
Effect of Ripening and In Vitro Digestion on Bioactive Peptides Profile in Ras Cheese and Their Biological Activities


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Ras Cheese Manufacturing and Compositional Analysis
2.3. Extraction of Water-Soluble Peptides (WSPs) from Ras Cheese
2.4. In Vitro Gastro-Intestinal Digestion of Ras Cheeses
2.5. Peptides Quantification and Analysis of the Biological Activities in Undigested and Digested Ras Cheeses
2.6. Peptidomics Analysis and Label-Free MS Peak Quantification
2.7. Bioactive Peptides Identification and Semi-Quantitative Analysis, and Absolute Quantification of the Bioactive Peptides VPP, IPP and APFPE
2.8. Statistical Analysis
3. Results and Discussion
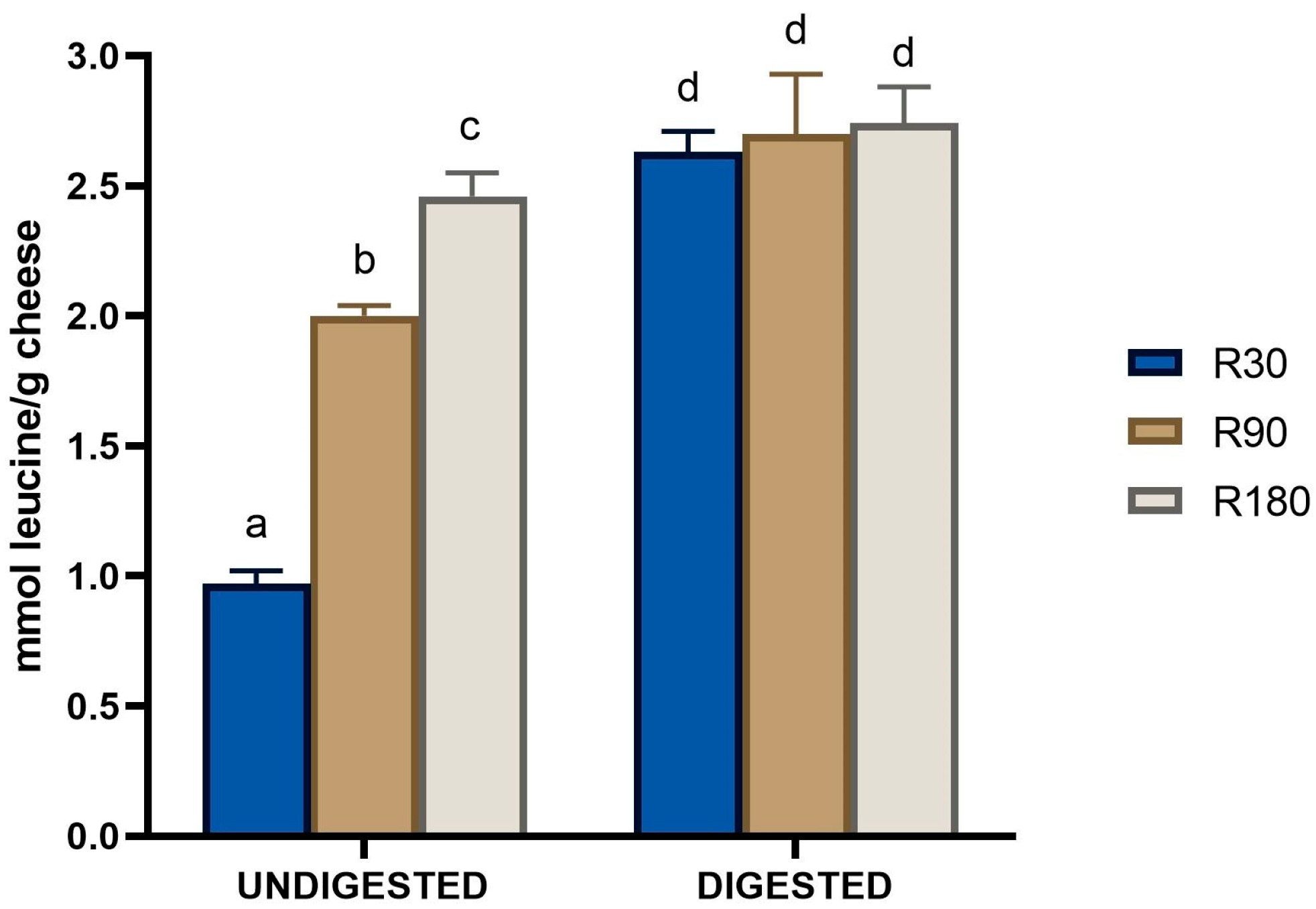
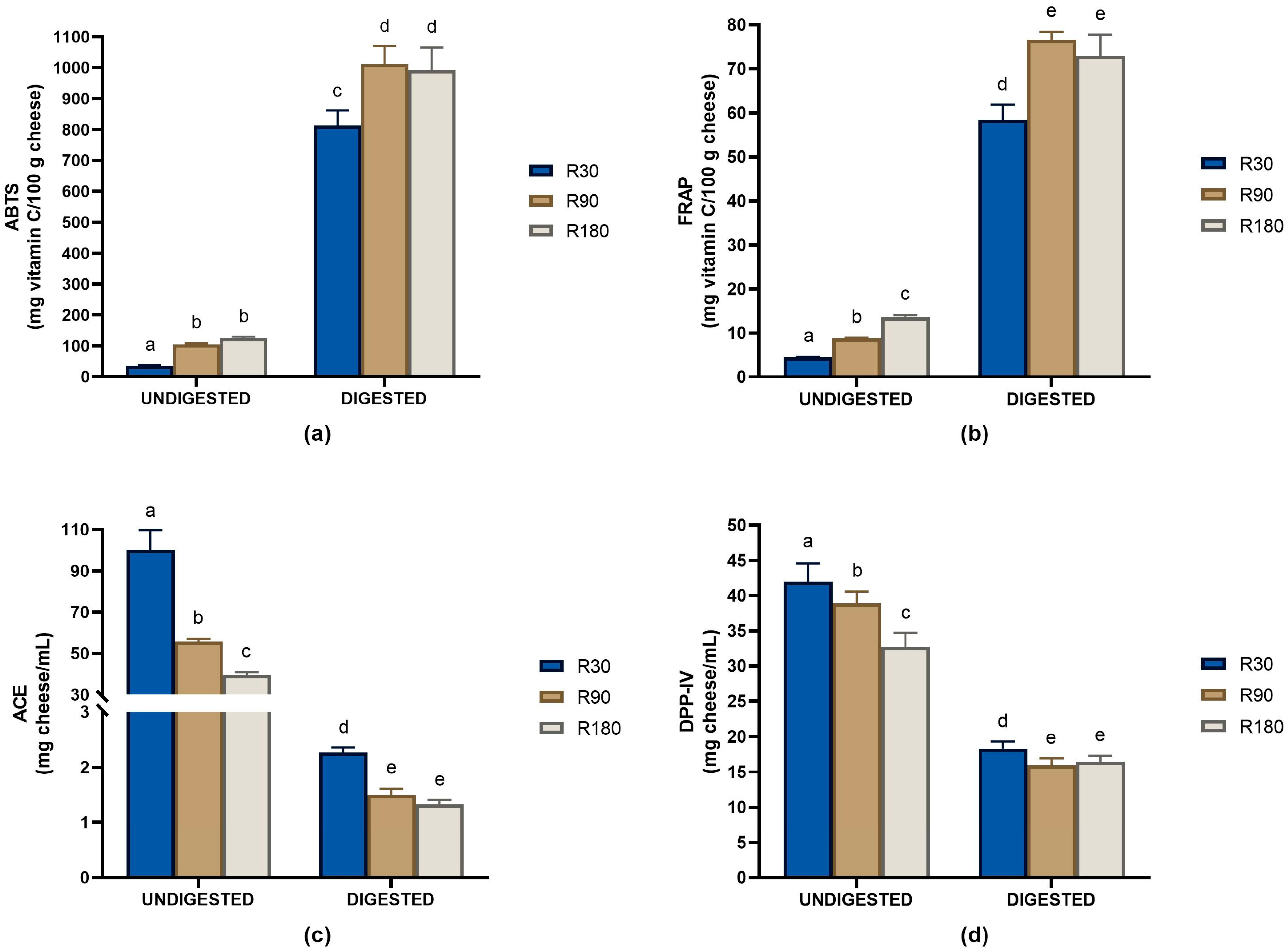
3.1. Peptide Quantification and Biological Activities of Undigested and Digested Ras Cheeses at Different Ripening Times
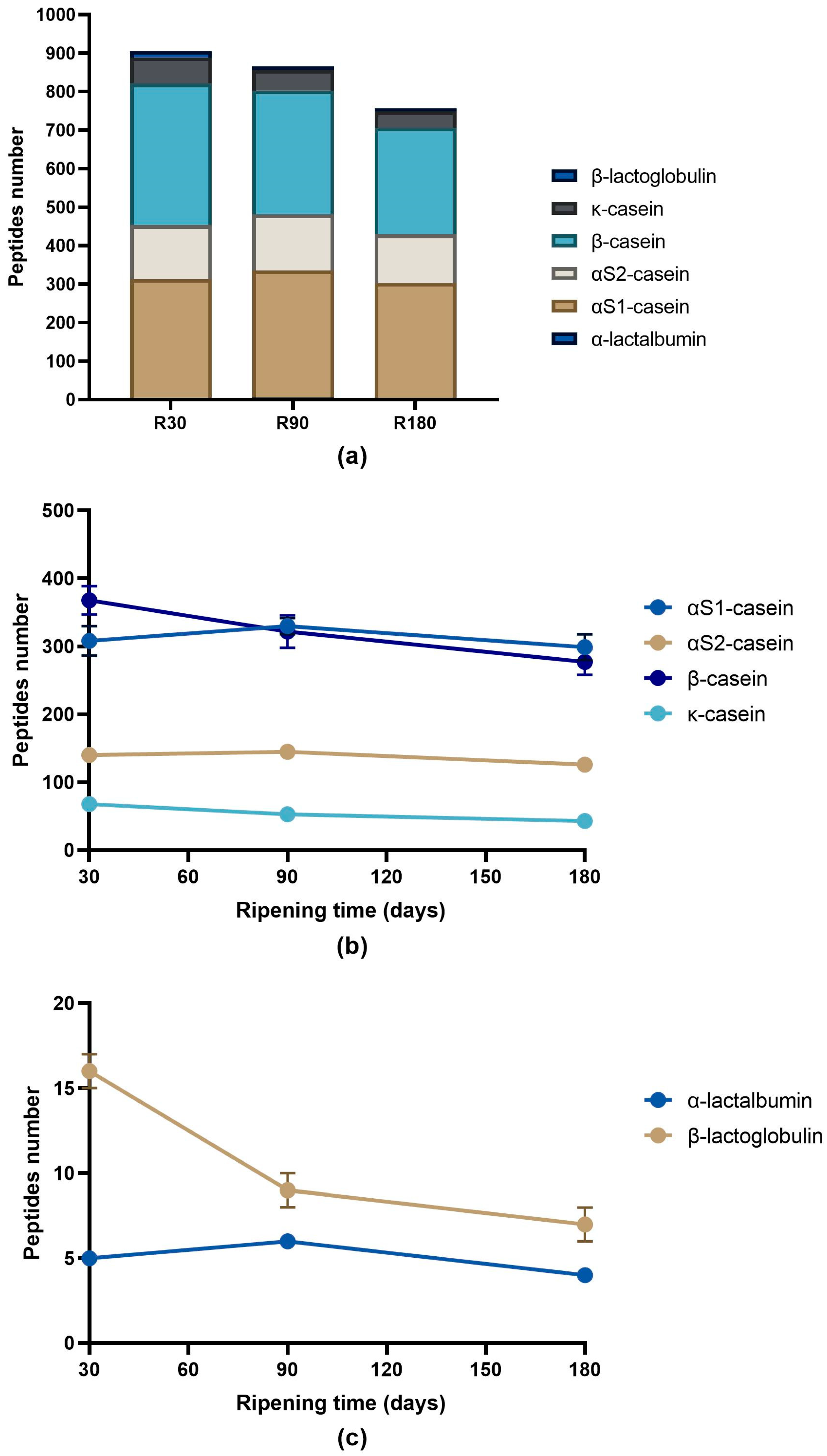
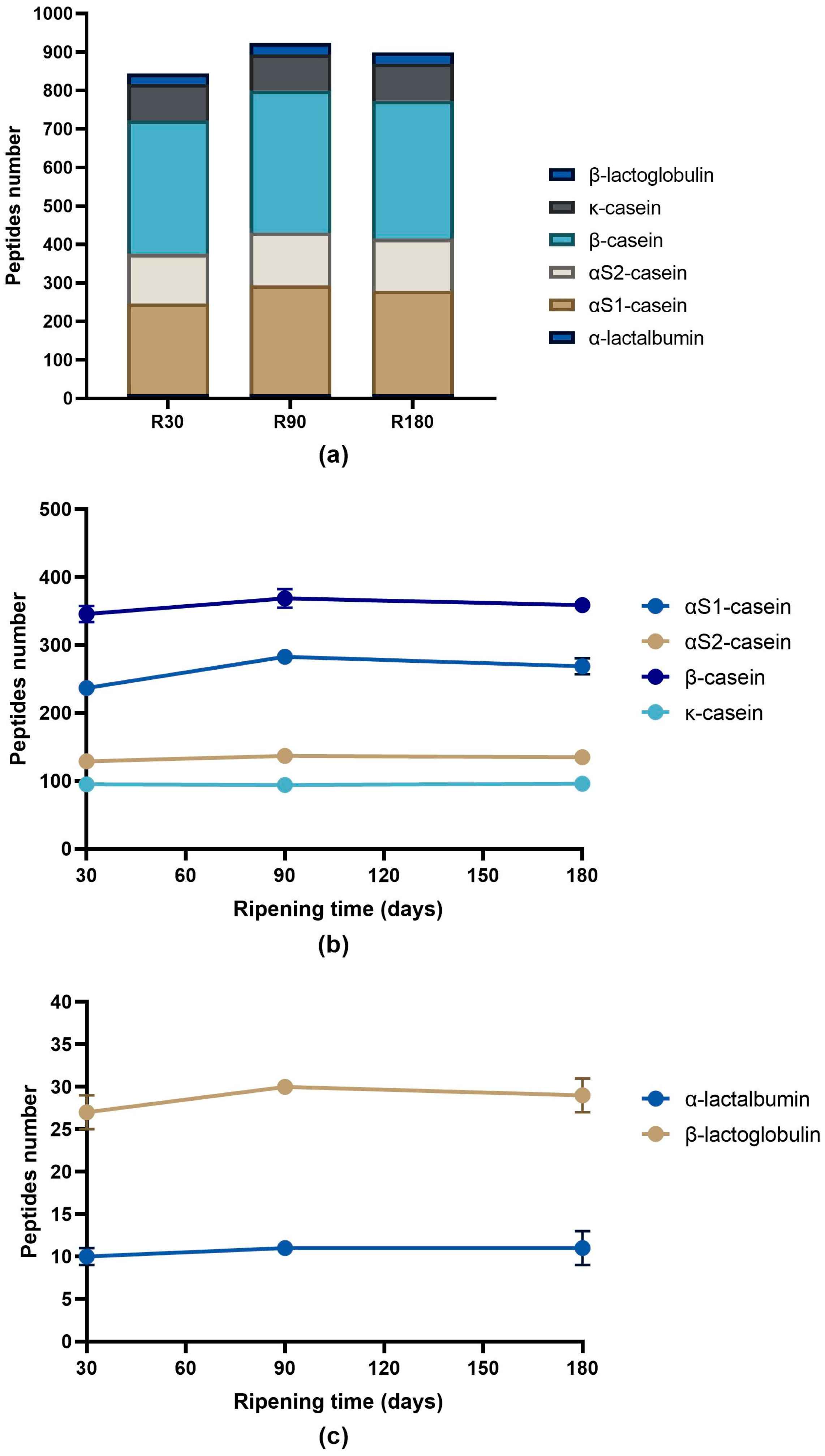
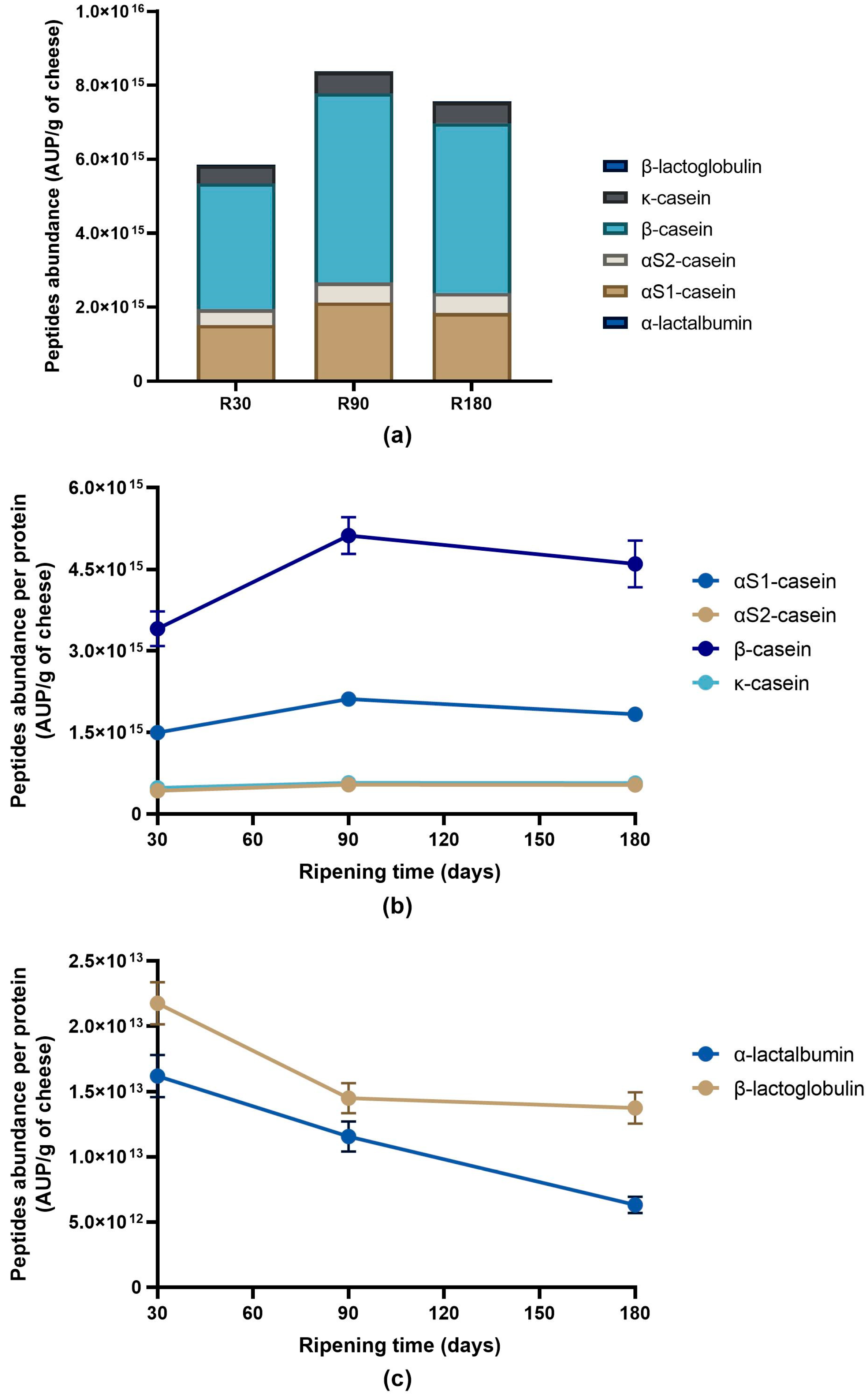
3.2. Peptide Profile Evolution during Ras Cheese Ripening
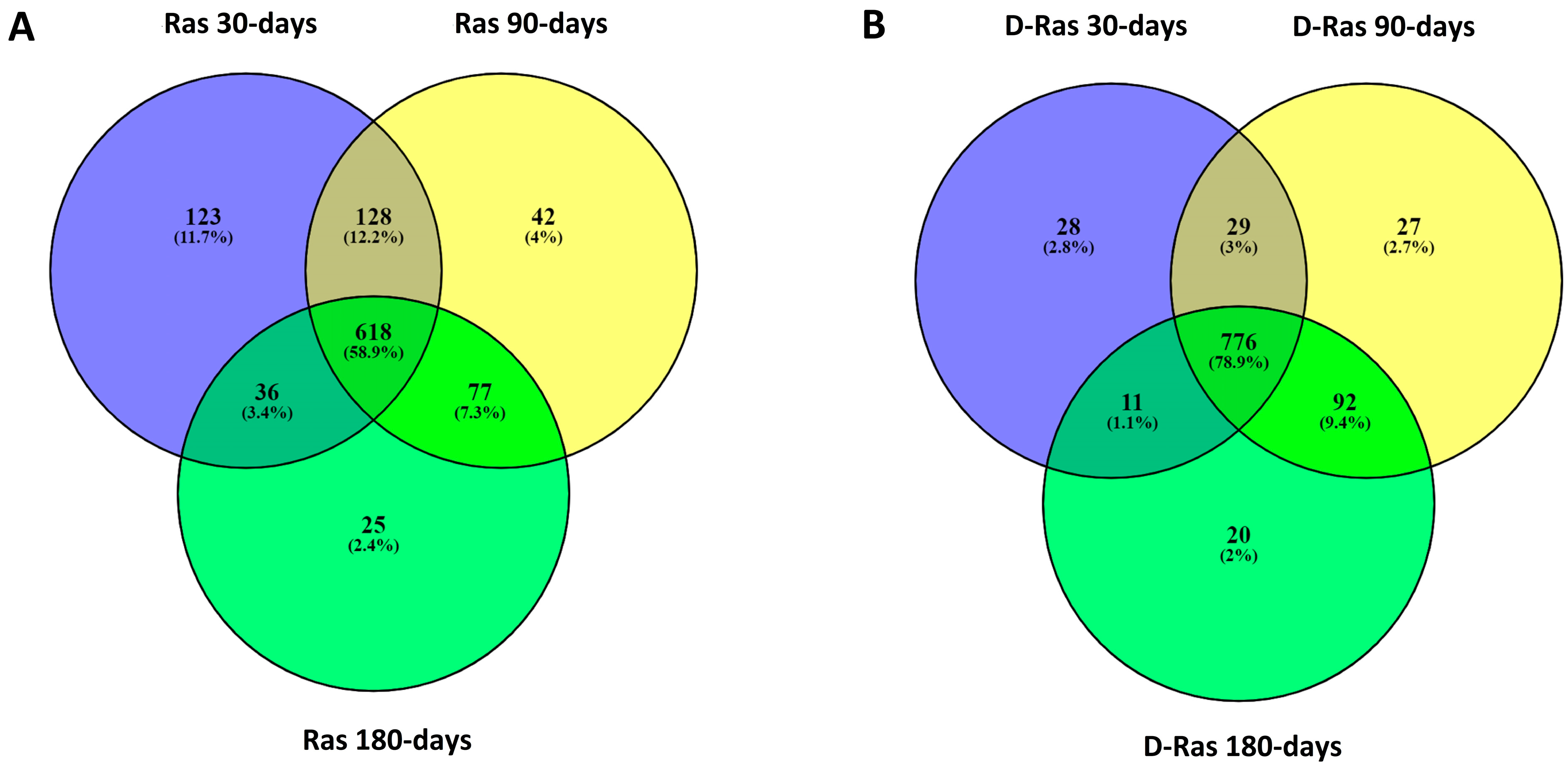
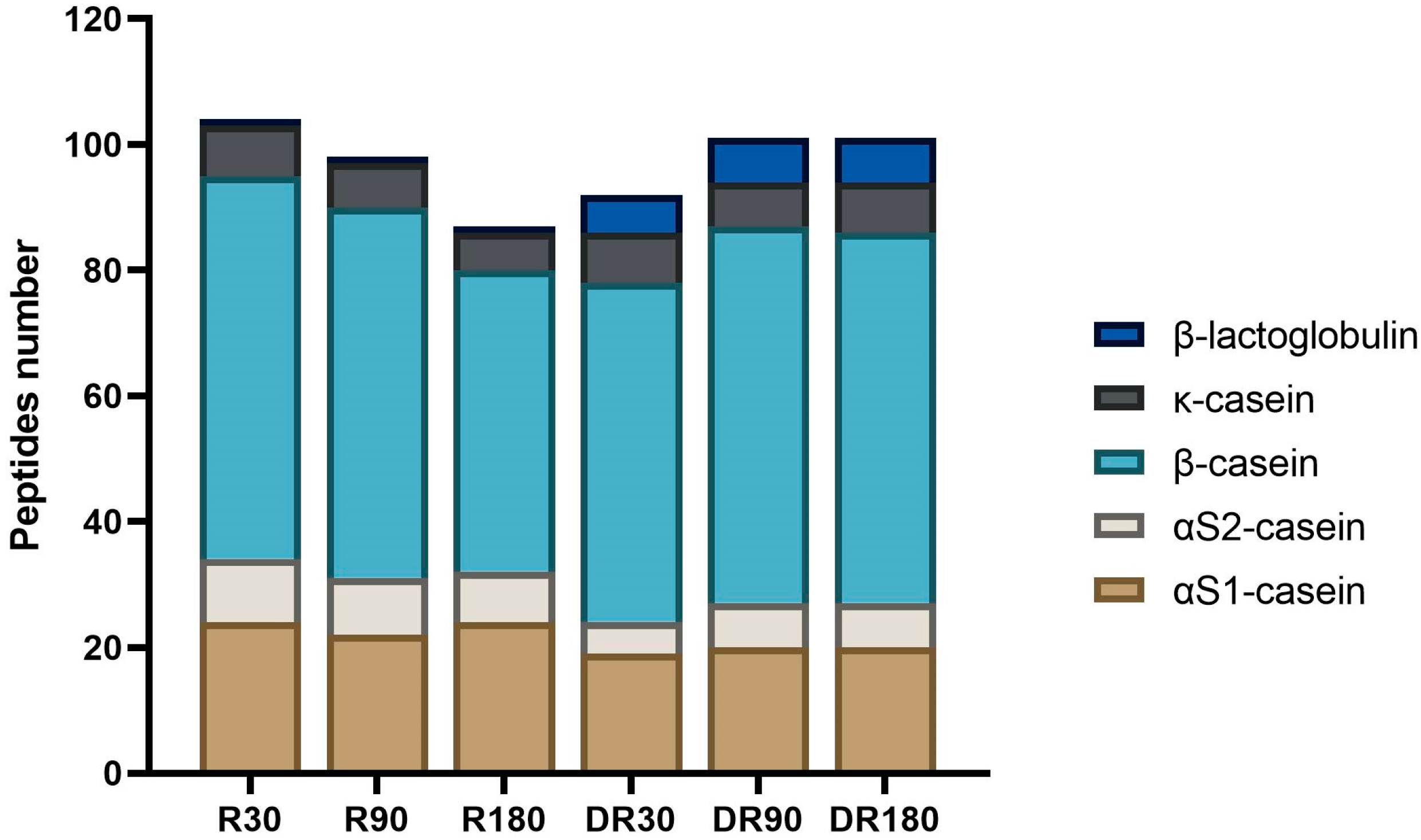
3.3. Peptide Profile Evolution during Ras Cheese In Vitro Digestion
3.4. Identification of Bioactive Peptide in Ras Cheese Samples before and after In Vitro Digestion
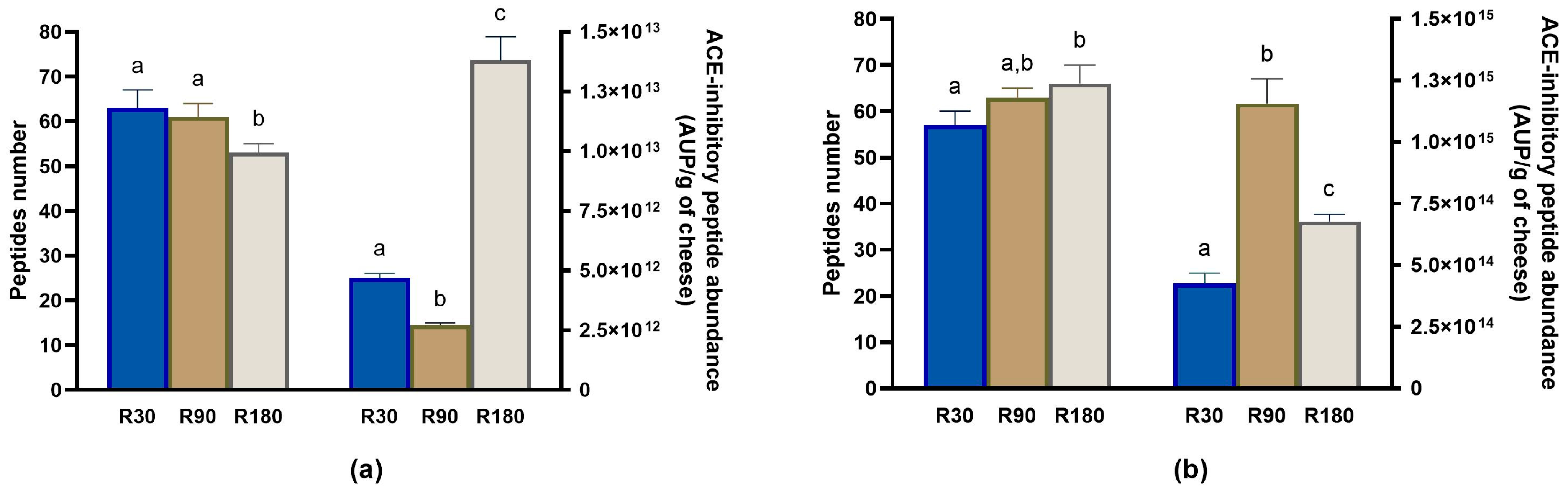
3.4.1. Evolution of ACE-Inhibitory Peptides during Ras Ripening and Digestion
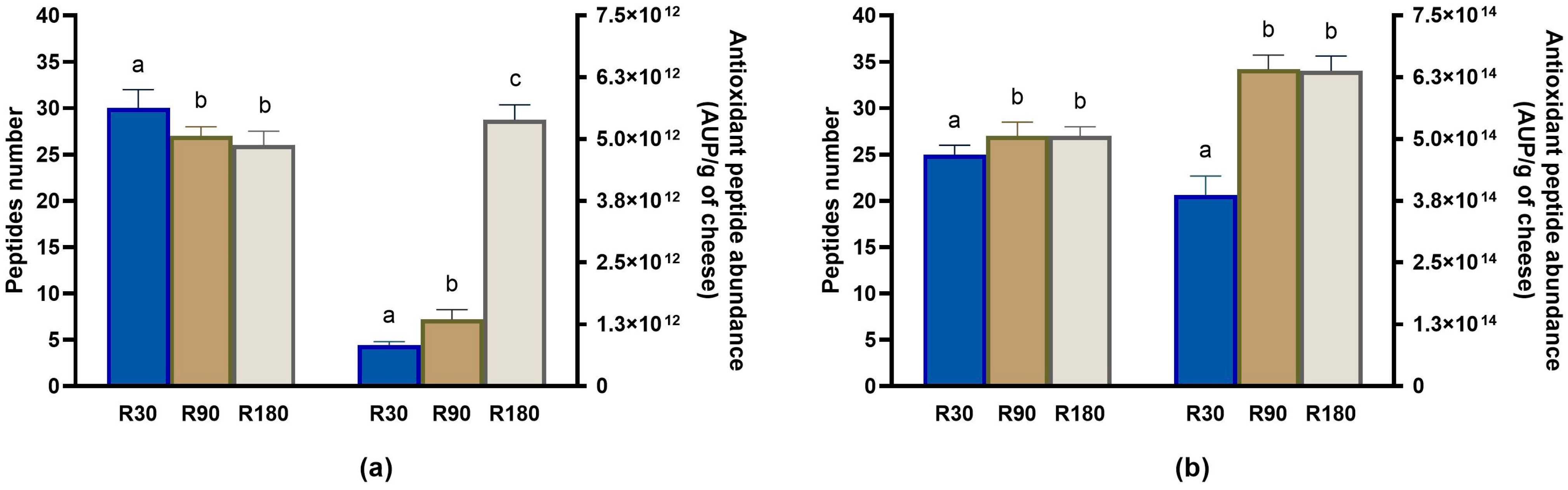
3.4.2. Evolution of Antioxidant Peptides during Ras Ripening and Digestion
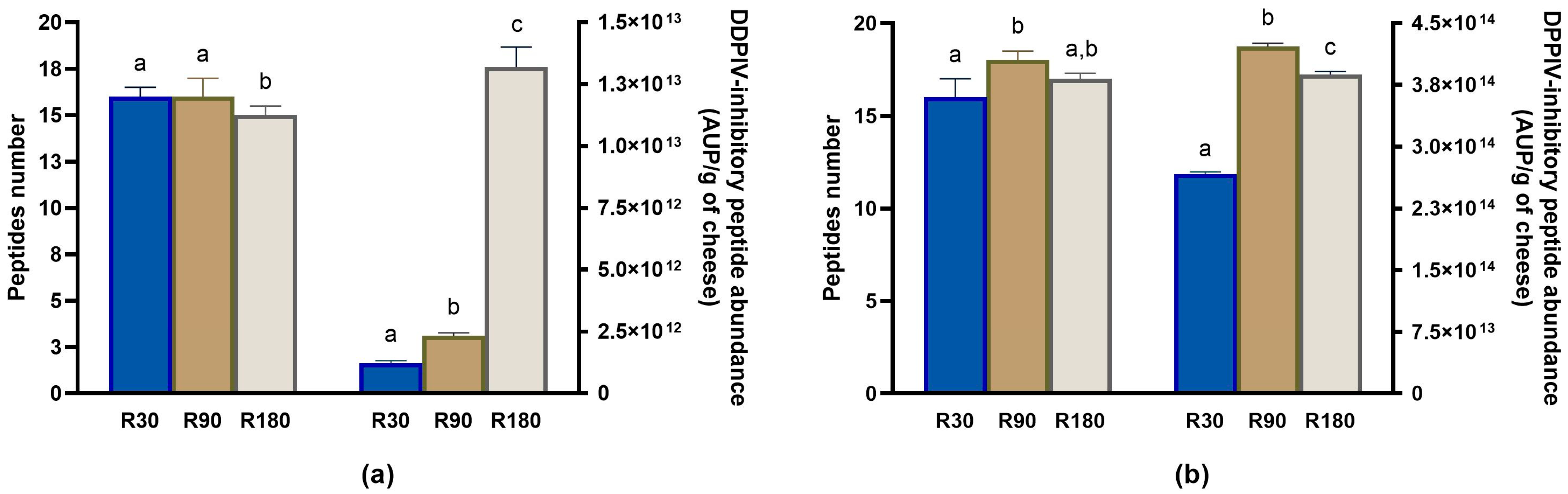
3.4.3. Evolution of DPP-IV-Inhibitory Peptides during Ras Ripening and Digestion
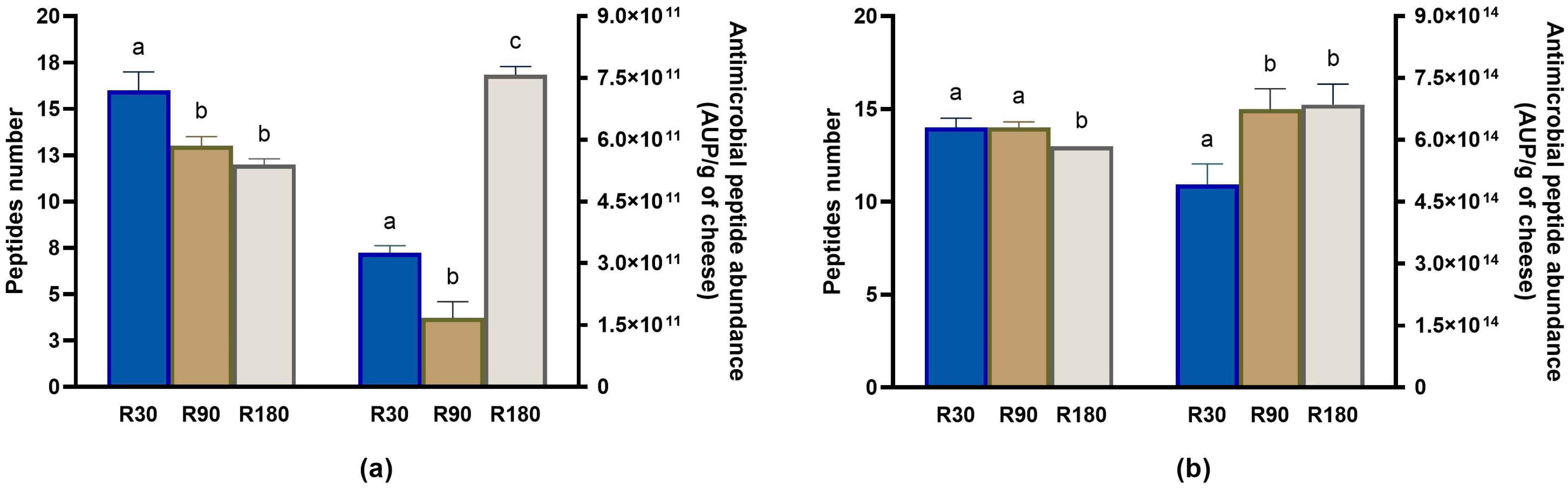
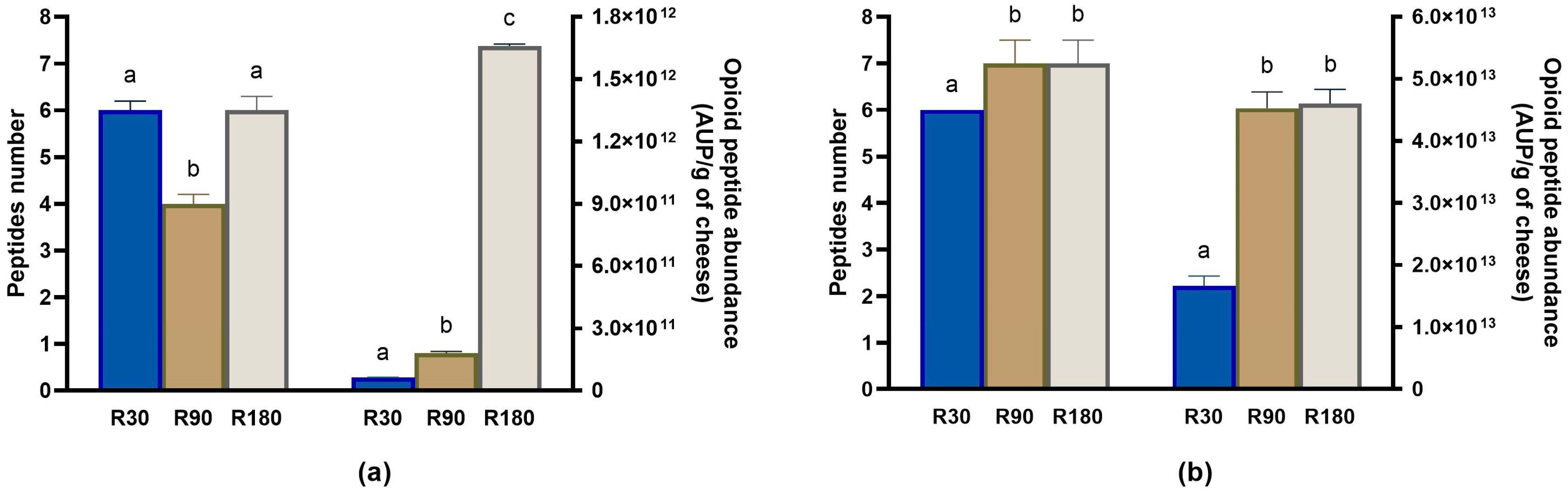
3.4.4. Evolution of Other Bioactive Peptides during Ras Ripening and Digestion
3.5. Quantification of the Bioactive Peptides VPP, IPP and APFPE in Ras Cheese Samples before and after In Vitro Digestion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Awad, S. Texture and flavour development in Ras cheese made from raw and pasteurised milk. Food Chem. 2006, 97, 394–400. [Google Scholar] [CrossRef]
