

Evidence for Prepulse Inhibition of Visually Evoked Motor Response in Drosophila melanogaster
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Strains and Maintenance
2.2. NMDA Receptor Antagonist
2.3. DISCO Platform
2.4. Movement Response, PPI and Stimuli Desensitization
2.5. Statistical Analyses
3. Results
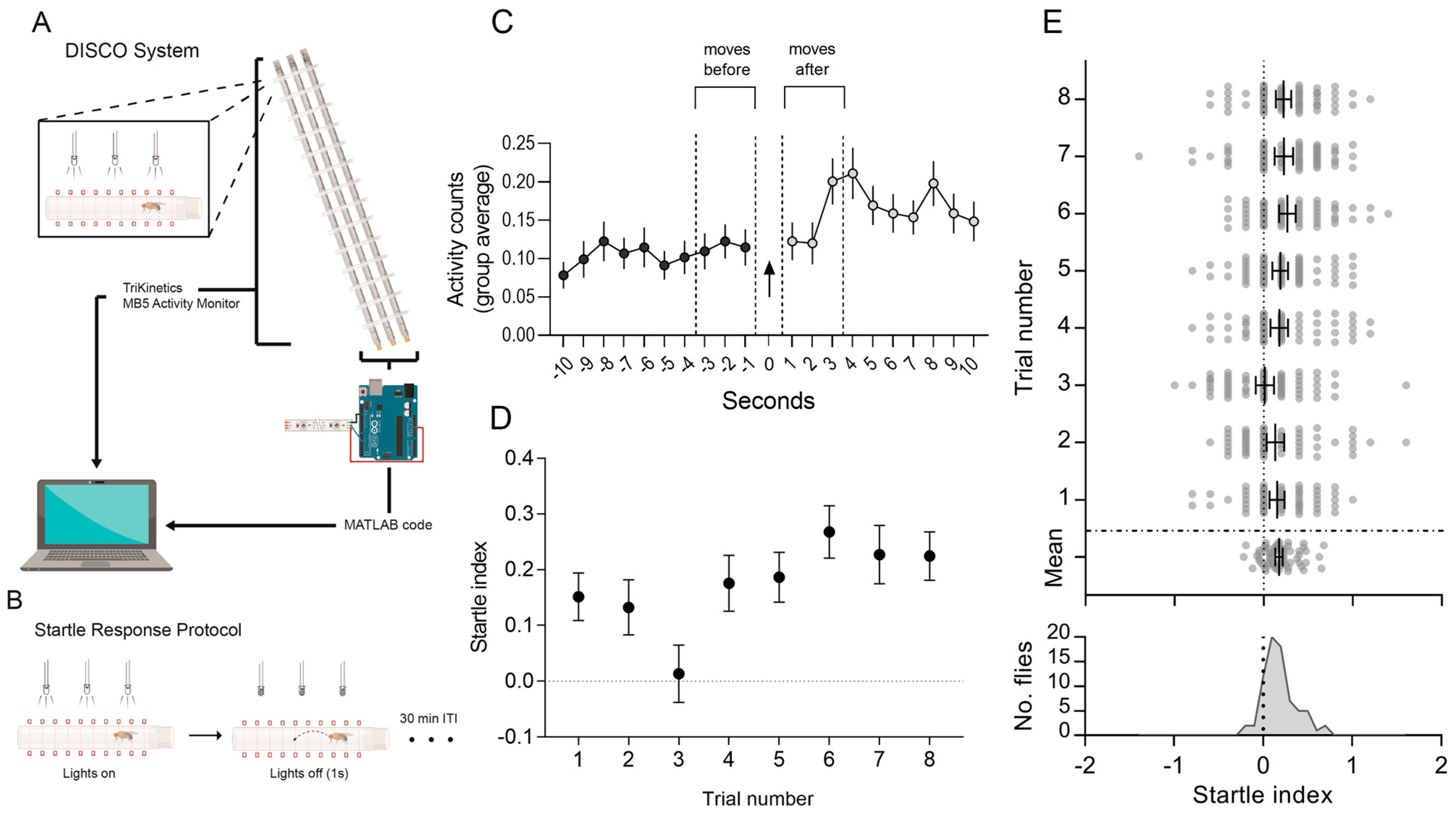
3.1. Automatic Detection of Escape Reflex
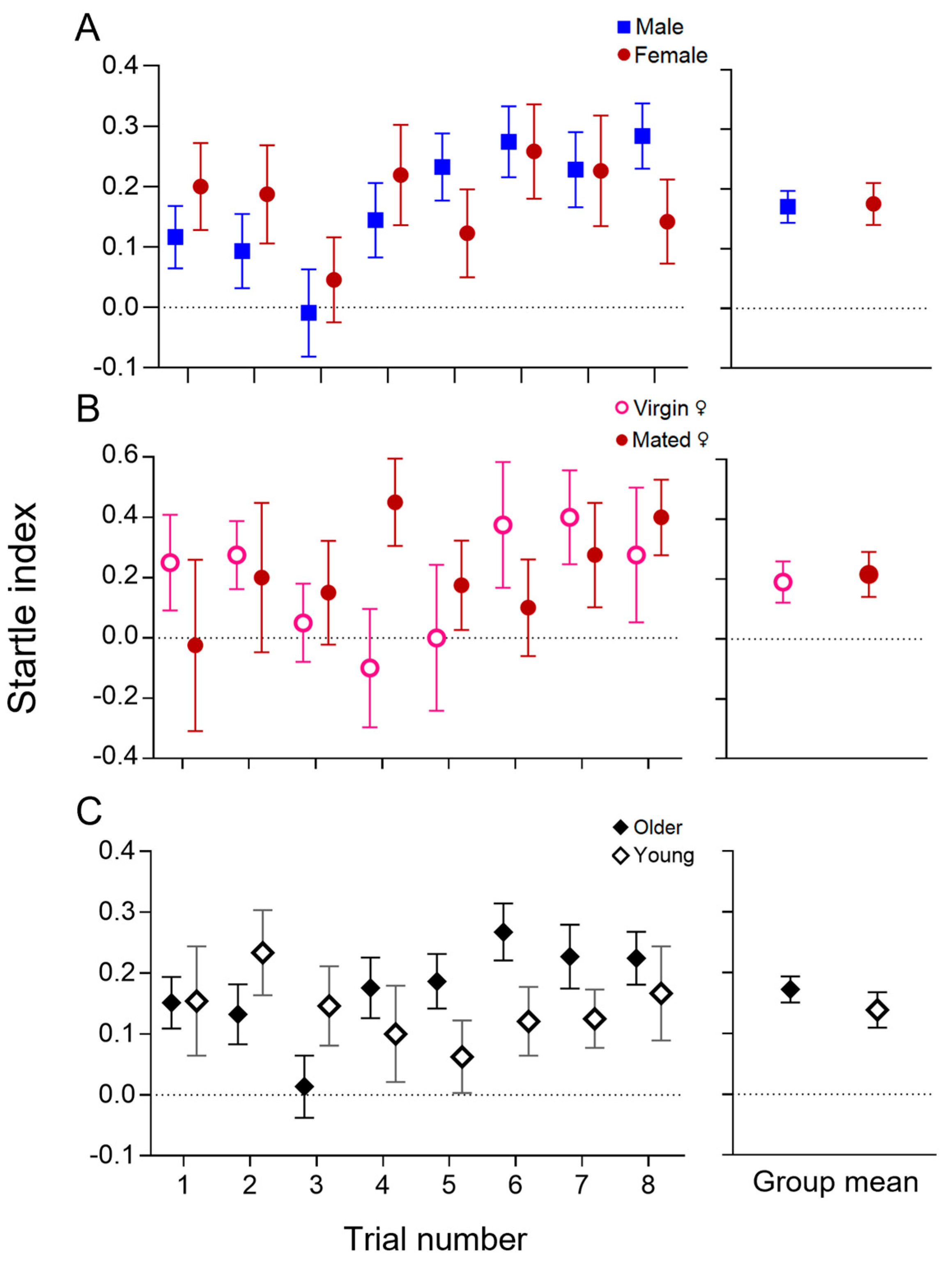
3.2. Characterisation of the Visually Evoked Response
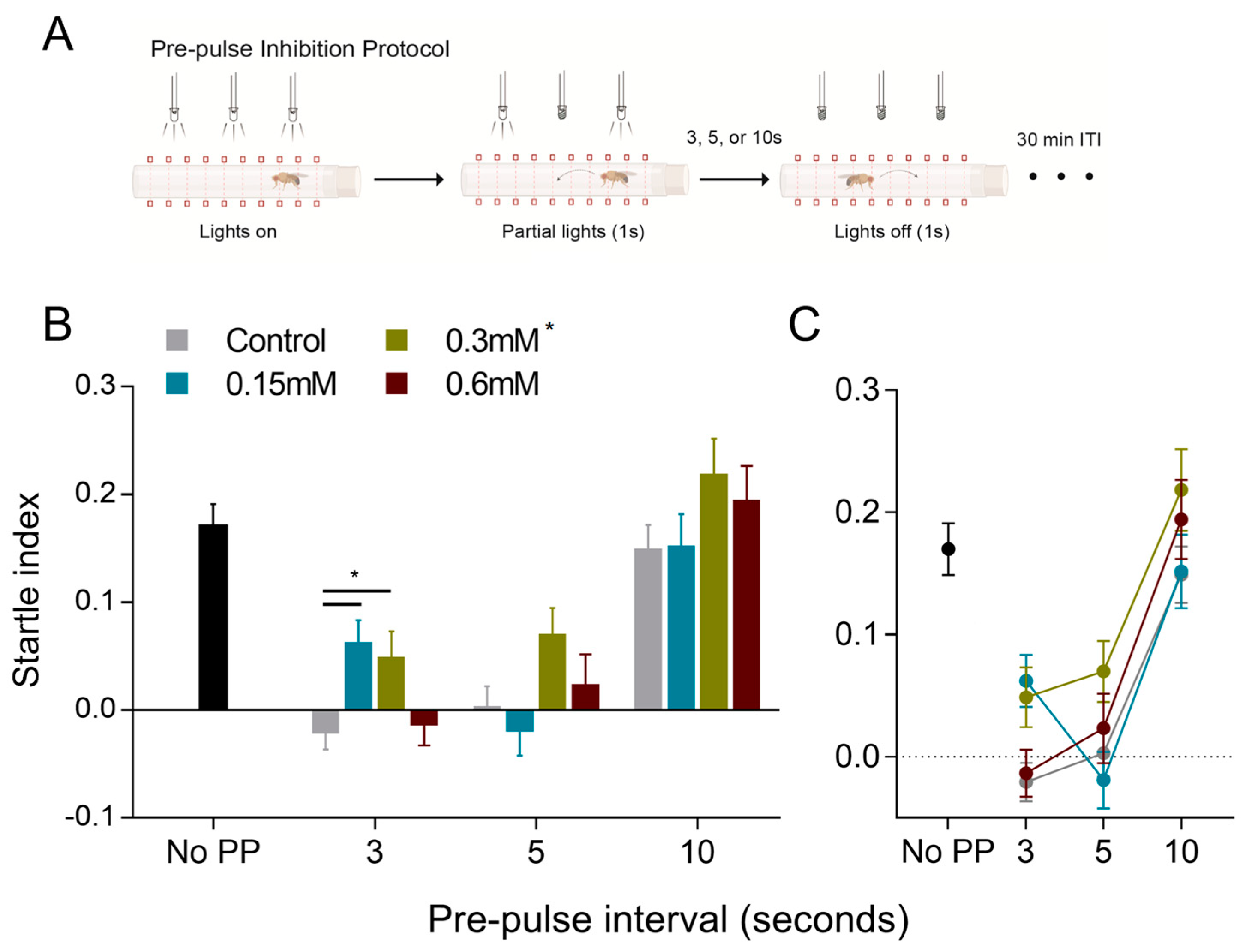
3.3. Prepulse Inhibition of the Movement Response
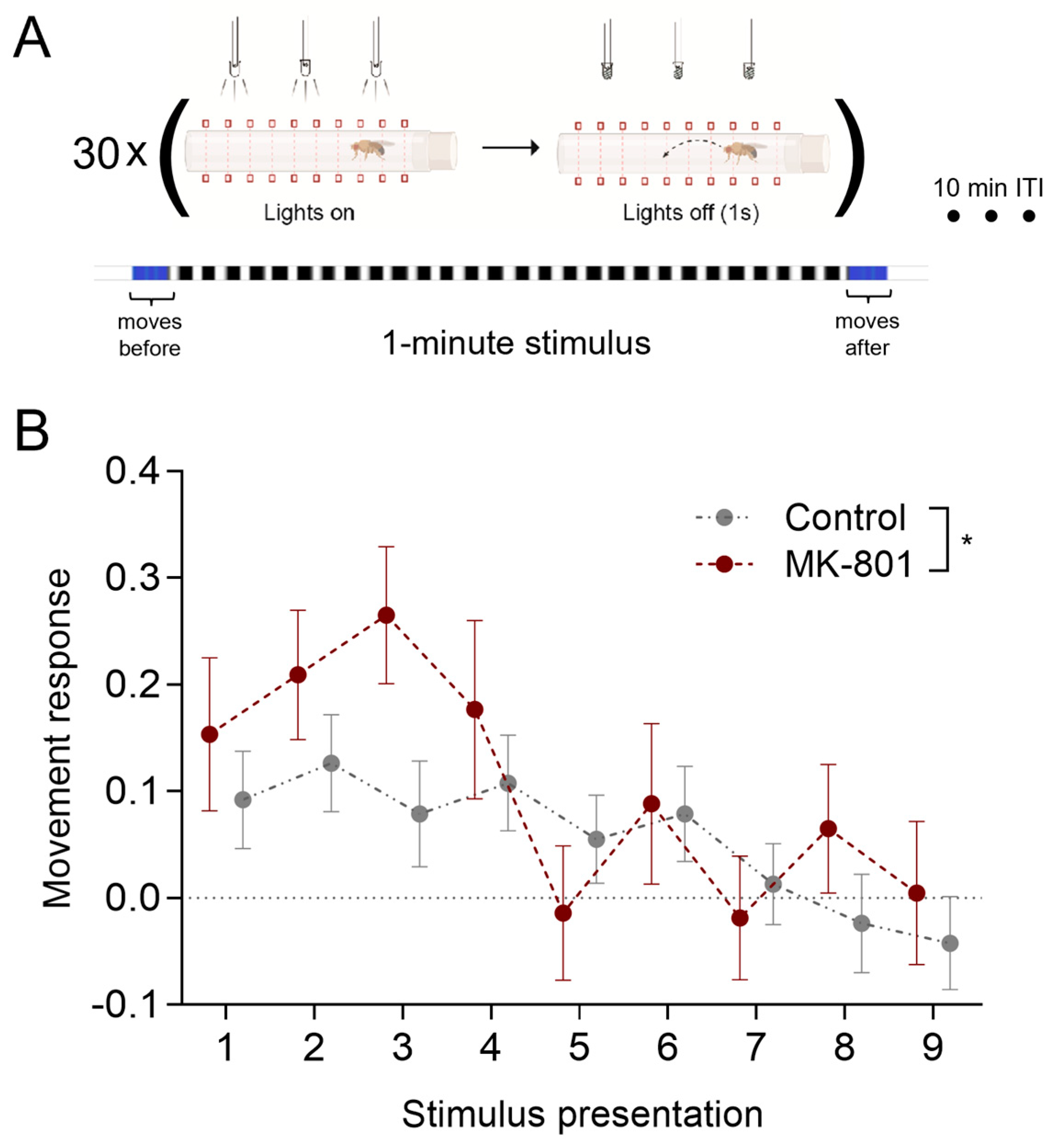
3.4. Inhibition of Visually Evoked Response by Multiple Stimuli Presentation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Augustin, H.; Zylberta, A.; Partridge, L. A Computational Model of the Escape Response Latency in the Giant Fiber System of Drosophila melanogaster. eNeuro 2019, 6, ENEURO.0423-18.2019. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.J.; Godenschwege, T.A.; Tanouye, M.A.; Phelan, P. Making an Escape: Development and Function of the Drosophila Giant Fibre System. Semin. Cell Dev. Biol. 2006, 17, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Trimarchi, J.R.; Schneiderman, A.M. Different Neural Pathways Coordinate Drosophila Flight Initiations Evoked by Visual and Olfactory Stimuli. J. Exp. Biol. 1995, 198, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Meinck, H.M. Startle and Its Disorders. Neurophysiol. Clin. 2006, 36, 357–364. [Google Scholar] [CrossRef]
- Vaidyanathan, U.; Patrick, C.J.; Cuthbert, B.N. Linking Dimensional Models of Internalizing Psychopathology to Neurobiological Systems: Affect-Modulated Startle as an Indicator of Fear and Distress Disorders and Affiliated Traits. Psychol. Bull. 2009, 135, 909–942. [Google Scholar] [CrossRef] [PubMed]
