
Molecular Characterization and Expression Pattern of leptin in Yellow Cheek Carp (Elopichthys bambusa) and Its Transcriptional Changes in Response to Fasting and Refeeding
 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Samples
2.2. Cloning of EbLep Gene
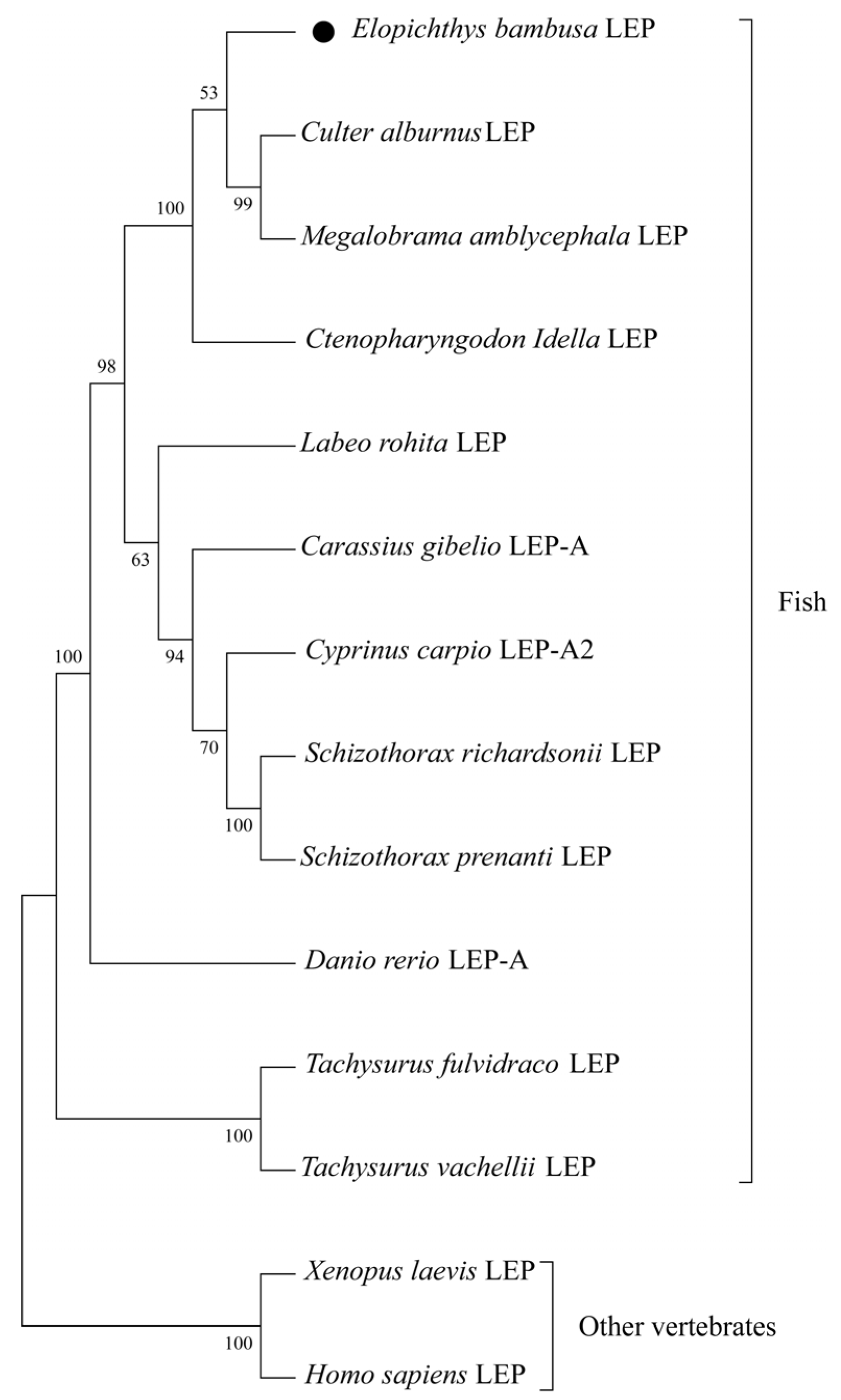
2.3. Sequence Analysis and Phylogenetic Tree Construction
2.4. Tissue Distribution and Fasting–Refeeding Expression
2.5. Data Analysis
3. Results
3.1. Characterization of EbLep
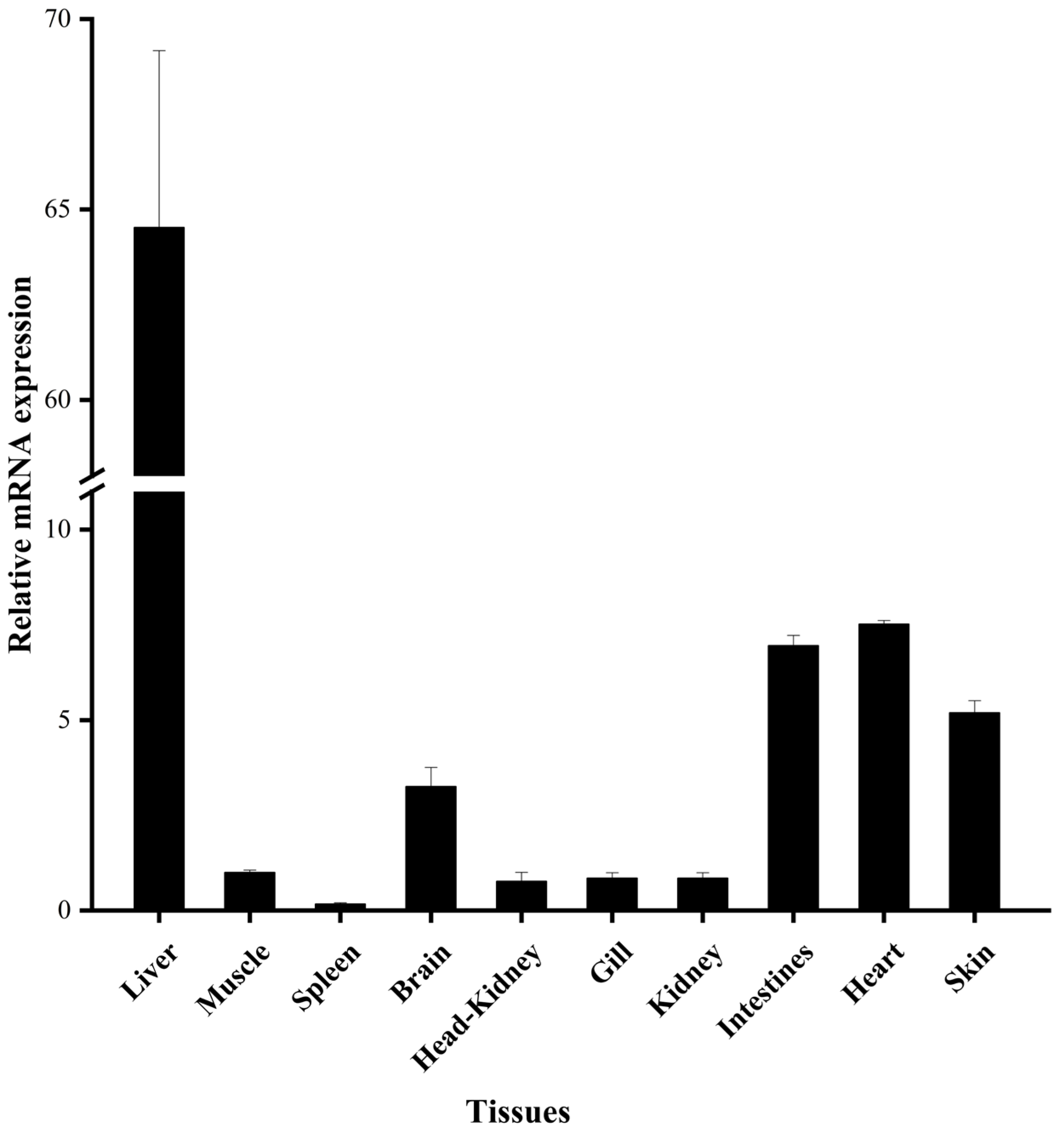
3.2. Tissue Expression of EbLep
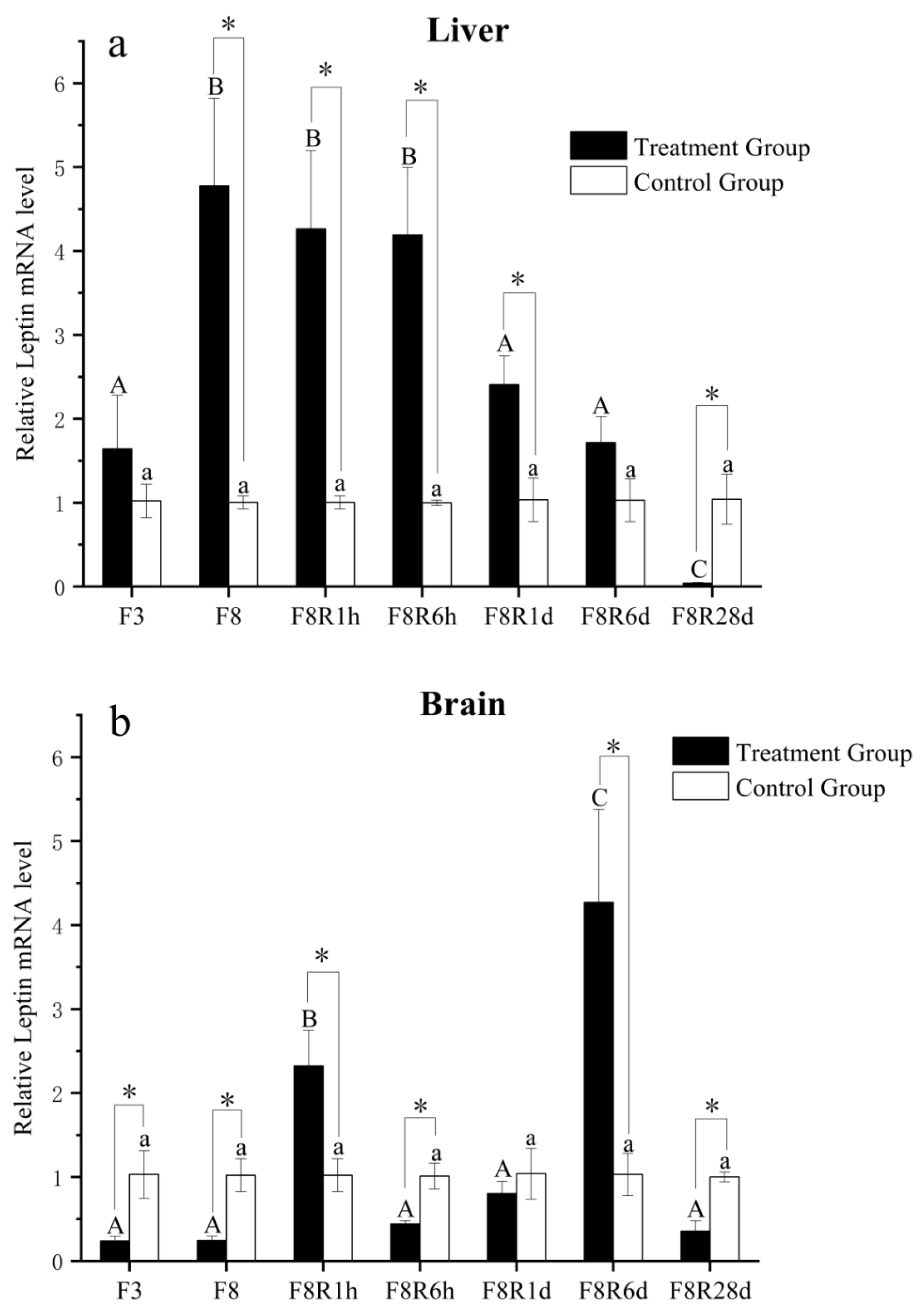
3.3. Expression of EbLep mRNA in the Liver and Brain under Fasting and Refeeding
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Ahima, R.S.; Osei, S.Y. Leptin signaling. Physiol. Behav. 2004, 81, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.W.; Niswender, K.D. Adiposity signaling and biological defense against weight gain: Absence of protection or central hormone resistance. J. Clin. Endocrinol. Metab. 2004, 89, 5889–5897. [Google Scholar] [CrossRef] [PubMed]
- Marty, N.; Dallaporta, M.; Thorens, B. Brain glucose sensing, counter-regulation, and energy homeostasis. Physiology 2007, 22, 241–251. [Google Scholar] [CrossRef]
- Münzberg, H.; Heymsfield, S.B. New insights into the regulation of Leptin gene expression. Cell Metab. 2019, 29, 1013–1014. [Google Scholar] [CrossRef]
- Procaccini, C.; La Rocca, C.; Carbone, F.; De Rosa, V.; Galgani, M.; Matarese, G. Leptin as immune mediator: Interaction between neuroendocrine and immune system. Dev. Comp. Immunol. 2017, 66, 120–129. [Google Scholar] [CrossRef]
- Crespi, E.J.; Denver, R.J. Leptin (ob gene) of the South African clawed frog Xenopus laevis. Proc. Natl. Acad. Sci. USA 2006, 103, 10092–10097. [Google Scholar] [CrossRef]
- Seroussi, E.; Knytl, M.; Pitel, F.; Elleder, D.; Krylov, V.; Leroux, S.; Morisson, M.; Yosefi, S.; Miyara, S.; Ganesan, S.; et al. Avian Expression Patterns and Genomic Mapping Implicate Leptin in Digestion and TNF in Immunity, Suggesting That Their Interacting Adipokine Role Has Been Acquired Only in Mammals. Int. J. Mol. Sci. 2019, 20, 4489. [Google Scholar] [CrossRef]
