
Tumor Microenvironment and Glioblastoma Cell Interplay as Promoters of Therapeutic Resistance

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
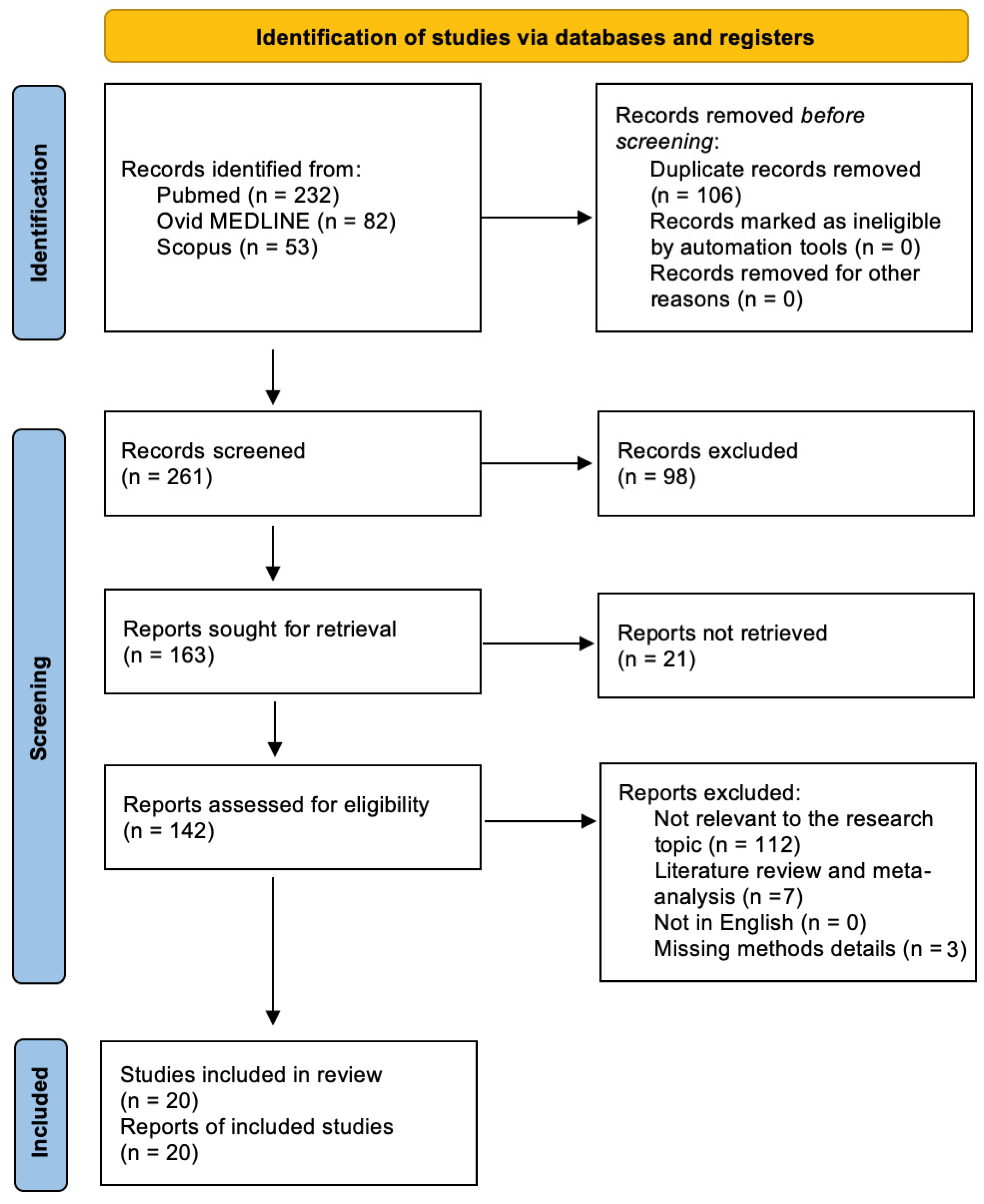
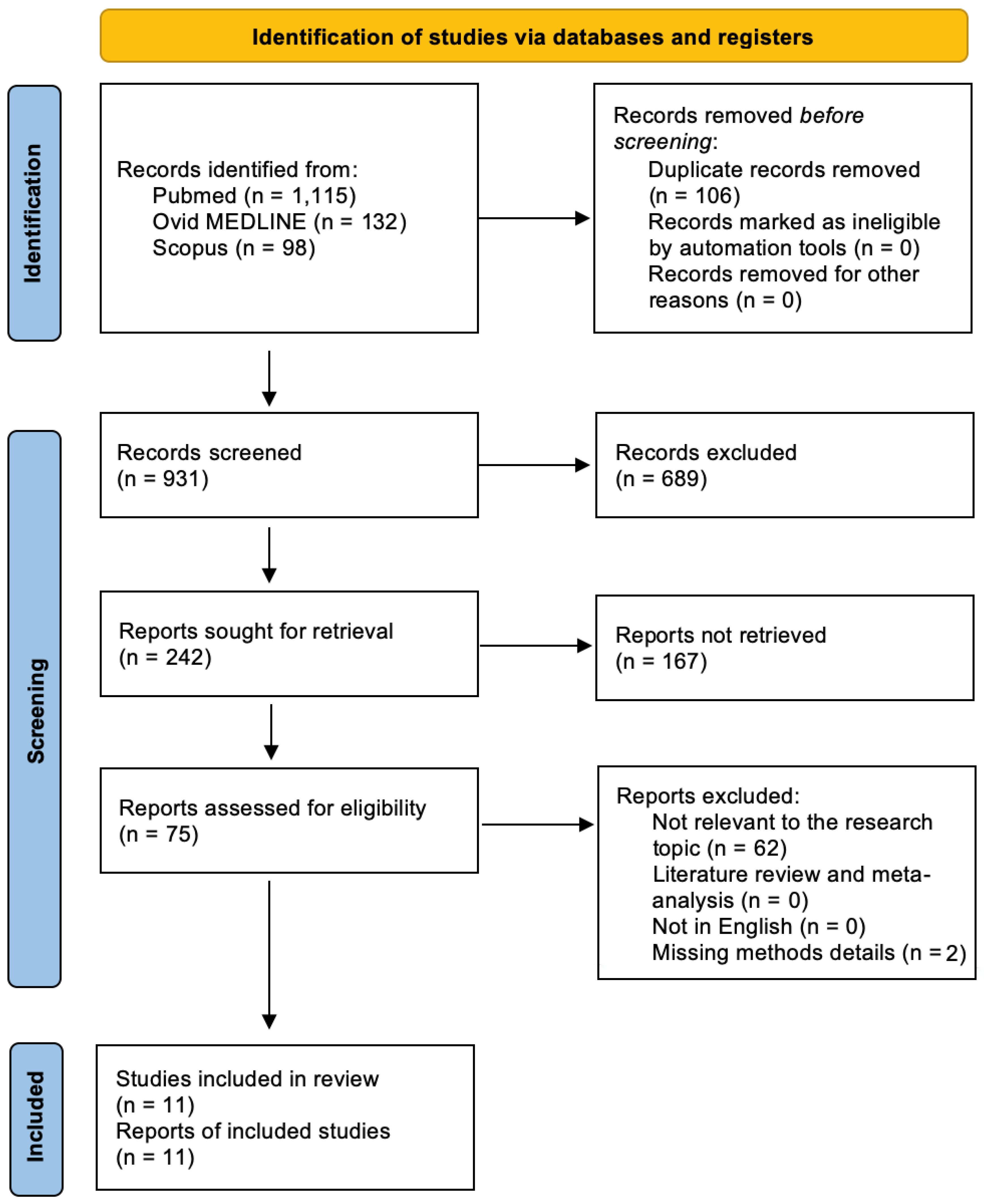
3.1. Literature Search
