scholar] [CrossRef]
- Heck, J.E.; Ritz, B.; Hung, R.J.; Hashibe, M.; Boffetta, P. The epidemiology of neuroblastoma: A review. Paediatr. Perinat. Epidemiol. 2009, 23, 125–143. [Google Scholar] [CrossRef]
- Zhou, Q.Y.; Quaife, C.J.; Palmiter, R.D. Targeted disruption of the tyrosine hydroxylase gene reveals that catecholamines are required for mouse fetal development. Nature 1995, 374, 640–643. [Google Scholar] [CrossRef]
- Thompson, S.A.; Matsumoto, A.M.; Palmiter, R.D. Noradrenaline is essential for mouse fetal development. Nature 1995, 374, 643–646. [Google Scholar]
- Chen, C.; Breslin, M.B.; Guidry, J.J.; Lan, M.S. 5′-Iodotubercidin represses insulinoma-associated-1 expression, decreases cAMP levels, and suppresses human neuroblastoma cell growth. J. Biol. Chem. 2019, 294, 5456–5465. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Wu, J.; Hicks, C.; Lan, M.S. Repurposing a plant alkaloid homoharringtonine targets insulinoma associated-1 in N-Myc-activated neuroblastoma. Cell. Signal. 2023, 109, 110753. [Google Scholar] [CrossRef]
- Karanika, E.; Soupsana, K.; Christogianni, A.; Stellas, D.; Klinakis, A.; Politou, A.S.; Georgatos, S. Haspin-dependent and independent effects of the kinase inhibitor 5-Iodotubercidin on self-renewal and differentiation. Sci. Rep. 2020, 10, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewald, B.; Sampath, D.; Plunkett, W. Nucleoside analogs: Molecular mechanisms signaling cell death. Oncogene 2008, 27, 6522–6537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Jia, D.; Liu, H.; Zhu, N.; Zhang, W.; Feng, J.; Yin, J.; Hao, B.; Cui, D.; Deng, Y.; et al. Identification of 5-Iodotubercidin as a genotoxic drug with anti-cancer potential. PLoS ONE 2013, 8, e62527. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Bode, A.M.; Dong, Z.; Lee, M.H. AKT as a Therapeutic Target for Cancer. Cancer Res. 2019, 79, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Avendano, C.; Menendez, J.C. Anticancer Drugs Acting via Radical Species: Radiotherapy and Photodynamic Therapy of Cancer; Carmen Avendono, J.C.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2015; Chapter 4; pp. 133–195. [Google Scholar]
- Holstege, C.P. Mithramycin; Wexler, P., Ed.; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Minotti, G.; Menna, P.; Salvatorelli, E.; Cairo, G.; Gianni, L. Anthracyclines: Molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol. Rev. 2004, 56, 185–229. [Google Scholar] [CrossRef] [Green Version]
- Dozic, S.; Howden, E.J.; Bell, J.R.; Mellor, K.M.; Delbridge, L.M.D.; Weeks, K.L. Cellular Mechanisms Mediating Exercise-Induced Protection against Cardiotoxic Anthracycline Cancer Therapy. Cells 2023, 12, 1312. [Google Scholar] [CrossRef]
- Thomas, X.; Archimbaud, E. Mitoxantrone in the treatment of acute myelogenous leukemia: A review. Hematol. Cell Ther. 1997, 39, 63–74. [Google Scholar] [CrossRef]
- Preethi, S.; Kumar, H.; Rawal, V.B.; Ajmeer, R.; Jain, V. Overview of mitoxantrone-a potential candidate for treatment of breast cancer. Int. J. Appl. Pharm. 2022, 14, 10–22. [Google Scholar]