- Wang, A.Z.; Husak, J.F.; Lovern, M. Leptin ameliorates the immunity, but not reproduction, trade-off with endurance in lizards. J. Comp. Physiol. B 2019, 189, 261–269. [Google Scholar] [CrossRef]
- Sahu, A. Minireview: A hypothalamic role in energy balance with special emphasis on leptin. Endocrinology 2004, 145, 2613–2620. [Google Scholar] [CrossRef]
- Kurokawa, T.; Uji, S.; Suzuki, T. Identification of cDNA coding for a homologue to mammalian leptin from pufferfish, Takifugu rubripes. Peptides 2005, 26, 745–750. [Google Scholar] [CrossRef]
- Li, G.G.; Liang, X.F.; Xie, Q.; Li, G.; Yu, Y.; Lai, K. Gene structure, recombinant expression and functional characterization of grass carp leptin. Gen. Comp. Endocrinol. 2010, 166, 117–127. [Google Scholar] [CrossRef]
- Tang, Y.; Yu, J.; Li, H.; Xu, P.; Li, J.; Ren, H. Molecular cloning, characterization and expression analysis of multiple leptin genes in Jian carp (Cyprinus carpio var. Jian). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 166, 133–140. [Google Scholar] [CrossRef]
- Gorissen, M.; Bernier, N.J.; Nabuurs, S.B.; Flik, G.; Huising, M.O. Two divergent leptin paralogues in zebrafish (Danio rerio) that originate early in teleostean evolution. J. Endocrinol. 2009, 201, 329–339. [Google Scholar] [CrossRef]
- Yuan, X.; Li, A.; Liang, X.-F.; Huang, W.; Song, Y.; He, S.; Tao, Y. Leptin expression in mandarin fish Siniperca chuatsi (Basilewsky): Regulation by postprandial and short-term fasting treatment. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 194, 8–18. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, H.; Zhang, Y.; Li, S.; Lu, D.; Zhang, H.; Meng, Z.; Liu, X.; Lin, H. Molecular cloning, characterization and expression profiles of multiple leptin genes and a leptin receptor gene in orange-spotted grouper (Epinephelus coioides). Gen. Comp. Endocrinol. 2013, 181, 295–305. [Google Scholar] [CrossRef]
