
The Role of Sex Differences in Bone Health and Healing

Abstract
:Simple Summary
Abstract
1. Introduction
2. Sex Differences in Bone Health and Healing
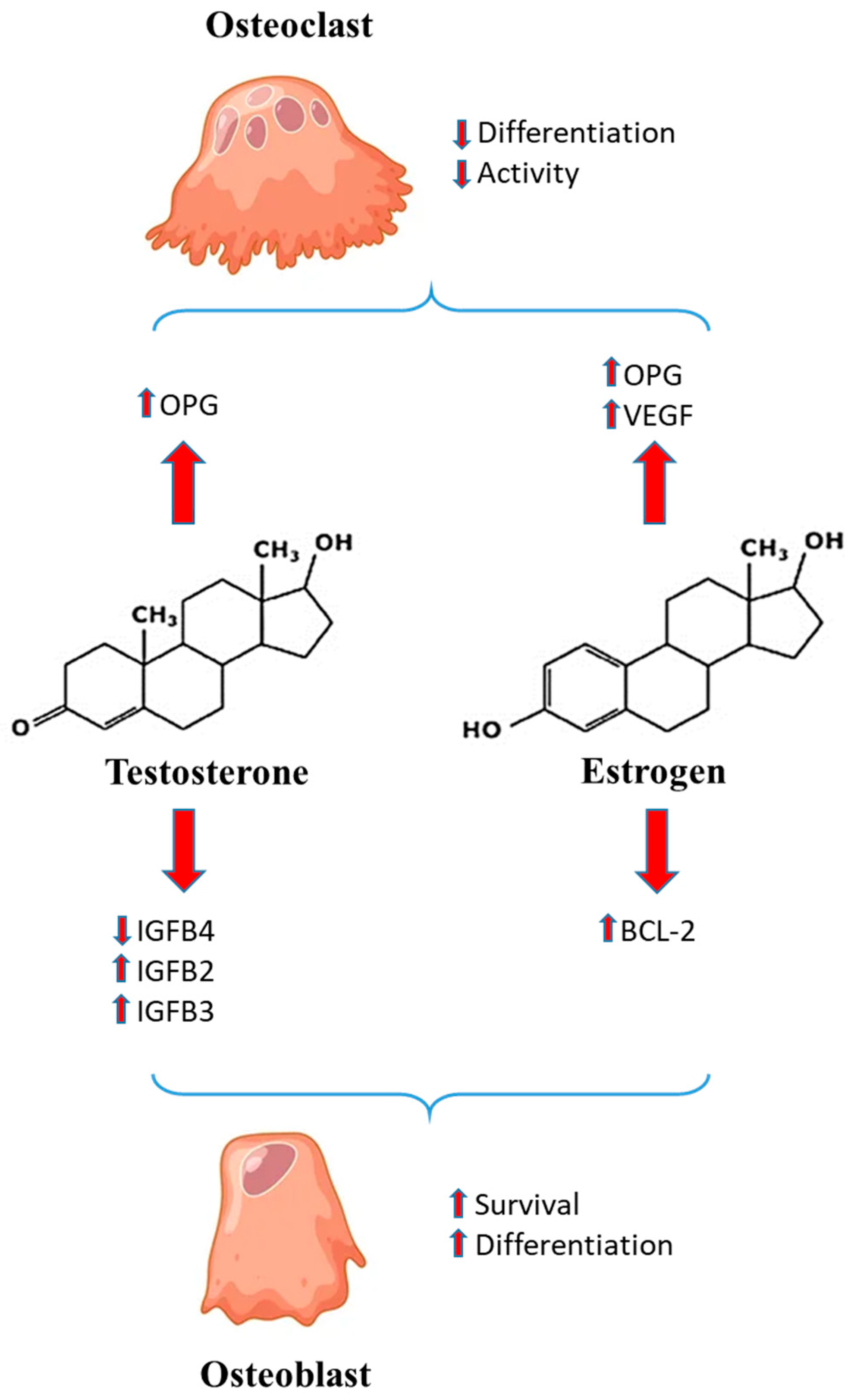
3. Sex Hormones and Bone Healing
 = induction
= induction  = inhibition.