- Grillon, C. Models and Mechanisms of Anxiety: Evidence from Startle Studies. Psychopharmacology 2008, 199, 421–437. [Google Scholar] [CrossRef]
- Geyer, M.A. Developing Translational Animal Models for Symptoms of Schizophrenia or Bipolar Mania. Neurotox. Res. 2008, 14, 71–78. [Google Scholar] [CrossRef]
- Swerdlow, N.R.; Sutherland, A.N. Using Animal Models to Develop Therapeutics for Tourette Syndrome. Pharmacol. Ther. 2005, 108, 281–293. [Google Scholar] [CrossRef]
- Hayes, S.H.; Radziwon, K.E.; Stolzberg, D.J.; Salvi, R.J. Behavioral Models of Tinnitus and Hyperacusis in Animals. Front. Neurol. 2014, 5, 179. [Google Scholar] [CrossRef]
- Eggermont, J.J. Animal Models of Spontaneous Activity in the Healthy and Impaired Auditory System. Front. Neural Circuits 2015, 9, 19. [Google Scholar] [CrossRef]
- Török, B.; Sipos, E.; Pivac, N.; Zelena, D. Modelling Posttraumatic Stress Disorders in Animals. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 90, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, N.R.; Braff, D.L.; Geyer, M.A. Sensorimotor Gating of the Startle Reflex: What We Said 25 Years Ago, What Has Happened Since Then, and What Comes Next. J. Psychopharmacol. 2016, 30, 1072–1081. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, N.R.; Talledo, J.; Sutherland, A.N.; Nagy, D.; Shoemaker, J.M. Antipsychotic Effects on Prepulse Inhibition in Normal “low Gating” Humans and Rats. Neuropsychopharmacology 2006, 31, 2011–2021. [Google Scholar] [CrossRef] [PubMed]
- Valls-Solé, J.; Muñoz, J.E.; Valldeoriola, F. Abnormalities of Prepulse Inhibition Do not Depend on Blink Reflex Excitability: A Study in Parkinson’s Disease and Huntington’s Disease. Clin. Neurophysiol. 2004, 115, 1527–1536. [Google Scholar] [CrossRef]