- Abou Donia, S.A. Recent developments in Ras cheese research: A review. Egypt. J. Dairy Sci. 2002, 30, 155–166. [Google Scholar]
- Ayad, E.H.E.; Awad, S.; El Attar, A.; de Jong, C.; El-Soda, M. Characterisation of Egyptian Ras cheese. 2. Flavour formation. Food Chem. 2004, 86, 553–561. [Google Scholar] [CrossRef]
- Phelan, J.A.; Renaud, J.; Fox, P.F. Some Non-European Cheese Varieties. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., Ed.; Springer: Boston, MA, USA, 1993; pp. 421–465. [Google Scholar]
- Álvarez Ramos, L.; Arrieta Baez, D.; Dávila Ortiz, G.; Carlos Ruiz Ruiz, J.; Manuel Toledo López, V. Antioxidant and antihypertensive activity of Gouda cheese at different stages of ripening. Food Chem. X 2022, 14, 100284. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, H.İ.; Akın, N. Effect of ripening time on peptide dynamics and bioactive peptide composition in Tulum cheese. J. Dairy Sci. 2021, 104, 3832–3852. [Google Scholar] [CrossRef] [PubMed]
- Martini, S.; Conte, A.; Tagliazucchi, D. Effect of ripening and in vitro digestion on the evolution and fate of bioactive peptides in Parmigiano-Reggiano cheese. Int. Dairy J. 2020, 105, 104668. [Google Scholar] [CrossRef]
- Castellone, V.; Prandi, B.; Bancalari, E.; Tedeschi, T.; Gatti, M.; Bottari, B. Peptide profile of Parmigiano Reggiano cheese after simulated gastrointestinal digestion: From quality drivers to functional compounds. Front. Microbiol. 2022, 13, 966239. [Google Scholar] [CrossRef]
- Ji, D.; Ma, J.; Xu, M.; Agyei, D. Cell-envelope proteinases from lactic acid bacteria: Biochemical features and biotechnological applications. Compr. Rev. Food Sci. Food Saf. 2021, 20, 369–400. [Google Scholar] [CrossRef]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic systems of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef]
- Christensen, J.E.; Dudley, E.G.; Pederson, J.A.; Steele, J.L. Peptidases and amino acid catabolism in lactic acid bacteria. Antonie Van Leeuwenhoek 1999, 76, 217–246. [Google Scholar] [CrossRef]