= inhibition.4. Inflammatory Milieu and Sex Differences
5. Cell Mechanisms
6. Other Molecular Factors
7. Pediatric Age and Sex Differences in Bone Healing
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mehta, L.S.; Beckie, T.M.; DeVon, H.A.; Grines, C.L.; Krumholz, H.M.; Johnson, M.N.; Lindley, K.J.; Vaccarino, V.; Wang, T.Y.; Watson, K.E.; et al. Acute Myocardial Infarction in Women: A Scientific Statement from the American Heart Association. Circulation 2016, 133, 916–947. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, N.T.; Mielke, M.M. Sex Differences in Alzheimer’s Disease. Neurol. Clin. 2023, 41, 343–358. [Google Scholar] [CrossRef]
- Murphy, G.; Isenberg, D. Effect of gender on clinical presentation in systemic lupus erythematosus. Rheumatology 2013, 52, 2108–2115. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, H.; Klein, S.L. Sex Differences in Immunity to Viral Infections. Front. Immunol. 2021, 12, 720952. [Google Scholar] [CrossRef]
- Bellenghi, M.; Puglisi, R.; Pontecorvi, G.; De Feo, A.; Carè, A.; Mattia, G. Sex and Gender Disparities in Melanoma. Cancers 2020, 12, 1819. [Google Scholar] [CrossRef]
- Jenkins, C. Differences Between Men and Women with Chronic Obstructive Pulmonary Disease. Clin. Chest Med. 2021, 42, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Solarino, G.; Bizzoca, D.; Moretti, A.M.; D’Apolito, R.; Moretti, B.; Zagra, L. Sex and Gender-Related Differences in the Outcome of Total Hip Arthroplasty: A Current Concepts Review. Medicina 2022, 58, 1702. [Google Scholar] [CrossRef] [PubMed]
- Pesce, V.; Vicenti, G.; Picca, G.; Rifino, F.; Carrozzo, M.; Moretti, B. A review of gender differences in proximal humerus fractures. J. Sex Gend. Specif. Med. 2016, 2, 141–145. [Google Scholar] [CrossRef]
- Fluck, D.; Lisk, R.; Yeong, K.; Mahmood, R.; Robin, J.; Fry, C.H.; Han, T.S. Sex differences in clinical outcomes amongst 1105 patients admitted with hip fractures. Intern. Emerg. Med. 2023. [Google Scholar] [CrossRef] [PubMed]
- Osipov, B.; Paralkar, M.P.; Emami, A.J.; Cunningham, H.C.; Tjandra, P.M.; Pathak, S.; Langer, H.T.; Baar, K.; Christiansen, B.A. Sex differences in systemic bone and muscle loss following femur fracture in mice. J. Orthop. Res. 2022, 40, 878–890. [Google Scholar] [CrossRef]
- Patel, J.; Chen, S.; Katzmeyer, T.; Pei, Y.A.; Pei, M. Sex-dependent variation in cartilage adaptation: From degeneration to regeneration. Biol. Sex Differ. 2023, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- Branche, K.; Bradsell, H.L.; Lencioni, A.; Frank, R.M. Sex-Based Differences in Adult ACL Reconstruction Outcomes. Curr. Rev. Musculoskelet. Med. 2022, 15, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Edison, B.R.; Pandya, N.; Patel, N.M.; Carter, C.W. Sex and Gender Differences in Pediatric Knee Injuries. Clin. Sports Med. 2022, 41, 769–787. [Google Scholar] [CrossRef]
- Morochovič, R.; Takáčová, K.; Tomčovčík, Ľ.; Cibur, P.; Burda, R. Factors influencing femoral neck fracture healing after internal fixation with dynamic locking plate. Arch. Orthop. Trauma Surg. 2019, 139, 629–638. [Google Scholar] [CrossRef]
- Claes, L.E.; Heigele, C.A. Magnitudes of local stress and strain along bony surfaces predict the course and type of fracture healing. J. Biomech. 1999, 32, 255–266. [Google Scholar] [CrossRef]
- Claes, L.; Wolf, S.; Augat, P. Mechanical modification of callus healing. Chirurg 2000, 71, 989–994. [Google Scholar] [CrossRef]