- Murashita, K.; Uji, S.; Yamamoto, T.; Rønnestad, I.; Kurokawa, T. Production of recombinant leptin and its effects on food intake in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 150, 377–384. [Google Scholar] [CrossRef]
- Yuan, X.-C.; Liang, X.-F.; Cai, W.-J.; Li, A.-X.; Huang, D.; He, S. Differential roles of two leptin gene paralogues on food intake and hepatic metabolism regulation in Mandarin fish. Front. Endocrinol. 2020, 11, 438. [Google Scholar] [CrossRef]
- Tsakoumis, E.; Ahi, E.P.; Schmitz, M. Impaired leptin signaling causes subfertility in female zebrafish. Mol. Cell. Endocrinol. 2022, 546, 111595. [Google Scholar] [CrossRef]
- Mohammadabadi, M.; Kord, M.; Nazari, M. Studying expression of leptin gene in different tissues of Kermani Sheep using Real Time PCR. Agric. Biotechnol. J. 2018, 10, 111–123. [Google Scholar]
- Ayelén, M.B.; José, L.S. Leptin signalling in teleost fish with emphasis in food intake regulation. Mol. Cell. Endocrinol. 2021, 526, 111209. [Google Scholar] [CrossRef]
- Martin, S.A.M.; Douglas, A.; Houlihan, D.F.; Secombes, C.J. Starvation alters the liver transcriptome of the innate immune response in Atlantic salmon (Salmo salar). BMC Genom. 2010, 11, 418. [Google Scholar] [CrossRef] [PubMed]
- Power, D.M.; Melo, J.; Santos, C.R.A. The effect of food deprivation on the liver, tyroid hormones and transthyretin in sea bream. J. Fish Biol. 2000, 56, 374–387. [Google Scholar] [CrossRef]
- Gambardella, C.; Gallus, L.; Amaroli, A.; Terova, G.; Masini, M.A.; Ferrando, S. Fasting and re-feeding impact on leptin and aquaglyceroporin 9 in the liver of European sea bass (Dicentrarchus labrax). Aquaculture 2012, 354–355, 1–6. [Google Scholar] [CrossRef]
- Won, E.T.; Baltzegar, D.A.; Picha, M.E. Cloning and characterization of leptin in a Perciform fish, the striped bass (Morone saxatilis): Control of feeding and regulation by nutritional state. Gen. Comp. Endocrinol. 2012, 178, 98–107. [Google Scholar] [CrossRef]
- Delgado, M.J.; Cerdá-Reverter, J.M.; Soengas, J.L. Hypothalamic integration of metabolic, endocrine, and circadian signals in fish: Involvement in the control of food intake. Front. Neurosci. 2017, 11, 354. [Google Scholar] [CrossRef]