- Ahmari, S.E.; Risbrough, V.B.; Geyer, M.A.; Simpson, H.B. Impaired Sensorimotor Gating in Unmedicated Adults with Obsessive-Compulsive Disorder. Neuropsychopharmacology 2012, 37, 1216–1223. [Google Scholar] [CrossRef]
- McAlonan, G.M. Brain Anatomy and Sensorimotor Gating in Asperger’s Syndrome. Brain 2002, 125, 1594–1606. [Google Scholar] [CrossRef]
- Sobin, C.; Kiley-Brabeck, K.; Karayiorgou, M. Lower Prepulse Inhibition in Children with the 22q11 Deletion Syndrome. Am. J. Psychiatry 2005, 162, 1090–1099. [Google Scholar] [CrossRef]
- Frankland, P.W.; Wang, Y.; Rosner, B.; Shimizu, T.; Balleine, B.W.; Dykens, E.M.; Ornitz, E.M.; Silva, A.J. Sensorimotor Gating Abnormalities in Young Males with Fragile X Syndrome and Fmr1-Knockout Mice. Mol. Psychiatry 2004, 9, 417–425. [Google Scholar] [CrossRef]
- Braff, D.L.; Geyer, M.A.; Swerdlow, N.R. Human Studies of Prepulse Inhibition of Startle: Normal Subjects, Patient Groups, and Pharmacological Studies. Psychopharmacology 2001, 156, 234–258. [Google Scholar] [CrossRef]
- Winslow, J.T.; Parr, L.A.; Davis, M. Acoustic Startle, Prepulse Inhibition, and Fear-Potentiated Startle Measured in Rhesus Monkeys. Biol. Psychiatry 2002, 51, 859–866. [Google Scholar] [CrossRef]
- Mongeluzi, D.L.; Hoppe, T.A.; Frost, W.N. Prepulse Inhibition of the Tritonia Escape Swim. J. Neurosci. 1998, 18, 8467–8472. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Shimizu, K.; Arahata, K.; Suzuki, M.; Shimizu, A.; Takei, K.; Yamauchi, J.; Hakeda-Suzuki, S.; Suzuki, T.; Morimoto, T. Prepulse Inhibition in Drosophila melanogaster Larvae. Biol. Open 2018, 7, bio034710. [Google Scholar] [CrossRef]