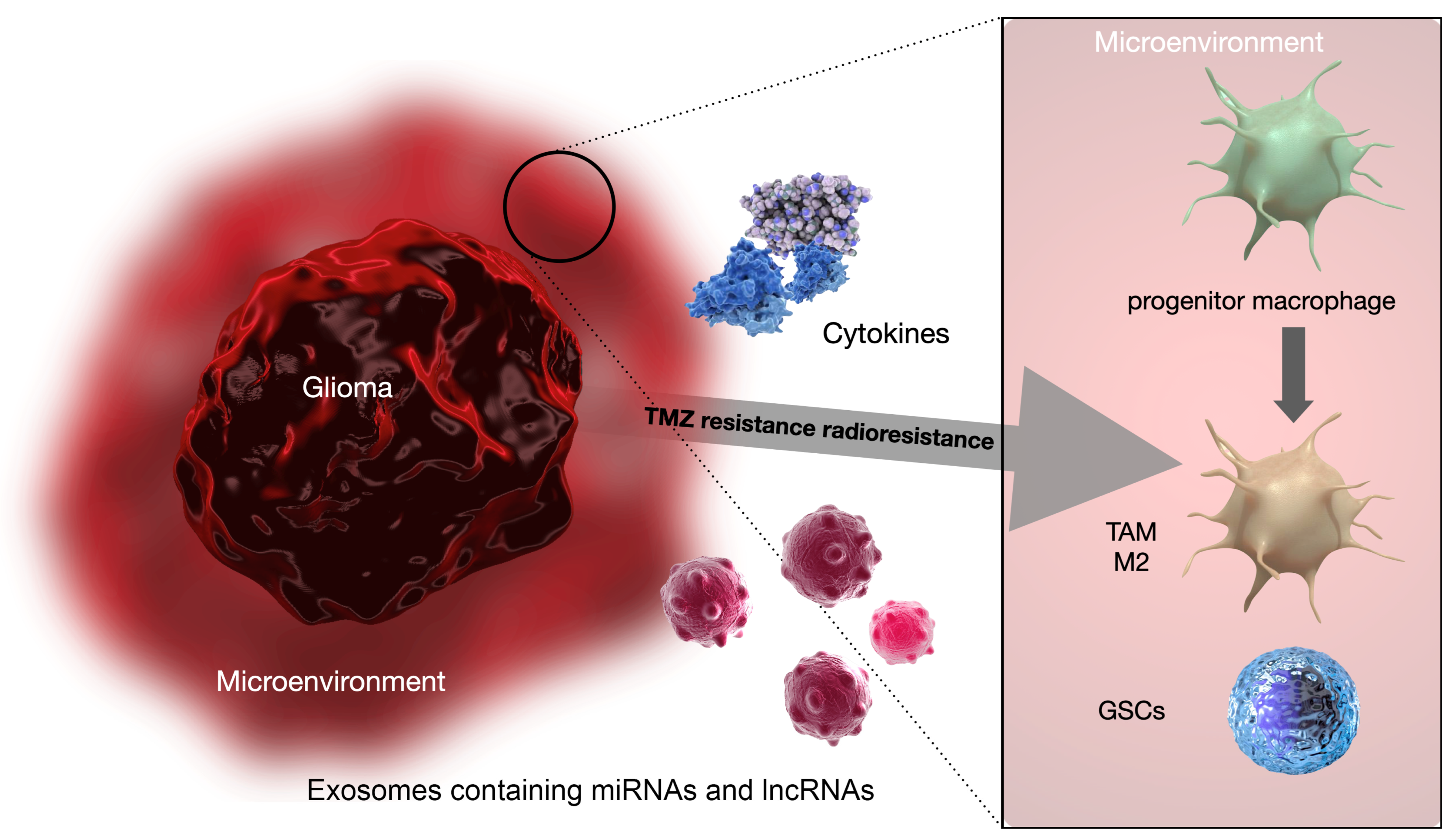
TME and Its Role in Radioresistance and Chemoresistance
3.2. Included Studies
- Exosomal non-coding RNAs, including both micro RNAs (miRNAs) and long non-coding RNAs (lncRNAs);
- M2 macrophage polarization.