- Soengas, J.L.; Cerda-Reverter, J.M.; Delgado, M.J. Central regulation of food intake in fishes: An evolutionary perspective. J. Mol. Endocrinol. 2018, 60, R171–R199. [Google Scholar] [CrossRef]
- Wan, S.L.; Wang, L.; Li, J.; Li, J.Z.; Liu, H.J.; Cai, C.M.; Lu, L.L.; Wen, Z.R. Flesh content and nutritive composition of yellow cheek carp (Elopichthys bambusa). Freshw. Fish. 2008, 273, 27–29. [Google Scholar] [CrossRef]
- Liang, Y.; Melack, J.M.; Wang, J. Primary production and fish yields in Chinese ponds and lakes. Trans. Am. Fish. Soc. 1981, 110, 346–350. [Google Scholar] [CrossRef]
- Chen, Z.; Dong, S.; Dai, L.; Xie, M.; Fu, W.; Yuan, X.; Yuan, S.; Liu, J.; Peng, L.; Li, S.; et al. Effect of food domestication on the growth of Elopichthys bambusa. Reprod. Breed. 2021, 1, 157–166. [Google Scholar] [CrossRef]
- Johnson, R.M.; Johnson, T.M.; Londraville, R.L. Evidence for leptin expression in fishes. J. Exp. Zool. 2000, 286, 718–724. [Google Scholar] [CrossRef]
- Wen, Z.-Y.; Qin, C.-J.; Wang, J.; He, Y.; Li, H.-T.; Li, R.; Wang, X.-D. Molecular characterization of two leptin genes and their transcriptional changes in response to fasting and refeeding in Northern snakehead (Channa argus). Gene 2020, 736, 144420. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, Y.; Wang, B.; Liu, X.; Liu, Q.; Song, X.; Shi, B.; Ren, K. Leptin and leptin receptor genes in tongue sole (Cynoglossus semilaevis): Molecular cloning, tissue distribution and differential regulation of these genes by sex steroids. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 224, 11–22. [Google Scholar] [CrossRef]
- Kurokawa, T.; Murashita, K. Genomic characterization of multiple leptin genes and a leptin receptor gene in the Japanese medaka, Oryzias latipes. Gen. Comp. Endocrinol. 2009, 161, 229–237. [Google Scholar] [CrossRef]
- Glasauer, S.M.K.; Neuhauss, S.C.F. Whole-genome duplication in teleost fishes and its evolutionary consequences. Mol. Genet. Genom. 2014, 289, 1045–1060. [Google Scholar] [CrossRef]
