
TRPC4 Channel Knockdown in the Hippocampal CA1 Region Impairs Modulation of Beta Oscillations in Novel Context
 , ,
, ,  and
and 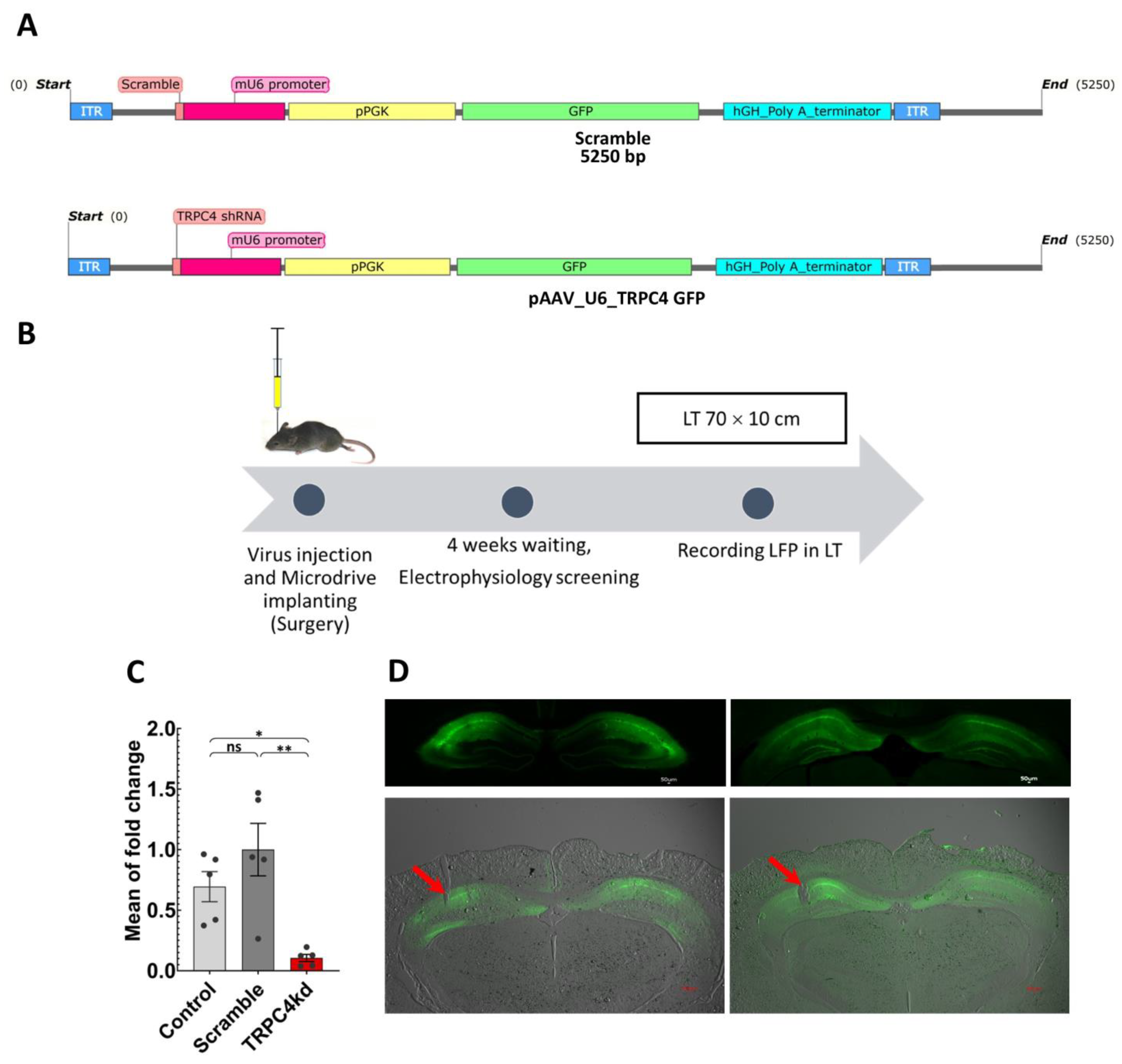{kind=link}
{kind=link}
{kind=link}
{kind=link}