- McSweeney, P.L.H. Biochemistry of cheese ripening. Int. J. Dairy Technol. 2004, 57, 127–144. [Google Scholar] [CrossRef]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Martini, S.; Solieri, L.; Cattivelli, A.; Pizzamiglio, V.; Tagliazucchi, D. An integrated peptidomics and in silico approach to identify novel anti-diabetic peptides in Parmigiano-Reggiano cheese. Biology 2021, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Shamsia, S.; Helal, A.; Conte, A. Angiotensin-converting enzyme inhibitory peptides from goats’ milk released by in vitro gastro-intestinal digestion. Int. Dairy J. 2017, 71, 6–16. [Google Scholar] [CrossRef]
- Stuknyte, M.; Cattaneo, S.; Masotti, F.; De Noni, I. Occurrence and fate of ACE-inhibitor peptides in cheeses and in their digestates following in vitro static gastrointestinal digestion. Food Chem 2015, 168, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rivera, L.; Diezhandino, I.; Gómez-Ruiz, J.A.; Fresno, J.M.; Miralles, B.; Recio, I. Peptidomic study of Spanish blue cheese (Valdeón) and changes after simulated gastrointestinal digestion. Electrophoresis 2014, 35, 1627–1636. [Google Scholar] [CrossRef] [PubMed]
- Hofi, A.A.; Youssef, E.H.; Ghoneim, M.A.; Tawab, G.A. Ripening changes in Cephalotyre ‘‘Ras’’ cheese manufactured from raw and pasteurised milk with special reference to flavour. J. Dairy Sci. 1970, 53, 1207–1212. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Kuchroo, C.N.; Fox, P.F. Soluble nitrogen in Cheddar cheese: Comparison of extraction procedures. Milchwissenschaft 1982, 37, 331–335. [Google Scholar]
- IDF. Determination of Chloride Content (Cheese and Processed Cheese); IDF Standard 5B; International Dairy Federation: Brussels, Belgium, 1988. [Google Scholar]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzene sulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Meth. Enzymol. 1999, 299, 15–27. [Google Scholar]
- Solieri, L.; Valentini, M.; Cattivelli, A.; Sola, L.; Helal, A.; Martini, S.; Tagliazucchi, D. Fermentation of whey protein concentrate by Streptococcus thermophilus strains releases peptides with biological activities. Process Biochem. 2022, 121, 590–600. [Google Scholar] [CrossRef]