- Deck, C.A.; Honeycutt, J.L.; Cheung, E.; Reynolds, H.M.; Borski, R.J. Assessing the functional role of leptin in energy homeostasis and the stress response in vertebrates. Front. Endocrinol. 2017, 8, 63. [Google Scholar] [CrossRef]
- Yan, A.; Li, J.; Liu, L.; Zhu, X.; Ren, C.; Hu, C.; Tang, D.; Chen, T. Tetraploid genes of leptin (leptin-AI,-AII,-BI and-BII) in goldfish: Molecular cloning, bioinformatics analysis, tissue distribution and differential regulation of transcript expression by glucocorticoids. Aquac. Rep. 2022, 25, 101191. [Google Scholar] [CrossRef]
- Huising, M.O.; Geven EJ, W.; Kruiswijk, C.P.; Nabuurs, S.B.; Stolte, E.H.; Spanings FA, T.; Kemenade, L.V.; Flik, G. Increased leptin expression in common carp (Cyprinus carpio) after food intake but not after fasting or feeding to satiation. Endocrinology 2006, 147, 5786–5797. [Google Scholar] [CrossRef]
- Rønnestad, I.; Nilsen, T.O.; Murashita, K.; Angotzi, A.R.; Gamst Moen, A.-G.; Stefansson, S.O.; Kurokawa, T. Leptin and leptin receptor genes in Atlantic salmon: Cloning, phylogeny, tissue distribution and expression correlated to long-term feeding status. Gen. Comp. Endocrinol. 2010, 168, 55–70. [Google Scholar] [CrossRef]
- Bartha, T.; Sayed-Ahmed, A.; Rudas, P. Expression of leptin and its receptors in various tissues of ruminants. Domest. Anim. Endocrinol. 2005, 29, 193–202. [Google Scholar] [CrossRef]
- Zhao, H.H.; Li, X.C.; Zeng CWang, W.M.; Dong, Z.J.; Gao, Z.X. Expression analysis of leptin genes in adults’ tissue and during early development in Megalobrama amblycephala. J. Huazhong Agric. Univ. 2016, 35, 92–98. [Google Scholar] [CrossRef]
- Kling, P.; Rønnestad, I.; Stefansson, S.O.; Murashita, K.; Kurokawa, T.; Björnsson, B.T.A. homologous salmonid leptin radioimmunoassay indicates elevated plasma leptin levels during fasting of rainbow trout. Gen. Comp. Endocrinol. 2009, 62, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.S.; Miao, L.; Li, M.Y.; Hou, H.H.; Li, X.M.; Xu, Y.M. Cloning and expression of leptin gene in Acrossocheilus fasciatus during fasting and refeeding. Oceanol. Limnol. Sin. 2017, 48, 822–829. [Google Scholar]