- Kelly, L.E. An Altered Electroretinogram Transient Associated with an Unusual Jump Response in a Mutant of Drosophila. Cell. Mol. Neurobiol. 1983, 3, 143–149. [Google Scholar] [CrossRef]
- Kepler, L.D.; McDiarmid, T.A.; Rankin, C.H. Habituation in High-Throughput Genetic Model Organisms as a Tool to Investigate the Mechanisms of Neurodevelopmental Disorders. Neurobiol. Learn. Mem. 2020, 171, 107208. [Google Scholar] [CrossRef]
- Engel, J.E.; Wu, C.-F.F. Neurogenetic Approaches to Habituation and Dishabituation in Drosophila. Neurobiol. Learn. Mem. 2009, 92, 166–175. [Google Scholar] [CrossRef]
- Engel, J.E.; Wu, C.F. Altered Habituation of an Identified Escape Circuit in Drosophila Memory Mutants. J. Neurosci. 1996, 16, 3486–3499. [Google Scholar] [CrossRef]
- Fischbach, K.F. Habituation and Sensitization of the Landing Response of Drosophila melanogaster. Naturwissenschaften 1981, 68, 332. [Google Scholar] [CrossRef]
- Fenckova, M.; Blok, L.E.R.; Asztalos, L.; Goodman, D.P.; Cizek, P.; Singgih, E.L.; Glennon, J.C.; IntHout, J.; Zweier, C.; Eichler, E.E.; et al. Habituation Learning Is a Widely Affected Mechanism in Drosophila Models of Intellectual Disability and Autism Spectrum Disorders. Biol. Psychiatry 2019, 86, 294–305. [Google Scholar] [CrossRef]
- von Reyn, C.R.; Nern, A.; Williamson, W.R.; Breads, P.; Wu, M.; Namiki, S.; Card, G.M. Feature Integration Drives Probabilistic Behavior in the Drosophila Escape Response. Neuron 2017, 94, 1190–1204.e6. [Google Scholar] [CrossRef]
- Moulin, T.C.; Dey, S.; Dashi, G.; Li, L.; Sridhar, V.; Safa, T.; Berkins, S.; Williams, M.J.; Schiöth, H.B. A Simple High-Throughput Method for Automated Detection of Drosophila melanogaster Light-Dependent Behaviours. BMC Biol. 2022, 20, 1–13. [Google Scholar] [CrossRef]