- Ueda, H.; Nakajima, H.; Hori, Y.; Goto, T.; Okuhara, M. Action of FR901228, a novel antitumor bicyclic depsipeptide produced by Chromobacterium violaceum no. 968, on Ha-ras transformed NIH3T3 cells. Biosci. Biotechnol. Biochem. 1994, 58, 1579–1583. [Google Scholar] [CrossRef]
- Ellis, L.; Pan, Y.; Smyth, G.K.; George, D.J.; McCormack, C.; Williams-Truax, R.; Mita, M.; Beck, J.; Burris, H.; Ryan, G.; et al. Histone deacetylase inhibitor panobinostat induces clinical responses with associated alterations in gene expression profiles in cutaneous T-cell lymphoma. Clin. Cancer Res. 2008, 14, 4500–4510. [Google Scholar] [CrossRef] [Green Version]
- Bouyahya, A.; El Omari, N.; Bakha, M.; Aanniz, T.; El Menyiy, N.; El Hachlafi, N.; El Baaboua, A.; El-Shazly, M.; Alshahrani, M.M.; Al Awadh, A.A.; et al. Pharmacological Properties of Trichostatin A, Focusing on the Anticancer Potential: A Comprehensive Review. Pharmaceuticals 2022, 15, 1235. [Google Scholar] [CrossRef] [PubMed]
- Pajak, B.; Gajkowska, B.; Orzechowski, A. Molecular basis of parthenolide-dependent proapoptotic activity in cancer cells. Folia Histochem. Cytobiol. 2008, 46, 129–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sztiller-Sikorska, M.; Czyz, M. Parthenolide as Cooperating Agent for Anti-Cancer Treatment of Various Malignancies. Pharmaceuticals 2020, 13, 194. [Google Scholar] [CrossRef]
- Gandhi, V.; Plunkett, W.; Cortes, J.E. Omacetaxine: A protein translation inhibitor for treatment of chronic myelogenous leukemia. Clin. Cancer Res. 2014, 20, 1735–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Guo, L.; Chen, Y.; Jiang, Y.; Wierda, W.G.; Plunkett, W. Homoharringtonine reduced Mcl-1 expression and induced apoptosis in chronic lymphocytic leukemia. Blood 2011, 117, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Vaklavas, C.; Chatzizisis, Y.S.; Tsimberidou, A.M. Common cardiovascular medications in cancer therapeutics. Pharmacol. Ther. 2011, 130, 177–190. [Google Scholar] [CrossRef]
- Braekeveldt, N.; Wigerup, C.; Gisselsson, D.; Mohlin, S.; Merselius, M.; Beckman, S.; Jonson, T.; Borjesson, A.; Backman, T.; Tadeo, I.; et al. Neuroblastoma patient-derived orthotopic xenografts retain metastatic patterns and geno- and phenotypes of patient tumours. Int. J. Cancer 2015, 136, E252–E261. [Google Scholar] [CrossRef] [Green Version]
- Khanna, C.; Jaboin, J.J.; Drakos, E.; Tsokos, M.; Thiele, C.J. Biologically relevant orthotopic neuroblastoma xenograft models: Primary adrenal tumor growth and spontaneous distant metastasis. In Vivo 2002, 16, 77–85. [Google Scholar]
- DeRose, Y.S.; Wang, G.; Lin, Y.C.; Bernard, P.S.; Buys, S.S.; Ebbert, M.T.; Factor, R.; Matsen, C.; Milash, B.A.; Nelson, E.; et al. Tumor grafts derived from women with breast cancer authentically reflect tumor pathology, growth, metastasis and disease outcomes. Nat. Med. 2011, 17, 1514–1520. [Google Scholar] [CrossRef]
- Weiss, W.A.; Aldape, K.; Mohapatra, G.; Feuerstein, B.G.; Bishop, J.M. Targeted expression of N-myc causes neuroblastoma in transgenic mice. EMBO J. 1997, 16, 2985–2995. [Google Scholar] [CrossRef]
- Isgro, M.A.; Bottoni, P.; Scatena, R. Neuron-Specific Enolase as a Biomarker: Biochemical and Clinical Aspects. Adv. Exp. Med. Biol. 2015, 867, 125–143. [Google Scholar]