- Tewari, P.; Sweeney, B.F.; Lemos, J.L.; Shapiro, L.; Gardner, M.J.; Morris, A.M.; Baker, L.C.; Harris, A.S.; Kamal, R.N. Evaluation of Systemwide Improvement Programs to Optimize Time to Surgery for Patients With Hip Fractures: A Systematic Review. JAMA Netw. Open. 2022, 5, e2231911. [Google Scholar] [CrossRef]
- Robinson, Y.; Heyde, C.E.; Försth, P.; Olerud, C. Kyphoplasty in osteoporotic vertebral compression fractures--guidelines and technical considerations. J. Orthop. Surg. Res. 2011, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Nikitovic, M.; Wodchis, W.P.; Krahn, M.D.; Cadarette, S.M. Direct health-care costs attributed to hip fractures among seniors: A matched cohort study. Osteoporos. Int. 2013, 24, 659–669. [Google Scholar] [CrossRef] [Green Version]
- Alpantaki, K.; Dohm, M.; Korovessis, P.; Hadjipavlou, A.G. Surgical options for osteoporotic vertebral compression fractures complicated with spinal deformity and neurologic deficit. Injury 2018, 49, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Hoyt, D.; Urits, I.; Orhurhu, V.; Orhurhu, M.S.; Callan, J.; Powell, J.; Manchikanti, L.; Kaye, A.D.; Kaye, R.J.; Viswanath, O. Current Concepts in the Management of Vertebral Compression Fractures. Curr. Pain Headache Rep. 2020, 24, 16. [Google Scholar] [CrossRef] [PubMed]
- Einhorn, T.A.; Gerstenfeld, L.C. Fracture healing: Mechanisms and interventions. Nat. Rev. Rheumatol. 2015, 11, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameis, A.; Randhawa, K.; Yu, H.; Côté, P.; Haldeman, S.; Chou, R.; Hurwitz, E.L.; Nordin, M.; Wong, J.J.; Shearer, H.M.; et al. The Global Spine Care Initiative: A review of reviews and recommendations for the non-invasive management of acute osteoporotic vertebral compression fracture pain in low- and middle-income communities. Eur. Spine J. 2018, 27 (Suppl. S6), 861–869. [Google Scholar] [CrossRef] [PubMed]
- Toosi, S.; Behravan, N.; Behravan, J. Nonunion fractures, mesenchymal stem cells and bone tissue engineering. J. Biomed. Mater. Res. A 2018, 106, 2552–2562. [Google Scholar] [CrossRef]
- Seeman, E. Clinical review 137: Sexual dimorphism in skeletal size, density, and strength. J. Clin. Endocrinol. Metab. 2001, 86, 4576–4584. [Google Scholar] [CrossRef]
- Cummings, S.R.; Black, D.M.; Rubin, S.M. Lifetime risks of hip, Colles’, or vertebral fracture and coronary heart disease among white postmenopausal women. Arch. Intern. Med. 1989, 149, 2445–2448. [Google Scholar] [CrossRef]
- Cummings, S.R.; Melton, L.J. Epidemiology and outcomes of osteoporotic fractures. Lancet 2002, 359, 1761–1767. [Google Scholar] [CrossRef]
- Rupp, M.; Kern, S.; El Khassawna, T.; Ismat, A.; Malhan, D.; Alt, V.; Heiss, C.; Raschke, M.J. Do Systemic Factors Influence the Fate of Nonunions to Become Atrophic? A Retrospective Analysis of 162 Cases. BioMed Res. Int. 2019, 2019, 6407098. [Google Scholar] [CrossRef]
- Li, X.P.; Li, X.Y.; Yang, M.H.; Zhu, S.W.; Wu, X.B.; Zhang, P. Changes of bone turnover markers after elderly hip fracture surgery. J. Bone Miner. Metab. 2021, 39, 237–244. [Google Scholar] [CrossRef]
- Johnson, J.P.; Kleiner, J.; Goodman, A.D.; Gil, J.A.; Daniels, A.H.; Hayda, R.A. Treatment of femoral neck fractures in patients 45–64 years of age. Injury 2019, 50, 708–712. [Google Scholar] [CrossRef]
- Lofrese, G.; Musio, A.; De Iure, F.; Cultrera, F.; Martucci, A.; Iaccarino, C.; Ibn Essayed, W.; Ghadirpour, R.; Servadei, F.; Cavallo, M.A.; et al. Type II odontoid fracture in elderly patients treated conservatively: Is fracture healing the goal? Eur. Spine J. 2019, 28, 1064–1071. [Google Scholar] [CrossRef]