- Ronca-Testoni, S. Direct spectrophotometric assay for angiotensin-converting enzyme in serum. Clin. Chem. 1983, 29, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Helal, A.; Tagliazucchi, D. Peptidomics profile, bioactive peptides identification and biological activities of six different cheese varieties. Biology 2023, 12, 78. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Martini, S.; Shamsia, S.; Helal, A.; Conte, A. Biological activity and peptidomic profile of in vitro digested cow, camel, goat and sheep milk. Int. Dairy J. 2018, 81, 19–27. [Google Scholar] [CrossRef] [Green Version]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open-source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef] [Green Version]
- Dallas, D.; Nielsen, S.D. Milk peptidomics to identify functional peptides and for quality control of dairy products. Meth. Mol. Biol. 2018, 1719, 223–240. [Google Scholar]
- Nielsen, S.D.; Beverly, R.L.; Qu, Y.; Dallas, D.C. Milk bioactive peptide database: A comprehensive database of milk protein-derived bioactive peptides and novel visualization. Food Chem. 2017, 232, 673–682. [Google Scholar] [CrossRef]
- Bütikofer, U.; Meyer, J.; Sieber, R.; Walther, B.; Wechsler, D. Occurrence of the angiotensin-converting enzyme-inhibiting tripeptides Val-Pro-Pro and Ile-Pro-Pro in different cheese varieties of Swiss origin. J. Dairy Sci. 2008, 91, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Kocak, A.; Sanli, T.; Anli, E.A.; Hayaloglu, A.A. Role of using adjunct cultures in release of bioactive peptides in white-brined goat-milk cheese. LWT Food Sci. Technol. 2020, 123, 109127. [Google Scholar] [CrossRef]
- Baptista, D.P.; da Silva Araújo, F.D.; Eberlin, M.N.; Gigante, M.L. Reduction of 25% salt in Prato cheese does not affect proteolysis and sensory acceptance. Int. Dairy J. 2017, 75, 101–110. [Google Scholar] [CrossRef]
- Galli, B.D.; Nikoloudaki, O.; Tonini, S.; Helal, A.; Di Cagno, R.; Gobbetti, M.; Tagliazucchi, D. How starter cultures affect the peptidomic profile and bioactive activities of the Asiago-PDO cheese throughout ripening. Food Res. Int. 2023, 167, 112743. [Google Scholar] [CrossRef]