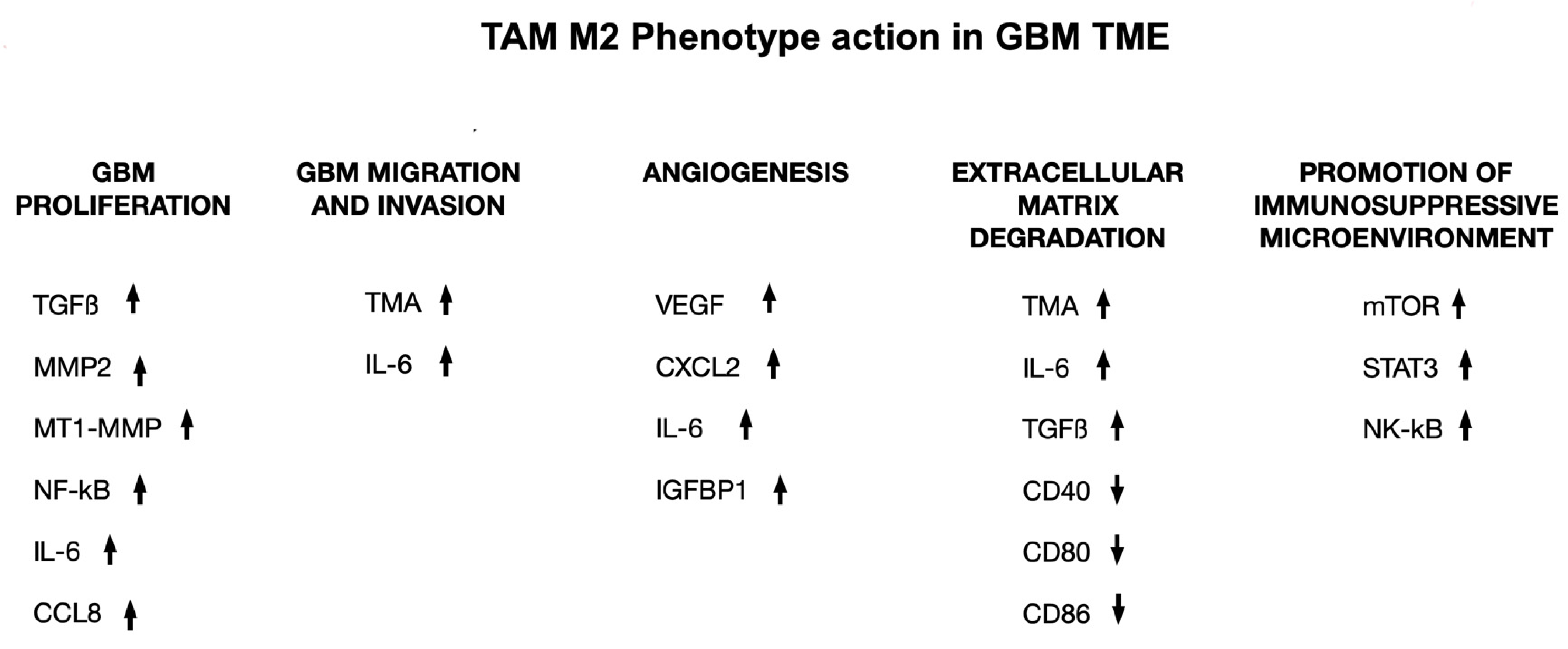
3.3. TME and Immunotherapeutic Strategies
4. Discussion
Future Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. European Organisation for Research and Treatment of Cancer Brain Tumor and Radiotherapy Groups, National Cancer Institute of Canada Clinical Trials Group. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Ius, T.; Pignotti, F.; Della Pepa, G.M.; La Rocca, G.; Somma, T.; Isola, M.; Battistella, C.; Gaudino, S.; Polano, M.; Dal Bo, M.; et al. A Novel Comprehensive Clinical Stratification Model to Refine Prognosis of Glioblastoma Patients Undergoing Surgical Resection. Cancers 2020, 12, 386. [Google Scholar] [CrossRef] [PubMed]
- Menna, G.; Mattogno, P.P.; Donzelli, C.M.; Lisi, L.; Olivi, A.; Della Pepa, G.M. Glioma-Associated Microglia Characterization in the Glioblastoma Microenvironment through A ‘Seed-and Soil’ Approach: A Systematic Review. Brain Sci. 2022, 12, 718. [Google Scholar] [CrossRef]
- Ius, T.; Somma, T.; Altieri, R.; Angileri, F.F.; Barbagallo, G.M.; Cappabianca, P.; Certo, F.; Cofano, F.; D’Elia, A.; Della Pepa, G.M.; et al. Is age an additional factor in the treatment of elderly patients with glioblastoma? A new stratification model: An Italian Multicenter Study. Neurosurg. Focus 2020, 49, E13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Tao, X.; Ji, B.; Gong, J. Hypoxia-Driven M2-Polarized Macrophages Facilitate Cancer Aggressiveness and Temozolomide Resistance in Glioblastoma. Oxid. Med. Cell. Longev. 2022, 2022, 1614336. [Google Scholar] [CrossRef] [PubMed]
- Ou, A.; Yung, W.K.A.; Majd, N. Molecular mechanisms of drug resistance in glioblastoma. Int. J. Mol. Sci. 2020, 22, 6385. [Google Scholar] [CrossRef] [PubMed]
- Fidoamore, A.; Cristiano, L.; Antonosante, A.; D’Angelo, M.; Di Giacomo, E.; Astarita, C.; Giordano, A.; Ippoliti, R.; Benedetti, E.; Cimini, A. Glioblastoma Stem Cells Microenvironment: The Paracrine Roles of the Niche in Drug and Radioresistance. Stem Cells Int. 2016, 2016, 6809105. [Google Scholar] [CrossRef] [PubMed]
- Kenig, S.; Faoro, V.; Bourkoula, E.; Podergajs, N.; Ius, T.; Vindigni, M.; Skrap, M.; Lah, T.; Cesselli, D.; Storici, P.; et al. Topoisomerase IIβ mediates the resistance of glioblastoma stem cells to replication stress-inducing drugs. Cancer Cell Int. 2016, 16, 58. [Google Scholar] [CrossRef]
- Manini, I.; Ruaro, M.E.; Sgarra, R.; Bartolini, A.; Caponnetto, F.; Ius, T.; Skrap, M.; Di Loreto, C.; Beltrami, A.P.; Manfioletti, G.; et al. Semaphorin-7A on Exosomes: A Promigratory Signal in the Glioma Microenvironment. Cancers 2019, 11, 758. [Google Scholar] [CrossRef]
- Tang, X.; Zuo, C.; Fang, P.; Liu, G.; Qiu, Y.; Huang, Y.; Tang, R. Targeting Glioblastoma Stem Cells: A Review on Biomarkers, Signal Pathways and Targeted Therapy. Front. Oncol. 2021, 11, 701291. [Google Scholar] [CrossRef]
- Andersen, R.S.; Anand, A.; Harwood, D.S.L.; Kristensen, B.W. Tumor-associated microglia and macrophages in the glioblastoma microenvironment and their implications for therapy. Cancers 2021, 13, 4255. [Google Scholar] [CrossRef] [PubMed]
- Jarosz-Biej, M.; Smolarczyk, R.; Cichoń, T.; Kułach, N. Tumor microenvironment as a “game changer” in cancer radiotherapy. Int. J. Mol. Sci. 2019, 20, 3212. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Liao, K.; Zhuang, Z.; Chen, B.; Zhou, Z.; Zhou, S.; Lin, G.; Zhang, F.; Lin, Y.; Miao, Y.; et al. AHIF promotes glioblastoma progression and radioresistance via exosomes. Int. J. Oncol. 2019, 54, 261–270. [Google Scholar] [CrossRef]
- Hide, T.; Komohara, Y.; Miyasato, Y.; Nakamura, H.; Makino, K.; Takeya, M.; Kuratsu, J.-I.; Mukasa, A.; Yano, S. Oligodendrocyte Progenitor Cells and Macrophages/Microglia Produce Glioma Stem Cell Niches at the Tumor Border. eBioMedicine 2018, 30, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Zeng, A.; Zhang, Z.; Shi, Z.; Yan, W.; You, Y. Exosomal transfer of miR-1238 contributes to temozolomide-resistance in glioblastoma. eBioMedicine 2019, 42, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Caponnetto, F.; Dalla, E.; Mangoni, D.; Piazza, S.; Radovic, S.; Ius, T.; Skrap, M.; Di Loreto, C.; Beltrami, A.P.; Manini, I.; et al. The miRNA Content of Exosomes Released from the Glioma Microenvironment Can Affect Malignant Progression. Biomedicines 2020, 8, 564. [Google Scholar] [CrossRef]
- Byrne, N.M.; Tambe, P.; Coulter, J.A. Radiation response in the tumour microenvironment: Predictive biomarkers and future perspectives. J. Pers. Med. 2021, 11, 53. [Google Scholar] [CrossRef]
- Li, I.; Nabet, B.Y. Exosomes in the tumor microenvironment as mediators of cancer therapy resistance. Mol. Cancer 2019, 18, 32. [Google Scholar] [CrossRef]
- Okkenhaug, K.; Graupera, M.; Vanhaesebroeck, B. Targeting PI3K in Cancer: Impact on Tumor Cells, their Protective Stroma, Angiogenesis, and Immunotherapy. Cancer Discov. 2016, 6, 1090–1105. [Google Scholar] [CrossRef]