- Poole, K.E.; Compston, J.E. Osteoporosis and its management. BMJ 2006, 333, 1251–1256. [Google Scholar] [CrossRef]
- Endo, Y.; Aharonoff, G.B.; Zuckerman, J.D.; Egol, K.A.; Koval, K.J. Gender differences in patients with hip fracture: A greater risk of morbidity and mortality in men. J. Orthop. Trauma 2005, 19, 29–35. [Google Scholar] [CrossRef]
- Manolagas, S.C.; Kousteni, S.; Jilka, R.L. Sex steroids and bone. Recent Prog. Horm. Res. 2002, 57, 385–409. [Google Scholar] [CrossRef]
- Kousteni, S.; Bellido, T.; Plotkin, L.I.; O’Brien, C.A.; Bodenner, D.L.; Han, L.; Han, K.; Digregorio, G.B.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S.; et al. Nongenotropic, sex-nonspecific signaling through the estrogen or androgen receptors: Dissociation from transcriptional activity. Cell 2001, 104, 719–730. [Google Scholar] [CrossRef]
- Kodama, I.; Niida, S.; Sanada, M.; Yoshiko, Y.; Tsuda, M.; Maeda, N.; Ohama, K. Estrogen regulates the production of VEGF for osteoclast formation and activity in op/op mice. J. Bone Miner. Res. 2004, 19, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Ono, W.; Sakagami, N.; Nishimori, S.; Ono, N.; Kronenberg, H.M. Parathyroid hormone receptor signalling in osterix-expressing mesenchymal progenitors is essential for tooth root formation. Nat. Commun. 2016, 7, 11277. [Google Scholar] [CrossRef] [Green Version]
- Sousa-Nunes, R.; Yee, L.L.; Gould, A.P. Fat cells reactivate quiescent neuroblasts via TOR and glial insulin relays in Drosophila. Nature 2011, 471, 508–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, L.E.; Soliman, S.S.; Li, X.; Bilder, D. Altered modes of stem cell division drive adaptive intestinal growth. Cell 2011, 147, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakada, D.; Oguro, H.; Levi, B.P.; Ryan, N.; Kitano, A.; Saitoh, Y.; Takeichi, M.; Wendt, G.R.; Morrison, S.J. Oestrogen increases haematopoietic stem-cell self-renewal in females and during pregnancy. Nature 2014, 505, 555–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velardi, E.; Tsai, J.J.; Radtke, S.; Cooper, K.; Argyropoulos, K.V.; Jae-Hung, S.; Young, L.F.; Lazrak, A.; Smith, O.M.; Lieberman, S.; et al. Suppression of luteinizing hormone enhances HSC recovery after hematopoietic injury. Nat. Med. 2018, 24, 239–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herber, C.B.; Krause, W.C.; Wang, L.; Bayrer, J.R.; Li, A.; Schmitz, M.; Fields, A.; Ford, B.; Zhang, Z.; Reid, M.S.; et al. Estrogen signaling in arcuate Kiss1 neurons suppresses a sex-dependent female circuit promoting dense strong bones. Nat. Commun. 2019, 10, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrew, T.W.; Koepke, L.S.; Wang, Y.; Lopez, M.; Steininger, H.; Struck, D.; Boyko, T.; Ambrosi, T.H.; Tong, X.; Sun, Y.; et al. Sexually dimorphic estrogen sensing in skeletal stem cells controls skeletal regeneration. Nat. Commun. 2022, 13, 6491. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B.F.; Xing, L. Biology of RANK, RANKL, and osteoprotegerin. Arthritis Res. Ther. 2007, 9 (Suppl. S1), S1. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.P.; Boyd, J.; Frank, G.R.; Takahashi, H.; Cohen, R.M.; Specker, B.; Williams, T.C.; Lubahn, D.B.; Korach, K.S. Estrogen resistance caused by a mutation in the estrogen-receptor gene in a man. N. Engl. J. Med. 1994, 331, 1056–1061. [Google Scholar] [CrossRef]