- Franco, L.M.; Okray, Z.; Linneweber, G.A.; Hassan, B.A.; Yaksi, E. Reduced Lateral Inhibition Impairs Olfactory Computations and Behaviors in a Drosophila Model of Fragile X Syndrome. Curr. Biol. 2017, 27, 1111–1123. [Google Scholar] [CrossRef]
- Geyer, M.A.; Krebs-Thomson, K.; Braff, D.L.; Swerdlow, N.R. Pharmacological Studies of Prepulse Inhibition Models of Sensorimotor Gating Deficits in Schizophrenia: A Decade in Review. Psychopharmacology 2001, 156, 117–154. [Google Scholar] [CrossRef]
- Moulin, T.C.; Stojanovic, T.; Rajesh, R.P.; Pareek, T.; Donzelli, L.; Williams, M.J.; Schiöth, H.B. Effects of Transient Administration of the NMDA Receptor Antagonist MK-801 in Drosophila melanogaster Activity, Sleep, and Negative Geotaxis. Biomedicines 2023, 11, 192. [Google Scholar] [CrossRef]
- Klamer, D.; Pålsson, E.; Revesz, A.; Engel, J.A.; Svensson, L. Habituation of Acoustic Startle Is Disrupted by Psychotomimetic Drugs: Differential Dependence on Dopaminergic and Nitric Oxide Modulatory Mechanisms. Psychopharmacology 2004, 176, 440–450. [Google Scholar] [CrossRef]
- Gibbs, A.G.; Chippindale, A.K.; Rose, M.R. Physiological Mechanisms of Evolved Desiccation Resistance in Drosophila melanogaster. J. Exp. Biol. 1997, 200, 1821–1832. [Google Scholar] [CrossRef]
- Aickin, M.; Gensler, H. Adjusting for Multiple Testing When Reporting Research Results: The Bonferroni vs. Holm Methods. Am. J. Public Health 1996, 86, 726–728. [Google Scholar] [CrossRef]
- Gibson, W.T.; Gonzalez, C.R.; Fernandez, C.; Ramasamy, L.; Tabachnik, T.; Du, R.R.; Felsen, P.D.; Maire, M.R.; Perona, P.; Anderson, D.J. Behavioral Responses to a Repetitive Visual Threat Stimulus Express a Persistent State of Defensive Arousal in Drosophila. Curr. Biol. 2015, 25, 1401–1415. [Google Scholar] [CrossRef]