- Liu, X.; Huang, Y.; Qi, Y.; Wu, S.; Hu, F.; Wang, J.; Shu, K.; Zhang, H.; Bartsch, J.W.; Nimsky, C.; et al. The GBM Tumor Microenvironment as a Modulator of Therapy Response: ADAM8 Causes Tumor Infiltration of Tams through HB-EGF/EGFR-Mediated CCL2 Expression and Overcomes TMZ Chemosensitization in Glioblastoma. Cancers 2022, 14, 4910. [Google Scholar] [CrossRef]
- Jamal, M.; Rath, B.H.; Williams, E.S.; Camphausen, K.; Tofilon, P.J. Microenvironmental regulation of glioblastoma radioresponse. Clin. Cancer Res. 2010, 16, 6049–6059. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.H.; Wu, C.P.; Lee, H.T.; Liang, J.A.; Yu, C.Y.; Lin, Y.J. NADPH oxidase subunit 4 mediates cycling hypoxia-promoted radiation resistance in glioblastoma multiforme. Free. Radic. Biol. Med. 2012, 53, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yin, J.; Lu, C.; Wei, Y.; Zeng, A.; You, Y. Exosomal transfer of long non-coding RNA SBF2-AS1 enhances chemoresistance to temozolomide in glioblastoma. J. Exp. Clin. Cancer Res. 2019, 38, 166. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, J.; Zhang, Z.; Gao, Z.; Qi, Y.; Qiu, W.; Pan, Z.; Guo, Q.; Li, B.; Zhao, S.; et al. Hypoxic glioma-derived exosomes promote M2-like macrophage polarization by enhancing autophagy induction. Cell Death Dis. 2021, 12, 373. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Pessina, S.; Cantini, G.; Kapetis, D.; Cazzato, E.; Di Ianni, N.; Finocchiaro, G.; Pellegatta, S. The multidrug-resistance transporter Abcc3 protects NK cells from chemotherapy in a murine model of malignant glioma. Oncoimmunology 2016, 5, e1108513. [Google Scholar] [CrossRef]
- Azambuja, J.H.; da Silveira, E.F.; de Carvalho, T.R.; Oliveira, P.S.; Pacheco, S.; Couto, C.T.D.; Beira, F.T.; Stefanello, F.M.; Spanevello, R.M.; Braganhol, E. Glioma sensitive or chemoresistant to temozolomide differentially modulate macrophage protumor activities. Biochim. Biophys. Acta-Gen. Subj. 2017, 1861, 2652–2662. [Google Scholar] [CrossRef]
- Wu, P.; Cai, J.; Chen, Q.; Han, B.; Meng, X.; Li, Y.; Li, Z.; Wang, R.; Lin, L.; Duan, C.; et al. Lnc-TALC promotes O6-methylguanine-DNA methyltransferase expression via regulating the c-Met pathway by competitively binding with miR-20b-3p. Nat. Commun. 2019, 10, 2045. [Google Scholar] [CrossRef]
- Witusik-Perkowska, M.; Zakrzewska, M.; Jaskolski, D.J.; Liberski, P.P.; Szemraj, J. Artificial microenvironment of in vitro glioblastoma cell cultures changes profile of miRNAs related to tumor drug resistance. OncoTargets Ther. 2019, 12, 3905–3918. [Google Scholar] [CrossRef]
- Chuang, H.-Y.; Su, Y.-K.; Liu, H.-W.; Chen, C.-H.; Chiu, S.-C.; Cho, D.-Y.; Lin, S.-Z.; Chen, Y.-S.; Lin, C.-M. Preclinical evidence of STAT3 inhibitor pacritinib overcoming temozolomide resistance via downregulating miR-21-enriched exosomes from M2 glioblastoma-associated macrophages. J. Clin. Med. 2019, 8, 959. [Google Scholar] [CrossRef]
- Pustchi, S.E.; Avci, N.G.; Akay, Y.M.; Akay, M. Astrocytes decreased the sensitivity of glioblastoma cells to temozolomide and bay 11-7082. Int. J. Mol. Sci. 2020, 21, 7154. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kaneda, M.M.; Ma, J.; Li, M.; Shepard, R.M.; Patel, K.; Koga, T.; Sarver, A.; Furnari, F.; Xu, B.; et al. PI3Kγ inhibition suppresses microglia/TAM accumulation in glioblastoma microenvironment to promote exceptional temozolomide response. Proc. Natl. Acad. Sci. USA 2021, 118, e2009290118. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Xiang, W.; Zhang, Q.; Wang, H.; Zhou, Y.; Tian, H.; Abdelmaksou, A.; Xue, J.; Sun, M.; Yi, D.; et al. CD90low glioma-associated mesenchymal stromal/stem cells promote temozolomide resistance by activating FOXS1-mediated epithelial-mesenchymal transition in glioma cells. Stem Cell Res. Ther. 2021, 12, 394. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Meng, X.; Wu, P.; Zha, C.; Han, B.; Li, L.; Sun, N.; Qi, T.; Qin, J.; Zhang, Y.; et al. Glioblastoma Cell-Derived lncRNA-Containing Exosomes Induce Microglia to Produce Complement C5, Promoting Chemotherapy Resistance. Cancer Immunol. Res. 2021, 9, 1383–1399. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Shi, Q.; Fan, X.; Yu, R.; Wu, Z.; Wang, B.; Tian, W.; Yu, T.; Pan, M.; You, Y.; et al. Diverse Macrophages Constituted the Glioma Microenvironment and Influenced by PTEN Status. Front. Immunol. 2022, 13, 841404. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wang, B.; Zheng, R.; Zhang, J.; Huang, C.; Zheng, R.; Huang, Z.; Qiu, W.; Liu, M.; Yang, K.; et al. Linc-RA1 inhibits autophagy and promotes radioresistance by preventing H2Bub1/USP44 combination in glioma cells. Cell Death Dis. 2020, 11, 758. [Google Scholar] [CrossRef]
- Jang, B.S.; Kim, I.A. Relationship between Macrophage and Radiosensitivity in Human Primary and Recurrent Glioblastoma: In Silico Analysis with Publicly Available Datasets. Biomedicines 2022, 10, 292. [Google Scholar] [CrossRef]