- Gaiaschi, A.; Beretta, B.; Poiesi, C.; Conti, A.; Giuffrida, M.G.; Galli, C.L.; Restani, P. Proteolysis of β-casein as a marker of Grana Padano cheese ripening. J. Dairy Sci. 2001, 84, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Ong, L.; Shah, N.P. Release and identification of angiotensin-converting enzyme-inhibitory peptides as influenced by ripening temperatures and probiotic adjuncts in Cheddar cheeses. LWT Food Sci. Technol. 2008, 41, 1555–1566. [Google Scholar] [CrossRef]
- Baptista, D.P.; Negrão, F.; Eberlin, M.N.; Gigante, M.L. Peptide profile and angiotensin-converting enzyme inhibitory activity of Prato cheese with salt reduction and Lactobacillus helveticus as an adjunct culture. Food Res. Int. 2020, 133, 109190. [Google Scholar] [CrossRef]
- Solieri, L.; Sola, L.; Vaccalluzzo, A.; Randazzo, C.L.; Martini, S.; Tagliazucchi, D. Characterization of cell-envelope proteinases from two Lacticaseibacillus casei strains isolated from Parmigiano Reggiano cheese. Biology 2022, 11, 139. [Google Scholar] [CrossRef]
- Lozo, J.; Strahinic, I.; Dalgalarrondo, M.; Chobert, J.-M.; Haertlé, T.; Topisirovic, C. Comparative analysis of β-casein proteolysis by PrtP proteinase from Lactobacillus paracasei subsp. paracasei BGHN14, PrtR proteinase from Lactobacillus rhamnosus BGT10 and PrtH proteinase from Lactobacillus helveticus BGRA43. Int. Dairy J. 2011, 21, 863–868. [Google Scholar] [CrossRef]
- Monnet, V.; Ley, J.P.; Gonzàlez, S. Substrate specificity of the cell envelope-located proteinase of Lactococcus lactis subsp. lactis NCDO 763. Int. J. Biochem. 1992, 24, 707–718. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Gerocarni, B.; Laghi, L.; Borghi, C. Blood pressure lowering effect of lactotripeptides assumed as functional foods: A meta-analysis of current available clinical trials. J. Hum. Hypertens. 2011, 25, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Cicero, A.F.G.; Fogacci, F.; Colletti, A. Potential role of bioactive peptides in prevention and treatment of chronic diseases: A narrative review. Br. J. Pharm. 2017, 174, 1378–1394. [Google Scholar] [CrossRef] [PubMed]