- Aguilar, A.J.; Conde-Sieira, M.; Polakof, S.; Míguez, J.M.; Soengas, J.L. Central leptin treatment modulates brain glucosensing function and peripheral energy metabolism of rainbow trout. Peptides 2010, 31, 1044–1054. [Google Scholar] [CrossRef]
- Jeong, I.; Kim, E.; Kim, S.; Kim, H.; Lee, D.W.; Seong, J.Y.; Park, H.C. mRNA expression and metabolic regulation of npy and agrp1/2 in the zebrafish brain. Neurosci. Lett. 2018, 668, 73–79. [Google Scholar] [CrossRef]
- Ahi, E.P.; Tsakoumis, E.; Brunel, M.; Schmitz, M. Transcriptional study reveals a potential leptin-dependent gene regulatory network in zebrafish brain. Fish Physiol. Biochem. 2021, 47, 1283–1298. [Google Scholar] [CrossRef]
- Volkof, H. The neuroendocrine regulation of food intake in fish: A review of current knowledge. Front. Neurosci. 2016, 10, 540. [Google Scholar] [CrossRef]
- Zhan, X.M.; Li, Y.L.; Wang, D.H. Effects of fasting and refeeding on body mass, thermogenesis, and serum leptin in Brandt’s voles (Lasiopodomys brandtii). J. Therm. Biol. 2009, 34, 237–243. [Google Scholar] [CrossRef]
- Chelikani, P.K.; Ambrose, J.D.; Keisler, D.H.; Kennelly, J.J. Effect of short-term fasting on plasma concentrations of leptin and other hormones and metabolites in dairy cattle. Domest. Anim. Endocrinol. 2004, 26, 33–48. [Google Scholar] [CrossRef]
- Yuan, D.; Wang, T.; Zhou, C.; Lin, F.; Chen, H.; Wu, H.; Wei, R.; Xin, Z.; Li, Z. Leptin and cholecystokinin in Schizothorax prenanti: Molecular cloning, tissue expression, and mRNA expression responses to periprandial changes and fasting. Gen. Comp. Endocrinol. 2014, 204, 13–24. [Google Scholar] [CrossRef]
- Thornton, J.E.; Cheung, C.C.; Clifton, D.K.; Steiner, R.A. Regulation of hypothalamic proopiomelanocortin mRNA by leptin in ob/ob mice. Endocrinology 1997, 138, 5063–5066. [Google Scholar] [CrossRef]
- Ghamari-Langroudi, M.; Srisai, D.; Cone, R. Multinodal regulation of the arcuate/paraventricular nucleus circuit by leptin. Proc. Natl. Acad. Sci. USA 2011, 108, 355–360. [Google Scholar] [CrossRef]