- Carani, C.; Qin, K.; Simoni, M.; Faustini-Fustini, M.; Serpente, S.; Boyd, J.; Korach, K.S.; Simpson, E.R. Effect of testosterone and estradiol in a man with aromatase deficiency. N. Engl. J. Med. 1997, 337, 91–95. [Google Scholar] [CrossRef]
- Gori, F.; Hofbauer, L.C.; Conover, C.A.; Khosla, S. Effects of androgens on the insulin-like growth factor system in an androgen-responsive human osteoblastic cell line. Endocrinology 1999, 140, 5579–5586. [Google Scholar] [CrossRef]
- Chen, Q.; Kaji, H.; Kanatani, M.; Sugimoto, T.; Chihara, K. Testosterone increases osteoprotegerin mRNA expression in mouse osteoblast cells. Horm. Metab. Res. 2004, 36, 674–678. [Google Scholar] [CrossRef]
- Hofbauer, L.C.; Hicok, K.C.; Chen, D.; Khosla, S. Regulation of osteoprotegerin production by androgens and anti-androgens in human osteoblastic lineage cells. Eur. J. Endocrinol. 2002, 147, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Venken, K.; De Gendt, K.; Boonen, S.; Ophoff, J.; Bouillon, R.; Swinnen, J.V.; Verhoeven, G.; Vanderschueren, D. Relative impact of androgen and estrogen receptor activation in the effects of androgens on trabecular and cortical bone in growing male mice: A study in the androgen receptor knockout mouse model. J. Bone Miner. Res. 2006, 21, 576–585. [Google Scholar] [CrossRef]
- Claes, L.; Recknagel, S.; Ignatius, A. Fracture healing under healthy and inflammatory conditions. Nat. Rev. Rheumatol. 2012, 8, 133–143. [Google Scholar] [CrossRef]
- Bastian, O.W.; Kuijer, A.; Koenderman, L.; Stellato, R.K.; van Solinge, W.W.; Leenen, L.P.; Blokhuis, T.J. Impaired bone healing in multitrauma patients is associated with altered leukocyte kinetics after major trauma. J. Inflamm. Res. 2016, 9, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt-Bleek, K.; Kwee, B.J.; Mooney, D.J.; Duda, G.N. Boon and Bane of Inflammation in Bone Tissue Regeneration and Its Link with Angiogenesis. Tissue Eng. Part B Rev. 2015, 21, 354–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toben, D.; Schroeder, I.; El Khassawna, T.; Mehta, M.; Hoffmann, J.-E.; Frisch, J.-T.; Schell, H.; Lienau, J.; Serra, A.; Radbruch, A.; et al. Fracture healing is accelerated in the absence of the adaptive immune system. J. Bone Miner. Res. 2011, 26, 113–124. [Google Scholar] [CrossRef]
- Reinke, S.; Geissler, S.; Taylor, W.R.; Schmidt-Bleek, K.; Juelke, K.; Schwachmeyer, V.; Dahne, M.; Hartwig, T.; Akyüz, L.; Meisel, C.; et al. Terminally differentiated CD8+ T cells negatively affect bone regeneration in humans. Sci. Transl. Med. 2013, 5, 177ra36. [Google Scholar] [CrossRef] [PubMed]
- Kovtun, A.; Bergdolt, S.; Wiegner, R.; Radermacher, P.; Huber-Lang, M.; Ignatius, A. The crucial role of neutrophil granulocytes in bone fracture healing. Eur. Cells Mater. 2016, 32, 152–162. [Google Scholar] [CrossRef]
- Haffner-Luntzer, M.; Fischer, V.; Prystaz, K.; Liedert, A.; Ignatius, A. The inflammatory phase of fracture healing is influenced by oestrogen status in mice. Eur. J. Med. Res. 2017, 22, 23. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Qian, W.-P.; Dark, K.; Toraldo, G.; Lin, A.S.P.; Guldberg, R.E.; Flavell, R.A.; Weitzmann, M.N.; Pacifici, R. Estrogen prevents bone loss through transforming growth factor beta signaling in T cells. Proc. Natl. Acad. Sci. USA 2004, 101, 16618–16623. [Google Scholar] [CrossRef]
- Cenci, S.; Toraldo, G.; Weitzmann, M.N.; Roggia, C.; Gao, Y.; Qian, W.P.; Sierra, O.; Pacifici, R. Estrogen deficiency induces bone loss by increasing T cell proliferation and lifespan through IFN-gamma-induced class II transactivator. Proc. Natl. Acad. Sci. USA 2003, 100, 10405–10410. [Google Scholar] [CrossRef]