- Chandran, M.; Candolfi, M.; Shah, D.; Mineharu, Y.; Yadav, V.N.; Koschmann, C.; Asad, A.S.; Lowenstein, P.R.; Castro, M.G. Single vs. combination immunotherapeutic strategies for glioma. Expert Opin. Biol. Ther. 2017, 17, 543–554. [Google Scholar] [CrossRef]
- Roesch, S.; Rapp, C.; Dettling, S.; Herold-Mende, C. When Immune Cells Turn Bad-Tumor-Associated Microglia/Macrophages in Glioma. Int. J. Mol. Sci. 2018, 19, 436. [Google Scholar] [CrossRef]
- Sahin, A.; Sanchez, C.; Bullain, S.; Waterman, P.; Weissleder, R.; Carter, B.S. Development of third generation anti-EGFRvIII chimeric T cells and EGFRvIII-expressing artificial antigen presenting cells for adoptive cell therapy for glioma. PLoS ONE 2018, 13, e0199414. [Google Scholar] [CrossRef]
- Goff, S.L.; Morgan, R.A.; Yang, J.C.; Sherry, R.M.; Robbins, P.F.; Restifo, N.P.; Feldman, S.A.; Lu, C.; Lu, L.; Zheng, Z.; et al. Pilot trial of adoptive transfer of chimeric antigen receptor transduced T cells targeting EGFRvIII in patients with glioblastoma. J. Immunother. 2019, 42, 126. [Google Scholar] [CrossRef] [PubMed]
- Flores-Toro, J.A.; Luo, D.; Gopinath, A.; Sarkisian, M.R.; Campbell, J.J.; Charo, I.F.; Singh, R.; Schall, T.J.; Datta, M.; Jain, R.K.; et al. CCR2 inhibition reduces tumor myeloid cells and unmasks a checkpoint inhibitor effect to slow progression of resistant murine gliomas. Proc. Natl. Acad. Sci. USA 2020, 117, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhao, Q.; Kong, Y.; Wang, J.; Yan, J.; Xia, X.; Jia, Z.; Heimberger, A.B.; Li, S. Regulation of tumor immune suppression and cancer cell survival by CXCL1/2 elevation in glioblastoma multiforme. Sci. Adv. 2021, 7, abc2511. [Google Scholar] [CrossRef] [PubMed]
- Serpe, C.; Monaco, L.; Relucenti, M.; Iovino, L.; Familiari, P.; Scavizzi, F.; Raspa, M.; Familiari, G.; Civiero, L.; D’Agnano, I.; et al. Microglia-Derived Small Extracellular Vesicles Reduce Glioma Growth by Modifying Tumor Cell Metabolism and Enhancing Glutamate Clearance through miR-124. Cells 2021, 10, 2066. [Google Scholar] [CrossRef]
- Andersen, J.K.; Miletic, H.; Hossain, J.A. Tumor-Associated Macrophages in Gliomas—Basic Insights and Treatment Opportunities. Cancers 2022, 14, 1319. [Google Scholar] [CrossRef] [PubMed]
- Xun, Y.; Yang, H.; Kaminska, B.; You, H. Toll-like receptors and toll-like receptor-targeted immunotherapy against glioma. J. Hematol. Oncol. 2021, 14, 176. [Google Scholar] [CrossRef]
- Ansems, M.; Span, P.N. The tumor microenvironment and radiotherapy response; a central role for cancer-associated fibroblasts. Clin. Transl. Radiat. Oncol. 2020, 22, 90–97. [Google Scholar] [CrossRef]
- Mannino, M.; Chalmers, A.J. Radioresistance of glioma stem cells: Intrinsic characteristic or property of the “microenvironment-stem cell unit”? Mol. Oncol. 2011, 5, 374–386. [Google Scholar] [CrossRef]
- Han, X.; Xue, X.; Zhou, H.; Zhang, G. A molecular view of the radioresistance of gliomas. Oncotarget 2017, 8, 100931–100941. [Google Scholar] [CrossRef]
- Miyazaki, T.; Ishikawa, E.; Matsuda, M.; Sugii, N.; Kohzuki, H.; Akutsu, H.; Sakamoto, N.; Takano, S.; Matsumura, A. Infiltration of CD163-positive macrophages in glioma tissues after treatment with anti-PD-L1 antibody and role of PI3Kγ inhibitor as a combination therapy with anti-PD-L1 antibody in in vivo model using temozolomide-resistant murine glioma-initiating cells. Brain Tumor Pathol. 2020, 37, 41–49. [Google Scholar] [CrossRef]
- Alhaddad, L.; Osipov, A.N.; Leonov, S. The Molecular and Cellular Strategies of Glioblastoma and Non-Small-Cell Lung Cancer Cells Conferring Radioresistance. Int. J. Mol. Sci. 2022, 23, 13577. [Google Scholar] [CrossRef]
- Locarno, C.V.; Simonelli, M.; Carenza, C.; Capucetti, A.; Stanzani, E.; Lorenzi, E.; Persico, P.; Della Bella, S.; Passoni, L.; Mavilio, D.; et al. Role of myeloid cells in the immunosuppressive microenvironment in gliomas. Immunobiology 2020, 225, 151853. [Google Scholar] [CrossRef] [PubMed]
- Perng, P.; Lim, M. Immunosuppressive mechanisms of malignant gliomas: Parallels at non-CNS sites. Front. Oncol. 2015, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Paulus, W.; Baur, I.; Huettner, C.; Schmausser, B.; Roggendorf, W.; Schlingensiepen, K.H.; Brysch, W.; SchmauBer, B. Effects of transforming growth factor-beta 1 on collagen synthesis, integrin expression, adhesion and invasion of glioma cells. J. Neuropathol. Exp. Neurol. 1995, 54, 236–244. [Google Scholar] [CrossRef]
- Quail, D.F.; Joyce, J.A. The microenvironmental landscape of brain tumors. Cancer Cell 2017, 31, 326–341. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhong, K.; Wang, Z.; Zhang, Z.; Tang, X.; Tong, A.; Zhou, L. Tumor-associated microglia and macrophages in glioblastoma:From basic insights to therapeutic opportunities. Front. Immunol. 2022, 13, 964898. [Google Scholar] [CrossRef]