- Cobb, T.; Sujkowski, A.; Morton, C.; Ramesh, D.; Wessells, R. Variation in Mobility and Exercise Adaptations between Drosophila Species. J. Comp. Physiol. A 2020, 206, 611–621. [Google Scholar] [CrossRef]
- Videlier, M.; Rundle, H.D.; Careau, V. Sex-Specific Among-Individual Covariation in Locomotor Activity and Resting Metabolic Rate in Drosophila melanogaster. Am. Nat. 2019, 194, E164–E176. [Google Scholar] [CrossRef]
- Moulin, T.C.; Ferro, F.; Hoyer, A.; Cheung, P.; Williams, M.J.; Schiöth, H.B. The Drosophila melanogaster Levodopa-Induced Depression Model Exhibits Negative Geotaxis Deficits and Differential Gene Expression in Males and Females. Front. Neurosci. 2021, 15, 653470. [Google Scholar] [CrossRef]
- Qiu, S.; Li, C.; Cao, G.; Xiao, C. Mating Experience Modifies Locomotor Performance and Promotes Episodic Motor Activity in Drosophila melanogaster. Zoology 2021, 144, 125854. [Google Scholar] [CrossRef] [PubMed]
- Valsamis, B.; Schmid, S. Habituation and Prepulse Inhibition of Acoustic Startle in Rodents. J. Vis. Exp. 2011, 55, e3446. [Google Scholar] [CrossRef]
- Davis, M.; Sheard, M.H. Effects of Lysergic Acid Diethylamide (LSD) on Temporal Recovery (Pre-Pulse Inhibition) of the Acoustic Startle Response in the Rat. Pharmacol. Biochem. Behav. 1975, 3, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Groves, P.; Boyle, R.; Welker, R.; Miller, S. On the Mechanism of Prepulse Inhibition☆. Physiol. Behav. 1974, 12, 367–375. [Google Scholar] [CrossRef]
- Mansbach, R.S. Effects of NMDA Receptor Ligands on Sensorimotor Gating in the Rat. Eur. J. Pharmacol. 1991, 202, 61–66. [Google Scholar] [CrossRef]