- Fekete, A.A.; Givens, D.I.; Lovegrove, J.A. The impact of milk proteins and peptides on blood pressure and function: A review of evidence from human intervention studies. Nutr. Res. Rev. 2013, 26, 177–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Takano, T. Anti-hypertensive effect of sour milk and peptides isolated from it that are inhibitors to angiotensin I-converting enzyme. J. Dairy. Sci. 1995, 78, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rivera, L.; Ferreira Santos, P.; Sevilla, M.A.; Montero, M.J.; Recio, I.; Miralles, B. Implication of opioid receptors in the antihypertensive effect of a bovine casein hydrolysate and αs1-casein-derived peptides. J. Agric. Food Chem. 2020, 68, 1877–1883. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.; Recio, I.; Ramos, M.; Delgado, M.A.; Aleixandre, M.A. Antihypertensive effect of peptides obtained from Enterococcus faecalis-fermented milk in rats. J. Dairy Sci. 2006, 89, 3352–3359. [Google Scholar] [CrossRef] [Green Version]
- Miguel, M.; Gómez-Ruiz, J.Á.; Recio, I.; Aleixandre, A. Changes in arterial blood pressure after single oral administration of milk-casein-derived peptides in spontaneously hypertensive rats. Mol. Nutr. Food Res. 2010, 54, 1422–1427. [Google Scholar] [CrossRef]
- Maeno, M.; Yamamoto, N.; Takano, T. Identification of an antihypertensive peptide from casein hydrolysate produced by a proteinase from Lactobacillus helveticus CP790. J. Dairy Sci. 1996, 79, 1316–1321. [Google Scholar] [CrossRef]
- Basiricò, L.; Catalani, E.; Morera, P.; Cattaneo, S.; Stuknyte, M.; Bernabucci, U.; De Noni, I.; Nardone, A. Release of angiotensin converting enzyme-inhibitor peptides during in vitro gastrointestinal digestion of Parmigiano Reggiano PDO cheese and their absorption through an in vitro model of intestinal epithelium. J. Dairy Sci. 2015, 98, 7595–7601. [Google Scholar] [CrossRef] [Green Version]
- Caira, S.; Pinto, G.; Picariello, G.; Vitaglione, P.; De Pascale, S.; Scaloni, A.; Addeo, F. In vivo absorptomics: Identification of bovine milk-derived peptides in human plasma after milk intake. Food Chem. 2022, 385, 132663. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Helal, A.; Verzelloni, E.; Conte, A. Bovine milk antioxidant properties: Effect of in vitro digestion and identification of antioxidant compounds. Dairy Sci. Technol. 2016, 96, 657–676. [Google Scholar] [CrossRef] [Green Version]