- Bianconi, A.; Aruta, G.; Rizzo, F.; Salvati, L.F.; Zeppa, P.; Garbossa, D.; Cofano, F. Systematic Review on Tumor Microenvironment in Glial Neoplasm: From Understanding Pathogenesis to Future Therapeutic Perspectives. Int. J. Mol. Sci. 2022, 23, 4166. [Google Scholar] [CrossRef]
- Butowski, N.; Colman, H.; De Groot, J.F.; Omuro, A.M.; Nayak, L.; Wen, P.Y.; Cloughesy, T.F.; Marimuthu, A.; Haidar, S.; Perry, A.; et al. Orally administered colony stimulating factor 1 receptor inhibitor Plx3397 in recurrent glioblastoma: An ivy foundation early phase clinical trials consortium phase ii study. Neuro-Oncol. 2016, 18, 557–564. [Google Scholar] [CrossRef]
- Chen, L.; Li, J.; Wang, F.; Dai, C.; Wu, F.; Liu, X.; Li, T.; Glauben, R.; Zhang, Y.; Nie, G.; et al. Tie2 expression on macrophages is required for blood vessel reconstruction and tumor relapse after chemotherapy. Cancer Res. 2016, 76, 6828–6838. [Google Scholar] [CrossRef]
- Hyman, D.M.; Rizvi, N.A.; Natale, R.B.; Armstrong, D.K.; Birrer, M.J.; Recht, L.; Dotan, E.; Makker, V.; Kaley, T.J.; Kuruvilla, D.; et al. Phase I study of Medi3617, a selective angiopoietin-2 inhibitor alone and combined with Carboplatin/Paclitaxel, paclitaxel, or bevacizumab for advanced solid tumors. Clin. Cancer Res. 2018, 24, 2749–2757. [Google Scholar] [CrossRef]
- Martin-Liberal, J.; Hollebecque, A.; Aftimos, P.; Jungels, C.; Martin-Romano, P.; Rodon, J.; Kremer, J.D.; Zhang, W.; Bendell, J. First-in-Human, dose-escalation, phase 1 study of anti- Angiopoietin-2 Ly3127804 as monotherapy and in combination with ramucirumab in patients with advanced solid tumours. Br. J. Cancer 2020, 123, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.P.; Nagpal, S.; Iv, M.; Soltys, S.G.; Bertrand, S.; Pelpola, J.S.; Ball, R.; Yang, J.; Sundaram, V.; Chernikova, S.B.; et al. Macrophage exclusion after radiation therapy (Mert): A first in human phase I/Ii trial using a Cxcr4 inhibitor in glioblastoma. Clin. Cancer Res. 2019, 25, 6948–6957. [Google Scholar] [CrossRef]
- van Hooren, L.; Vaccaro, A.; Ramachandran, M.; Vazaios, K.; Libard, S.; van de Walle, T.; Georganaki, M.; Huang, H.; Pietilä, I.; Lau, J.; et al. Agonistic Cd40 therapy induces tertiary lymphoid structures but impairs responses to checkpoint blockade in glioma. Nat. Commun. 2021, 12, 4127. [Google Scholar] [CrossRef]
- Vonderheide, R.H. Cd40 agonist antibodies in cancer immunotherapy. Annu. Rev. Med. 2020, 71, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Zhu, S.; Qiao, Y.; Liu, Y.-J.; Chen, W.; Zhao, G.; Chen, J. Recent advances in the role of toll-like receptors and tlr agonists in immunotherapy for human glioma. Protein Cell 2014, 5, 899–911. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhou, J.; Luo, P.; Gao, H.; Ma, Y.; Chen, Y.-S.; Li, L.; Zou, D.; Zhang, Y.; Jing, Z. Prosaposin promotes the proliferation and tumorigenesis of glioma through toll-like receptor 4 (Tlr4)-mediated nf-kappab signaling pathway. eBioMedicine 2018, 37, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Morad, G.; Helmink, B.A.; Sharma, P.; Wargo, J.A. Hallmarks of response, resistance, and toxicity to immune checkpoint blockade. Cell 2021, 184, 5309–5337. [Google Scholar] [CrossRef] [PubMed]
- Hartley, G.P.; Chow, L.; Ammons, D.T.; Wheat, W.H.; Dow, S.W. Programmed cell death ligand 1 (Pd-L1) signaling regulates macrophage proliferation and activation. Cancer Immunol. Res. 2018, 6, 1260–1273. [Google Scholar] [CrossRef] [PubMed]
- Crivii, C.B.; Boșca, A.B.; Melincovici, C.S.; Constantin, A.M.; Mărginean, M.; Dronca, E.; Suflețel, R.; Gonciar, D.; Bungărdean, M.; Șovrea, A. Glioblastoma Microenvironment and Cellular Interactions. Cancers 2022, 14, 1092. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, J.; Zheng, H.; Li, C.; Xiong, J.; Wang, W.; Bao, H.; Jin, H.; Liang, P. Modulating lncRNA SNHG15/CDK6/miR-627 circuit by palbociclib, overcomes temozolomide resistance and reduces M2-polarization of glioma associated microglia in glioblastoma multiforme. J. Exp. Clin. Cancer Res. 2019, 38, 380. [Google Scholar] [CrossRef]
- Hegi, M.E.; Rajakannu, P.; Weller, M. Epidermal growth factor receptor: A re-emerging target in glioblastoma. Curr. Opin. Neurol. 2012, 25, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, M.; Zhao, J.; Ren, T.; Yan, X.; Zhang, L.; Wang, X. Research Progress About Glioma Stem Cells in the Immune Microenvironment of Glioma. Front. Pharmacol. 2021, 12, 750857. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author, Year | Methods | Cell Lines Studied | Mechanism of Resistance |
|---|---|---|---|
| Pessina et al., 2015 [26] | Cellular culture and immunohistochemistry | Murine NK1.1 + CD3-; human GBM GL261 | Multidrug-resistance transporter Abcc3 provides TMZ resistance to NK cells |