- Xia, S.; Chiang, A.S. NMDA Receptors in Drosophila. In Biology of the NMDA Receptor, 1st ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 213–233. [Google Scholar] [CrossRef]
- Brockie, P.J.; Madsen, D.M.; Zheng, Y.; Mellem, J.; Maricq, A.V. Differential Expression of Glutamate Receptor Subunits in the Nervous System of Caenorhabditis Elegans and Their Regulation by the Homeodomain Protein UNC-42. J. Neurosci. 2001, 21, 1510–1522. [Google Scholar] [CrossRef]
- Zannat, M.T.; Locatelli, F.; Rybak, J.; Menzel, R.; Leboulle, G. Identification and Localisation of the NR1 Sub-Unit Homologue of the NMDA Glutamate Receptor in the Honeybee Brain. Neurosci. Lett. 2006, 398, 274–279. [Google Scholar] [CrossRef]
- Xia, S.; Miyashita, T.; Fu, T.F.; Lin, W.Y.; Wu, C.L.; Pyzocha, L.; Lin, I.R.; Saitoe, M.; Tully, T.; Chiang, A.S. NMDA Receptors Mediate Olfactory Learning and Memory in Drosophila. Curr. Biol. 2005, 15, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.J.; Kohn, A.B.; Bobkova, Y.V.; Moroz, L.L. Molecular Characterization of NMDA-like Receptors in Aplysia and Lymnaea: Relevance to Memory Mechanisms. Biol. Bull. 2006, 210, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Chiang, A.S.; Lin, W.Y.; Liu, H.P.; Pszczolkowski, M.A.; Fu, T.F.; Chiu, S.L.; Holbrook, G.L. Insect NMDA Receptors Mediate Juvenile Hormone Biosynthesis. Proc. Natl. Acad. Sci. USA 2002, 99, 37–42. [Google Scholar] [CrossRef]
- Cattaert, D.; Birman, S. Blockade of the Central Generator of Locomotor Rhythm by Noncompetitive NMDA Receptor Antagonists in Drosophila Larvae. J. Neurobiol. 2001, 48, 58–73. [Google Scholar] [CrossRef]
- Brockie, P.J.; Mellem, J.E.; Hills, T.; Madsen, D.M.; Maricq, A.V. The C. Elegans Glutamate Receptor Subunit NMR-1 Is Required for Slow NMDA-Activated Currents That Regulate Reversal Frequency during Locomotion. Neuron 2001, 31, 617–630. [Google Scholar] [CrossRef]