- Lee, S.J.; Verma, S.; Simonds, S.E.; Kirigiti, M.A.; Kievit, P.; Lindsley, S.R.; Loche, A.; Smith, M.S.; Cowley, M.A.; Grove, K.L. Leptin stimulates neuropeptide Y and cocaine amphetamine-regulated transcript co-expressing neuronal activity in the dorsomedial hypothalamus in diet-induced obese mice. J. Neurosci. 2013, 33, 15306–15317. [Google Scholar] [CrossRef]
- Xie, M.; Xiang, J.G.; Guo, X.F. Measurement of empty-feeding rate of gut in yellow cheek carp (Elopichthys bambusa). Sci. Fish Farming 2015, 10, 50. [Google Scholar]
- López, M.; Tovar, S.; Vazquez, M.J.; Nogueiras, R.; Senaris, R.; Dieguez, C. Sensing the fat: Fatty acid metabolism in the hypothalamus and the melanocortin system. Peptides 2005, 26, 1753–1758. [Google Scholar] [CrossRef]
- Fuentes, E.N.; Kling, P.; Einarsdottir, I.E.; Alvarez, M.; Valdés, J.A.; Molina, A.; Björnsson, B.T. Plasma leptin and growth hormone levels in the fine flounder (Paralichthys adspersus) increase gradually during fasting and decline rapidly after refeeding. Gen. Comp. Endocrinol. 2012, 177, 120–127. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purpose | Name | Sequence (5′-3′) | Annealing Temperature | E-Values |
|---|---|---|---|---|
| Core Fragment Sequence Acquisition | LEP-1 | F: ATGTATTYTCCAGYTCTTC | 55 °C | |
| R: GCATGAACTSTKTCAGTC | ||||
| 5′-RACE PCR | LEP-5′-RACE-1 | ATGATGGTGTCTGCCT | ||
| LEP-5′-RACE-2 | TGAGGCTATCCTGATG | |||
| LEP-5′-RACE-3 | CCATCAATCAGACCAAGAAT | |||
| 3′-RACE PCR | LEP-3′-RACE-1 | CCAAGGGGCATGTGAGCCAGTTAC | ||
| LEP-3′-RACE-2 | GCACACCAAAGGAGCCAGCCAATG | |||
| Expression Examination | q-leptin | F: ACTGTCTCCAAAGATCCTCA | 60 °C | 100.3% |
| R: AAAAGGGTGGACACATCATT | ||||
| Reference Genes | β-actin | F: CCTGTATGCCTCTGGTCG | 60 °C | 98.6% |
| R: CTCGGCTGTGGTGGTGAA |
| Eb LEP | Ci LEP | Ca LEP | Ma LEP | Cg LEP-A | CcLEP-A2 | Lr LEP | Sp LEP | Sr LEP | Dr LEP-A | Tf LEP | Tv LEP | Xl LEP | Hs LEP | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Eb LEP | 100.00% | |||||||||||||
| Ci LEP | 91.60% | 100.00% | ||||||||||||
| Ca LEP | 91.60% | 89.70% | 100.00% | |||||||||||
| Ma LEP | 91.00% | 89.70% | 96.10% | 100.00% | ||||||||||