| Azambuja et al., 2017 [27] | Cellular culture and immunohistochemistry | Human GBM GL261; human GBM cells–astrocytes LN229 | M2 macrophage polarization with high IL-10 release and antioxidant potential of the TME contribute to GBM TMZ resistance |
| Hide et al., 2018 [14] | Cellular culture and immunohistochemistry | Human GBM A172 and T98G | M2 macrophages produce HB-EGF and IL-1β, conferring TMZ resistance potential on GBM cells |
| Zhang et al., 2019 [23] | Cellular culture and immunohistochemistry | Human GBM U87, LN229, A172, T98, U251; human embryonic kidney 293 T cells | Exosomal transfer of lnc RNASBF2-AS1 from TME to GBM enhances chemoresistance to TMZ |
| Yin et al., 2019 [15] | Cellular culture and RT-qPCR | Human GBM U251 and GBM-1 | MiR-1238 exosomes levels are higher in TMZ-resistant GBM |
| Wu et al., 2019 [28] | Cellular culture and RT-qPCR | Human GBM LN229, U251, 551 W, and HG7 | TMZ-associated lncRNA in GBM recurrence (lnc-TALC) may play a role in regulating the c-Met signaling pathway, obtained by activation of the Stat3/p300 complex due to competitive binding with miR-20b-3p. This can promote DNA repair enzyme expression of O-methylguanine-DNA methyl-transferase (MGMT) and favor TMZ resistance due to histone H3 acetylation modulation. |
| Witusik-Perkowska et al., 2019 [29] | Cellular culture and RT-qPCR | Human GBM cell cultures derived from three patients with GBM | TME of in vitro GBM cell cultures changes the profile of specific miRNAs related to tumor drug resistance (miRNA-221, miRNA-31, miRNA-21, miRNA-222) |
| Chuang et al., 2019 [30] | Cellular culture and RT-qPCR | Human GBM U87MG and LN18 | miRNA-21-enriched exosomes from M2 GBM-associated macrophages provide TMZ resistance to GBM; STAT3 Inhibitor Pacritinib can overcome this mechanism of TMZ resistance |
| Pustchi et al., 2020 [31] | Cellular culture, immunohistochemistry | LN229 GBM cells–astrocytes | GFAP-vimentin and Notch1-survivin signaling in astrocytes of TME are implicated in TMZ resistance of GBM |
| Li et al., 2021 [32] | Cellular culture genomics | Human microglial HMC3 and murine microglial BV-2 | GBM-associated microglia secreted IL11 to activate STAT3-MYC signaling, inducing enhanced TMZ resistance |
| Xue et al., 2021 [33] | Cellular culture | Human GBM U87 and GBM-1 | CD90low GBM-associated mesenchymal stem and stromal cells favor TMZ resistance by the activation of FOXS1-mediated epithelial-mesenchymal transition in GBM cells |
| Li et al., 2021 [34] | Cellular culture, genomics, and immunohistochemistry | Human microglial HMC3, and murine microglial cell line BV-2 | GBM cell-derived lncRNA-containing exosomes induce microglia to produce Complement C5 and develop TMZ resistance |
| Zhou et al., 2022 [35] | Cellular culture and immunohistochemistry | Human GBM U251, LN229, and U87; human monocyte cell line THP-1 cells | PTEN loss mutation in the macrophages of the TME may be associated with the development of chemoresistance |
| Zhang et al., 2022 [5] | Cellular culture, genomics, and immunohistochemistry | Human GBM LN229 and U251; THP-1 monocyte-derived macrophages | Hypoxic M2 macrophages can activate the PI3K/Akt/Nrf2 pathway by the secretion of VEGF in GBM cells to favor cancer stemness, aggressiveness, and TMZ resistance. |
| Liu et al., 2022 [20] | Cellular culture, genomics, and immunohistochemistry | Human GBM U87MG and U251MG; THP-1 monocyte | ADAM8 causes tumor infiltration of tumor-associated macrophages through HB-EGF/EGFR-mediated CCL2 expression and promotes TMZ resistance in GBM |
| Author, Year | Methods | Cell Lines Studied | Mechanism of Resistance |
|---|---|---|---|
| Jamal et al., 2010 [21] | Cellular culture and immunohistochemistry | Human GBM NSC11 and GBMJ1 | Orthotopic xenografts GBM cells can show increased capability of repairing DNA double-strand breaks and tend to be less susceptible to induction when compared with cells cultured in vitro, thus promoting the TME as a possible source of GBM radioresistance |
| Hsieh et al., 2012 [22] | Cellular culture and immunohistochemistry | GBM cell lines GBM8401 and U251 | The subunit 4 in NADPH oxidase of the TME can favor GBM cycling hypoxia-promoted radiation resistance |
| Hide et al., 2018 [14] | Cellular culture and immunohistochemistry | Human GBM A172 and T98G, human GBM cells | Oligodendrocyte progenitor cells secrete FGF1 and EGF, and macrophages produce HB-EGF and IL-1β, conferring stemness radioresistant potential on GBM cells |
| Dai et al., 2018 [13] | Cellular culture and RT-qPCR | Human GBM U87-MG, U251-MG, A172 and T98G. | LncRNA AHIF promotes GBM progression and radioresistance via exosomes |
| Zheng et al., 2020 [36] | Cellular culture and RT-qPCR | Human GBM M059J, U251, M059K, and U87 | LncRNA linc-RA1 inhibits autophagy and promotes Radioresistance by preventing H2Bub1/USP44 combination in GBM cells |