- McDiarmid, T.A.; Bernardos, A.C.; Rankin, C.H. Habituation Is Altered in Neuropsychiatric Disorders—A Comprehensive Review with Recommendations for Experimental Design and Analysis. Neurosci. Biobehav. Rev. 2017, 80, 286–305. [Google Scholar] [CrossRef]
- van Alphen, B.; van Swinderen, B. Drosophila Strategies to Study Psychiatric Disorders. Brain Res. Bull. 2013, 92, 1–11. [Google Scholar] [CrossRef]
- Moulin, T.C.; Covill, L.E.; Itskov, P.M.; Williams, M.J.; Schiöth, H.B. Rodent and Fly Models in Behavioral Neuroscience: An Evaluation of Methodological Advances, Comparative Research, and Future Perspectives. Neurosci. Biobehav. Rev. 2021, 120, 1–12. [Google Scholar] [CrossRef]
- Chang, X.; Lima, L.d.A.; Liu, Y.; Li, J.; Li, Q.; Sleiman, P.M.A.; Hakonarson, H. Common and Rare Genetic Risk Factors Converge in Protein Interaction Networks Underlying Schizophrenia. Front. Genet. 2018, 9, 434. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiöth, H.B.; Donzelli, L.; Arvidsson, N.; Williams, M.J.; Moulin, T.C. Evidence for Prepulse Inhibition of Visually Evoked Motor Response in Drosophila melanogaster. Biology 2023, 12, 635. https://doi.org/10.3390/biology12040635
Schiöth HB, Donzelli L, Arvidsson N, Williams MJ, Moulin TC. Evidence for Prepulse Inhibition of Visually Evoked Motor Response in Drosophila melanogaster. Biology. 2023; 12(4):635. https://doi.org/10.3390/biology12040635
Chicago/Turabian StyleSchiöth, Helgi B., Laura Donzelli, Nicklas Arvidsson, Michael J. Williams, and Thiago C. Moulin. 2023. "Evidence for Prepulse Inhibition of Visually Evoked Motor Response in Drosophila melanogaster" Biology 12, no. 4: 635. https://doi.org/10.3390/biology12040635