- Boutrou, R.; Gaudichon, C.; Dupont, D.; Jardin, J.; Airinei, G.; Marsset-Baglieri, A.; Benamouzig, R.; Tomé, D.; Leonil, J. Sequential release of milk protein-derived bioactive peptides in the jejunum in healthy humans. Am. J. Clin. Nutr. 2013, 97, 1314–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beverly, R.L.; Woonnimani, P.; Scottoline, B.P.; Lueangsakulthai, J.; Dallas, D.C. Peptides from the intestinal tract of breast milk-fed infants have antimicrobial and bifidogenic activity. Int. J. Mol. Sci. 2021, 22, 2377. [Google Scholar] [CrossRef] [PubMed]
- Sedaghati, M.; Ezzatpanah, H.; Mashhadi Akbar Boojar, M.; Tajabadi Ebrahimi, M.; Kobarfard, F. Isolation and identification of some antibacterial peptides in the plasmin-digest of β-casein. LWT-Food Sci. Technol. 2016, 68, 217–225. [Google Scholar] [CrossRef]
- Condezo-Hoyos, L.; Noratto, G. Aged American cheddar cheese as source of protein derived compounds that modulate obese mice fecal bacteria and colon inflammation in vitro. Symbiosis 2016, 4, 1–10. [Google Scholar]
- Bottari, B.; Quartieri, A.; Prandi, B.; Raimondi, S.; Leonardi, A.; Rossi, M.; Ulrici, A.; Gatti, M.; Sforza, S.; Nocetti, M.; et al. Characterization of the peptide fraction from digested Parmigiano Reggiano cheese and its effect on growth of lactobacilli and bifidobacteria. Int. J. Food Microbiol. 2017, 255, 32–41. [Google Scholar] [CrossRef]
- Tulipano, G. Role of bioactive peptide sequences in the potential impact of dairy protein intake on metabolic health. Int. J. Mol. Sci. 2020, 21, 8881. [Google Scholar] [CrossRef]
- Aslam, H.; Ruusunen, A.; Berk, M.; Loughman, A.; Rivera, L.; Pasco, J.A.; Jacka, F.N. Unravelled facets of milk derived opioid peptides: A focus on gut physiology, fractures and obesity. Int. J. Food Sci. Nutr. 2020, 71, 36–49. [Google Scholar] [CrossRef]
- Asledottir, T.; Le, T.T.; Poulsen, N.A.; Devold, T.G.; Larsen, L.B.; Vegarud, G.E. Release of β-casomorphin-7 from bovine milk of different β-casein variants after ex vivo gastrointestinal digestion. Int. Dairy J. 2018, 81, 8–11. [Google Scholar] [CrossRef]