| Jang et al., 2022 [37] | Cellular culture and immunohistochemistry | M1/M2 macrophages | M1/M2 macrophage ratios and radiosensitivity are inversely associated: radioresistant TME contain more M2 than M1 macrophages |
| Zhou et al., 2022 [35] | Cellular culture and immunohistochemistry | GBM cell lines (U251, LN229, and U87) and the human monocyte cell line THP-1 cells | PTEN loss mutation in the macrophages of the TME may be associated with the development of radioresistance |
| Author, Year | Type of Article | Cell and Pathway Involved | Target Molecules | Drug Tested | Type of Test (Human/Mice/In Vitro) | Results |
|---|---|---|---|---|---|---|
| Chandran et al., 2017 [38] | Review | Tumor associated lymphocytes | CTLA4-PD1 | CTLA4-PD1 inhibitors (e.g., Ipilimumab) | In vitro | OS improvement in association with CTX and RT |
| Roesch et al., 2018 [39] | Review | GAMs | VEGF-VEGFR; IL-10, STAT3 | Sunitinib, Bevacizumab; STAT3 inhibitor | In vitro | Increased OS and tumor growth inhibition |
| Sahin et al., 2018 [40] | Research | Chimeric T Cells | EGFRvIII | Anti-EGFRvIII Chimeric T Cells | In vitro and mice | Increased OS in mice |
| Goff et al., 2019 [41] | Research | Chimeric T Cells | EGFRvIII | Anti-EGFRvIII Chimeric T Cells | Human (phase I pilot trial) | No OS increment |
| Li et al., 2019 [18] | Research | GAMs | CDK6 | Palbociclib | In vitro | Increased TMZ sensitivity |
| Flores-Toro et al., 2020 [42] | Research | GSCs | CCR2 and PD-1 | CCR2 antagonist CCX872 and anti-PD-1 | Mice | Increased OS |
| Hu et al., 2021 [43] | Research | Cytokines | CXCL1/2 | CXCL1/2 inhibitor + TMZ | Mice | Increased OS |
| Li et al., 2021 [34] | Review | GSCs | SOX2 | Zika virus | In vitro | Apoptosis of GSCs |
| Serpe et al., 2021 [44] | Research | Extracellular vesicles | miRNA-124 | miRNA-124 upregulation | In vitro and mice | Tumor mass reduction in vitro and increased OS in mice |
| Andersen et al., 2021 [45] | Review | GAMs | TNF-α and NF-κB | inhibition of NF-κB signaling | In vitro and mice | Increased infiltration of cytotoxic T cells and decreased tumor growth. |
| Xun et al., 2021 [46] | Review | TLRs | TL3, TLR7, and TLR8 | Hiltonol (TLR3 agonist) and Imiquimod (TLR7 and TLR8 agonist) | Mice | Increase in PFS and OS |
| Category of Immunotherapeutic Agent | Drug Tested | Combined Therapies | Clinical Trials Registry Identifier | Trial Phase | Patients (n) | Reported Biological Response |
|---|---|---|---|---|---|---|
| CSF1R Inhibitors | Pexidartinib | / | NCT01349036 | II | 38 | Monocyte ↓ |
| ANG2 Inhibitors | MEDI3617 | Bevacizumab | NCT01248949 | I | 13 | ORR: 0% |
| Trebananib | Bevacizumab | NCT01609790 | II | 130 | NR | |
| CXCR4 Inhibitors | Plerixafor | TMZ + RT | NCT01977677 | I/II | 29 | CXCL12 ↑ |
| Plerixafor | Bevacizumab | NCT01339039 | I | 26 | Lymphocytes ↑, Monocytes ↑ CXCL12 ↑, ANG2 ↓, sMET ↓, IL-8 ↓ | |
| CD40 Agonists | APX005M | / | NCT03389802 | I | 45 | NR |
| 2141-V11 | D2C7-IT | NCT04547777 | I | 30 | NR | |
| TLR Agonists | Poly-ICLC (TLR3 agonist) | / | NCT01188096 | II | 47 | 50% LGG respond, 25% HGG respond |
| RT | NCT00052715 | II | 31 | NR | ||
| TMZ + RT | NCT00262730 | II | 97 | NR | ||
| GAA/TT-Peptide Vaccine | NCT00795457 | I | 13 | 91% respond | ||
| Peptide Vaccines | NCT00874861 | I | 10 | 55% respond | ||
| IMA950 Peptide Vaccine | NCT01920191 | I/II | 19 | NR | ||
| Dendritic Cell Vaccine | NCT00068510 | I | 28 | TNF-a ↑, IL-6 ↑, Lymphocytes ↑ | ||
| HSPPC-96 (TLR4 agonist) | / | NCT02122822 | I | 20 | NR | |
| / | NCT00293423 | II | 96 | NR | ||
| CpG-ODN (TLR9 agonist) | / | NCT00190424 | II | 34 | No benefit | |
| PD-1 Inhibitors | Nivolumab | / | NCT02017717 | III | 529 | ORR: 7.8% |
| / | NCT02550249 | II | 29 | CXCL10 ↑, CCL4 ↑, CCL3L1 ↑ | ||
| Ipilimumab | NCT03233152 | I | 27 | No benefit | ||
| Lirilumab | NCT02813135 | II | 397 | NR | ||
| Cemiplimab | Veledimex | NCT04006119 | 40 | NR | ||
| Pembrolizumab | Bevacizumab + RT | NCT02313272 | 32 | NR | ||
| Bevacizumab | NCT02337491 | 80 | ORR:20% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agosti, E.; Panciani, P.P.; Zeppieri, M.; De Maria, L.; Pasqualetti, F.; Tel, A.; Zanin, L.; Fontanella, M.M.; Ius, T. Tumor Microenvironment and Glioblastoma Cell Interplay as Promoters of Therapeutic Resistance. Biology 2023, 12, 736. https://doi.org/10.3390/biology12050736
Agosti E, Panciani PP, Zeppieri M, De Maria L, Pasqualetti F, Tel A, Zanin L, Fontanella MM, Ius T. Tumor Microenvironment and Glioblastoma Cell Interplay as Promoters of Therapeutic Resistance. Biology. 2023; 12(5):736. https://doi.org/10.3390/biology12050736
Chicago/Turabian StyleAgosti, Edoardo, Pier Paolo Panciani, Marco Zeppieri, Lucio De Maria, Francesco Pasqualetti, Alessandro Tel, Luca Zanin, Marco Maria Fontanella, and Tamara Ius. 2023. "Tumor Microenvironment and Glioblastoma Cell Interplay as Promoters of Therapeutic Resistance" Biology 12, no. 5: 736. https://doi.org/10.3390/biology12050736