- Gao, H.; Huang, J.; Wei, Q.; He, C. Advances in Animal Models for Studying Bone Fracture Healing. Bioengineering 2023, 10, 201. [Google Scholar] [CrossRef]
- Andrew, J.G.; Andrew, S.M.; Freemont, A.J.; Marsh, D.R. Inflammatory cells in normal human fracture healing. Acta Orthop. Scand. 1994, 65, 462–466. [Google Scholar] [CrossRef] [Green Version]
- Chung, R.; Cool, J.C.; Scherer, M.A.; Foster, B.K.; Xian, C.J. Roles of neutrophil-mediated inflammatory response in the bony repair of injured growth plate cartilage in young rats. J. Leukoc. Biol. 2006, 80, 1272–1280. [Google Scholar] [CrossRef]
- McKibbin, B. The biology of fracture healing in long bones. J. Bone Jt. Surg. Br. 1978, 60-B, 150–162. [Google Scholar] [CrossRef]
- Nathan, K.; Lu, L.Y.; Lin, T.; Pajarinen, J.; Jämsen, E.; Huang, J.-F.; Romero-Lopez, M.; Maruyama, M.; Kohno, Y.; Yao, Z.; et al. Precise immunomodulation of the M1 to M2 macrophage transition enhances mesenchymal stem cell osteogenesis and differs by sex. Bone Jt. Res. 2019, 8, 481–488. [Google Scholar] [CrossRef]
- Pajarinen, J.; Lin, T.; Gibon, E.; Kohno, Y.; Maruyama, M.; Nathan, K.; Lu, L.; Yao, Z.; Goodman, S.B. Mesenchymal stem cell-macrophage crosstalk and bone healing. Biomaterials 2019, 196, 80–89. [Google Scholar] [CrossRef]
- Niemann, T.; Greiner, J.F.W.; Kaltschmidt, C.; Kaltschmidt, B. EPO regulates neuronal differentiation of adult human neural-crest derived stem cells in a sex-specific manner. BMC Neurosci. 2023, 24, 19. [Google Scholar] [CrossRef]
- Gupta, S.; Khanal, S.; Bhavnani, N.; Mathias, A.; Lallo, J.; Kiriakou, A.; Ferrell, J.; Raman, P. Sex-specific differences in atherosclerosis, thrombospondin-1, and smooth muscle cell differentiation in metabolic syndrome versus non-metabolic syndrome mice. Front. Cardiovasc. Med. 2022, 9, 1020006. [Google Scholar] [CrossRef] [PubMed]
- Knewtson, K.E.; Ohl, N.R.; Robinson, J.L. Estrogen Signaling Dictates Musculoskeletal Stem Cell Behavior: Sex Differences in Tissue Repair. Tissue Eng. Part B Rev. 2022, 28, 789–812. [Google Scholar] [CrossRef] [PubMed]
- Mank, J.E.; Rideout, E.J. Developmental mechanisms of sex differences: From cells to organisms. Development 2021, 148, dev199750. [Google Scholar] [CrossRef]
- Steppe, L.; Bülow, J.; Tuckermann, J.; Ignatius, A.; Haffner-Luntzer, M. Bone Mass and Osteoblast Activity Are Sex-Dependent in Mice Lacking the Estrogen Receptor α in Chondrocytes and Osteoblast Progenitor Cells. Int. J. Mol. Sci. 2022, 23, 2902. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.B.; Cohen, D.J.; Olivares-Navarrete, R.; Williams, J.K.; Cochran, D.L.; Boyan, B.D.; Schwartz, Z. Human osteoblasts exhibit sexual dimorphism in their response to estrogen on microstructured titanium surfaces. Biol. Sex Differ. 2018, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.S.; Beyer, R.P.; Smyth, M.D.; Clarke, C.M.; Timms, A.E.; Bammler, T.K.; Stamper, B.D.; Mecham, B.H.; Gustafson, J.A.; Cunningham, M.L. Osteoblast differentiation profiles define sex specific gene expression patterns in craniosynostosis. Bone 2015, 76, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Lista, P.; Straface, E.; Brunelleschi, S.; Franconi, F.; Malorni, W. On the role of autophagy in human diseases: A gender perspective. J. Cell. Mol. Med. 2011, 15, 1443–1457. [Google Scholar] [CrossRef]