- Kintzel, K.; Sonntag, J.; Strauss, E.; Obladen, M. Neuron-specific enolase: Reference values in cord blood. Clin. Chem. Lab. Med. 1998, 36, 245–247. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Compound Nature | IC50 | Ref. |
|---|---|---|---|
| 5′-iodotubercidin | Adenosine kinase (ADK) inhibitor. | 0.4 µM | [22,23,24] |
| A674563 | AKT1 inhibitor induces apoptosis through p53 and PARP cleavage. | 0.28 µM | [25] |
| Dactinomycin | Also known as actinomycin D preventing RNA elongation, used to treat Wilms tumor, rhabdomyosarcoma, Ewing’s sarcoma, trophoblastic neoplasm, testicular cancer, and certain types of ovarian cancer. | 0.03 µM | [26] |
| Plicamycin | RNA synthesis inhibitor, used to treat testicular cancer, Paget’s disease of bone. | 0.062 µM | [27] |
| Daunorubicin-HCl | Daunorubicin exhibits cytotoxic activity through topoisomerase-mediated interaction with DNA, thereby inhibiting DNA replication, repair, RNA, and protein synthesis, used to treat AML, ALL, CML, and Kaposi’s sarcoma. | 0.35 µM | [28,29] |
| Idarubicin-HCl | It is an anthracycline anti-leukemic drug. | 0.68 µM | [28,29] |
| Mitoxantrone | Topoisomerase II inhibitor, used to treat metastatic breast cancer, acute myeloid leukemia, and non-Hodgkin’s lymphoma. | 0.83 µM | [30,31] |
| Romidepsin | Natural product histone deacetylase inhibitor (HDACi), used in cutaneous T-cell lymphoma (CTCL) and peripheral T-cell lymphomas (PTCLs). | 0.2 µM | [32] |
| Panobinostat | Non-selective pan-histone HDACi (hydroxamic acid), FDA approval for use in patients with multiple myeloma. | 0.11 µM | [33] |
| Trichostatin (TSA) | Class I and II mammalian HDACi family of enzymes inhibit cell cycle progression. | 0.086 µM | [34] |
| Parthenolide | It is a sesquiterpene lactone of the germacranolide class. It inhibits HDAC1 protein, which leads to sustained DNA damage response in certain cells. | 0.2 µM | [35,36] |
| Homoharringtonine (Omacetaxine) | Natural extract, it inhibits protein translation by preventing the initial elongation step of protein synthesis for the treatment of chronic myeloid leukemia (CML). | 0.034 µM | [21,37,38] |
| Lanatoside C | A cardiac glucoside (CG). It was used in complex treatment of patients with heart failure and supraventricular arrhythmias. | 0.65 µM | [39] |
| Proscillaridin | A cardiac glucoside (CG). It was used in the treatment of congestive heart failure and cardiac arrhythmia. | 0.013 µM | [39] |
| Scillaren A | A crystalline steroidal glycoside present in squill or sea onion, which can be hydrolyzed to glucose and proscillaridin A and used as CG. | 0.04 µM | [39] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, M.S.; Chen, C. Small Molecules Targeting INSM1 for the Treatment of High-Risk Neuroblastoma. Biology 2023, 12, 1134. https://doi.org/10.3390/biology12081134
Lan MS, Chen C. Small Molecules Targeting INSM1 for the Treatment of High-Risk Neuroblastoma. Biology. 2023; 12(8):1134. https://doi.org/10.3390/biology12081134
Chicago/Turabian StyleLan, Michael S., and Chiachen Chen. 2023. "Small Molecules Targeting INSM1 for the Treatment of High-Risk Neuroblastoma" Biology 12, no. 8: 1134. https://doi.org/10.3390/biology12081134