| Cg LEP-A | 76.10% | 75.50% | 72.90% | 73.50% | 100.00% | |||||||||
| Cc LEP-A2 | 76.80% | 76.80% | 74.20% | 74.20% | 87.10% | 100.00% | ||||||||
| Lr LEP | 74.80% | 73.40% | 72.90% | 71.60% | 78.80% | 77.40% | 100.00% | |||||||
| Sp LEP | 74.20% | 74.20% | 72.30% | 73.50% | 83.20% | 86.50% | 75.50% | 100.00% | ||||||
| Sr LEP | 74.20% | 72.90% | 72.30% | 73.50% | 83.20% | 86.50% | 75.50% | 97.40% | 100.00% | |||||
| Dr LEP-A | 60.00% | 59.40% | 58.70% | 59.40% | 63.90% | 65.20% | 63.90% | 61.90% | 61.90% | 100.00% | ||||
| Tf LEP | 40.00% | 40.00% | 40.00% | 40.60% | 38.70% | 41.30% | 37.40% | 40.60% | 41.30% | 36.80% | 100.00% | |||
| Tv LEP | 40.00% | 40.00% | 40.00% | 40.60% | 39.40% | 41.90% | 38.10% | 41.30% | 41.90% | 37.40% | 99.40% | 100.00% | ||
| Xl LEP | 27.70% | 30.30% | 28.40% | 28.40% | 28.40% | 26.50% | 27.10% | 27.10% | 26.50% | 27.70% | 29.00% | 29.00% | 100.00% | |
| Hs LEP | 23.20% | 22.60% | 21.30% | 21.30% | 22.60% | 25.20% | 23.90% | 25.20% | 23.90% | 23.20% | 21.30% | 21.30% | 35.50% | 100.00% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, M.; Gao, J.; Wu, H.; Cheng, X.; Zhang, Z.; Song, R.; Li, S.; Zhou, J.; Li, C.; Zeng, G. Molecular Characterization and Expression Pattern of leptin in Yellow Cheek Carp (Elopichthys bambusa) and Its Transcriptional Changes in Response to Fasting and Refeeding. Biology 2023, 12, 758. https://doi.org/10.3390/biology12050758
Xie M, Gao J, Wu H, Cheng X, Zhang Z, Song R, Li S, Zhou J, Li C, Zeng G. Molecular Characterization and Expression Pattern of leptin in Yellow Cheek Carp (Elopichthys bambusa) and Its Transcriptional Changes in Response to Fasting and Refeeding. Biology. 2023; 12(5):758. https://doi.org/10.3390/biology12050758
Chicago/Turabian StyleXie, Min, Jinwei Gao, Hao Wu, Xiaofei Cheng, Zhou Zhang, Rui Song, Shaoming Li, Jie Zhou, Cheng Li, and Guoqing Zeng. 2023. "Molecular Characterization and Expression Pattern of leptin in Yellow Cheek Carp (Elopichthys bambusa) and Its Transcriptional Changes in Response to Fasting and Refeeding" Biology 12, no. 5: 758. https://doi.org/10.3390/biology12050758