- Wu, C.; Chen, Y.; Su, Y.; Chiu, C.; Guan, S.; Yang, R.; Liu, S. Gender difference of CCAAT/enhancer binding protein homologous protein deficiency in susceptibility to osteopenia. J. Orthop. Res. 2019, 37, 942–947. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition). Autophagy 2021, 17, 1–382. [Google Scholar] [CrossRef]
- Camuzard, O.; Santucci-Darmanin, S.; Breuil, V.; Cros, C.; Gritsaenko, T.; Pagnotta, S.; Cailleteau, L.; Battaglia, S.; Panaïa-Ferrari, P.; Heymann, D.; et al. Sex-specific autophagy modulation in osteoblastic lineage: A critical function to counteract bone loss in female. Oncotarget 2016, 7, 66416–66428. [Google Scholar] [CrossRef] [Green Version]
- Haffner-Luntzer, M.; Fischer, V.; Ignatius, A. Differences in Fracture Healing Between Female and Male C57BL/6J Mice. Front. Physiol. 2021, 12, 712494. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, J.A.; Keshav, V.; Mitchell, A.; O’Connor, J.P. Local inhibition of 5-lipoxygenase enhances bone formation in a rat model. Bone Jt. Res. 2013, 2, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Moon, T.C.; Befus, A.D.; Kulka, M. Mast cell mediators: Their differential release and the secretory pathways involved. Front. Immunol. 2014, 5, 569. [Google Scholar] [CrossRef] [Green Version]
- Pace, S.; Pergola, C.; Dehm, F.; Rossi, A.; Gerstmeier, J.; Troisi, F.; Pein, H.; Schaible, A.M.; Weinigel, C.; Rummler, S.; et al. Androgen-mediated sex bias impairs efficiency of leukotriene biosynthesis inhibitors in males. J. Clin. Investig. 2017, 127, 3167–3176. [Google Scholar] [CrossRef] [Green Version]
- Hartman, M.L.; Veldhuis, J.D.; Thorner, M.O. Normal control of growth hormone secretion. Horm. Res. 1993, 40, 37–47. [Google Scholar] [CrossRef]
- Hobbs, C.J.; Plymate, S.R.; Rosen, C.J.; Adler, R.A. Testosterone administration increases insulin-like growth factor-I levels in normal men. J. Clin. Endocrinol. Metab. 1993, 77, 776–779. [Google Scholar] [CrossRef]
- Weissberger, A.J.; Ho, K.K. Activation of the somatotropic axis by testosterone in adult males: Evidence for the role of aromatization. J. Clin. Endocrinol. Metab. 1993, 76, 1407–1412. [Google Scholar] [CrossRef]
- Veldhuis, J.D.; Frystyk, J.; Iranmanesh, A.; Ørskov, H. Testosterone and estradiol regulate free insulin-like growth factor I (IGF-I), IGF binding protein 1 (IGFBP-1), and dimeric IGF-I/IGFBP-1 concentrations. J. Clin. Endocrinol. Metab. 2005, 90, 2941–2947. [Google Scholar] [CrossRef] [Green Version]
- Ashpole, N.M.; Logan, S.; Yabluchanskiy, A.; Mitschelen, M.C.; Yan, H.; Farley, J.A.; Hodges, E.L.; Ungvari, Z.; Csiszar, A.; Chen, S.; et al. IGF-1 has sexually dimorphic, pleiotropic, and time-dependent effects on healthspan, pathology, and lifespan. Geroscience 2017, 39, 129–145. [Google Scholar] [CrossRef] [Green Version]
- Strube, P.; Mehta, M.; Baerenwaldt, A.; Trippens, J.; Wilson, C.J.; Ode, A.; Perka, C.; Duda, G.N.; Kasper, G. Sex-specific compromised bone healing in female rats might be associated with a decrease in mesenchymal stem cell quantity. Bone 2009, 45, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.P.; Später, D.; Taketo, M.M.; Birchmeier, W.; Hartmann, C. Canonical Wnt/beta-catenin signaling prevents osteoblasts from differentiating into chondrocytes. Dev. Cell 2005, 8, 727–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Chen, J.; Gantz, M.; Punyanitya, M.; Heymsfield, S.B.; Gallagher, D.; Albu, J.; Engelson, E.; Kotler, D.; Pi-Sunyer, X.; et al. Ethnic and sex differences in bone marrow adipose tissue and bone mineral density relationship. Osteoporos Int. 2012, 23, 2293–2301. [Google Scholar] [CrossRef] [Green Version]