- Tyagi, A.; Daliri, E.B.-M.; Kwami Ofosu, F.; Yeon, S.-J.; Oh, D.-H. Food-derived opioid peptides in human health: A review. Int. J. Mol. Sci. 2020, 21, 8825. [Google Scholar] [CrossRef]
- Liu, Z.; Udenigwe, C.C. Role of food-derived opioid peptides in the central nervous and gastrointestinal systems. J. Food Biochem. 2019, 43, e12629. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Govindasamy-Lucey, S.; Lucey, J.A. Angiotensin-I-converting enzyme-inhibitory peptides in commercial Wisconsin Cheddar cheeses of different ages. J. Dairy Sci. 2016, 99, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bütikofer, U.; Meyer, J.; Sieber, R.; Wechsler, D. Quantification of the angiotensin-converting enzyme inhibiting tripeptides Val-Pro-Pro and Ile-Pro-Pro in hard, semi-hard and soft cheeses. Int. Dairy J. 2007, 17, 968–975. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Undigested Ras Cheese | Digested Ras Cheese | |||||
|---|---|---|---|---|---|---|
| Sequence | R30 | R90 | R180 | R30 | R90 | R180 |
| IPP | 2.67 ± 0.11 a | 20.91 ± 1.02 b | 26.77 ± 1.65 c | 6.89 ± 0.41 d | 36.50 ± 2.69 e | 24.76 ± 1.33 c |
| VPP | 0.74 ± 0.02 a | 8.90 ± 0.59 b | 13.13 ± 0.98 c | 2.47 ± 0.19 d | 17.44 ± 1.09 e | 14.69 ± 1.07 c |
| APFPE | 3.27 ± 0.26 a | 9.58 ± 0.77 b | 82.09 ± 5.56 c | 13.59 ± 1.20 d | 17.15 ± 1.54 d | 52.01 ± 3.81 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helal, A.; Cattivelli, A.; Conte, A.; Tagliazucchi, D. Effect of Ripening and In Vitro Digestion on Bioactive Peptides Profile in Ras Cheese and Their Biological Activities. Biology 2023, 12, 948. https://doi.org/10.3390/biology12070948
Helal A, Cattivelli A, Conte A, Tagliazucchi D. Effect of Ripening and In Vitro Digestion on Bioactive Peptides Profile in Ras Cheese and Their Biological Activities. Biology. 2023; 12(7):948. https://doi.org/10.3390/biology12070948
Chicago/Turabian StyleHelal, Ahmed, Alice Cattivelli, Angela Conte, and Davide Tagliazucchi. 2023. "Effect of Ripening and In Vitro Digestion on Bioactive Peptides Profile in Ras Cheese and Their Biological Activities" Biology 12, no. 7: 948. https://doi.org/10.3390/biology12070948