- Liney, G.P.; Bernard, C.P.; Manton, D.J.; Turnbull, L.W.; Langton, C.M. Age, gender, and skeletal variation in bone marrow composition: A preliminary study at 3.0 Tesla. J. Magn. Reson. Imaging 2007, 26, 787–793. [Google Scholar] [CrossRef]
- Mattioli, D.; Vinicola, V.; Aragona, M.; Montuori, M.; Tarantino, U.; Capuani, S. Behavior during aging of bone-marrow fatty-acids profile in women’s calcaneus to search for early potential osteoporotic biomarkers: A 1H-MR Spectroscopy study. Bone 2022, 164, 116514. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, G.; Capuani, S.; Manenti, G.; Vinicola, V.; Fusco, A.; Baldi, J.; Scimeca, M.; Hagberg, G.; Bozzali, M.; Simonetti, G.; et al. Bone Marrow Lipid Profiles from Peripheral Skeleton as Potential Biomarkers for Osteoporosis: A 1H-MR Spectroscopy Study. Acad. Radiol. 2016, 23, 273–283. [Google Scholar] [CrossRef]
- Pino, A.M.; Rodríguez, J.P. Is fatty acid composition of human bone marrow significant to bone health? Bone 2019, 118, 53–61. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Maynard, L.M.; Towne, B.; Roche, A.F.; Wisemandle, W.; Li, J.; Guo, S.S.; Chumlea, W.C.; Siervogel, R.M. Sex differences in bone mass acquisition during growth: The Fels Longitudinal Study. J. Clin. Densitom. 2001, 4, 147–157. [Google Scholar] [CrossRef]
- Baxter-Jones, A.D.G.; Jackowski, S.A. Sex differences in bone mineral content and bone geometry accrual: A review of the Paediatric Bone Mineral Accural Study (1991–2017). Ann. Hum. Biol. 2021, 48, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Gabel, L.; Macdonald, H.M.; McKay, H.A. Sex Differences and Growth-Related Adaptations in Bone Microarchitecture, Geometry, Density, and Strength from Childhood to Early Adulthood: A Mixed Longitudinal HR-pQCT Study. J. Bone Miner. Res. 2017, 32, 250–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabel, L.; Nettlefold, L.; Brasher, P.M.; Moore, S.A.; Ahamed, Y.; Macdonald, H.M.; McKay, H.A. Reexamining the Surfaces of Bone in Boys and Girls During Adolescent Growth: A 12-Year Mixed Longitudinal pQCT Study. J. Bone Miner. Res. 2015, 30, 2158–2167. [Google Scholar] [CrossRef] [Green Version]
- Lindaman, L.M. Bone healing in children. Clin. Podiatr. Med. Surg. 2001, 18, 97–108. [Google Scholar] [PubMed]


{kind=link}
| Pathology | Sex/Gender Differences | ||||
|---|---|---|---|---|---|
| Incidence | Course | Symptoms | Example | References | |
| Cardiovascular diseases | Yes | Yes | Yes | Infarction | [1] |
| Neurodegenerative Diseases | Yes | No | No | Alzheimer | [2] |
| Autoimmune diseases | Yes | Yes | Yes | Lupus | [3] |
| Infectious diseases | Yes | Yes | No | Hepatitis B | [4] |
| Cancers | Yes | Yes | No | Melanoma | [5] |
| Respiratory diseases | Yes | No | No | Chronic obstructive pulmonary disease | [6] |
| Orthopedics | Yes | Yes | No | Hip arthroplasty, Hip, Femur, Humerus fractures | [7,8,9,10] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortona, E.; Pagano, M.T.; Capossela, L.; Malorni, W. The Role of Sex Differences in Bone Health and Healing. Biology 2023, 12, 993. https://doi.org/10.3390/biology12070993
Ortona E, Pagano MT, Capossela L, Malorni W. The Role of Sex Differences in Bone Health and Healing. Biology. 2023; 12(7):993. https://doi.org/10.3390/biology12070993
Chicago/Turabian StyleOrtona, Elena, Maria Teresa Pagano, Lavinia Capossela, and Walter Malorni. 2023. "The Role of Sex Differences in Bone Health and Healing" Biology 12, no. 7: 993. https://doi.org/10.3390/biology12070993