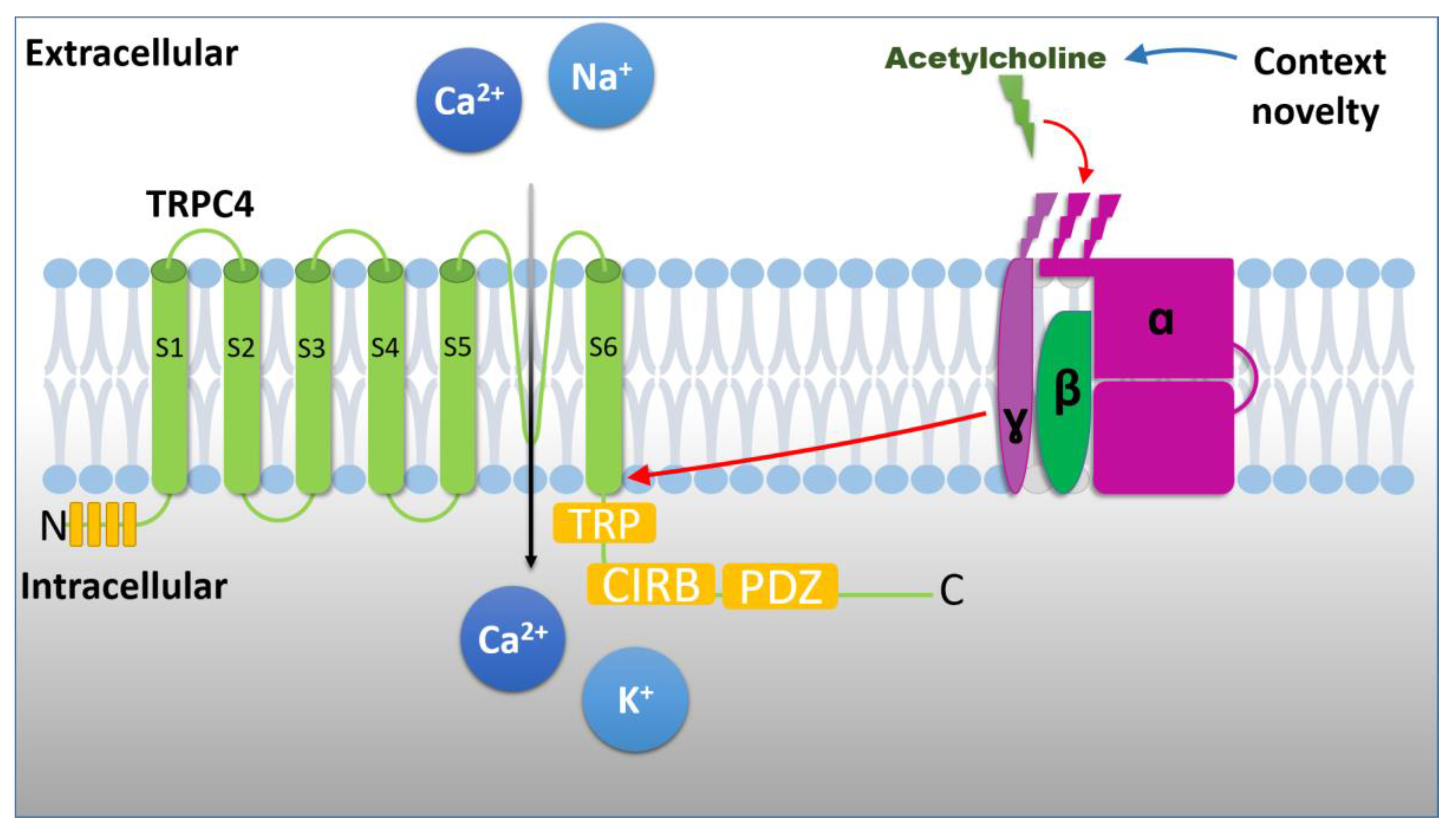{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Developing TRPC4 shRNA and Scramble Viruses
2.2. Animals
2.3. Virus Injection and Surgery
2.4. Building Tetrodes and In Vivo Electrophysiology Recording
2.5. Novel Environment
2.6. Histology
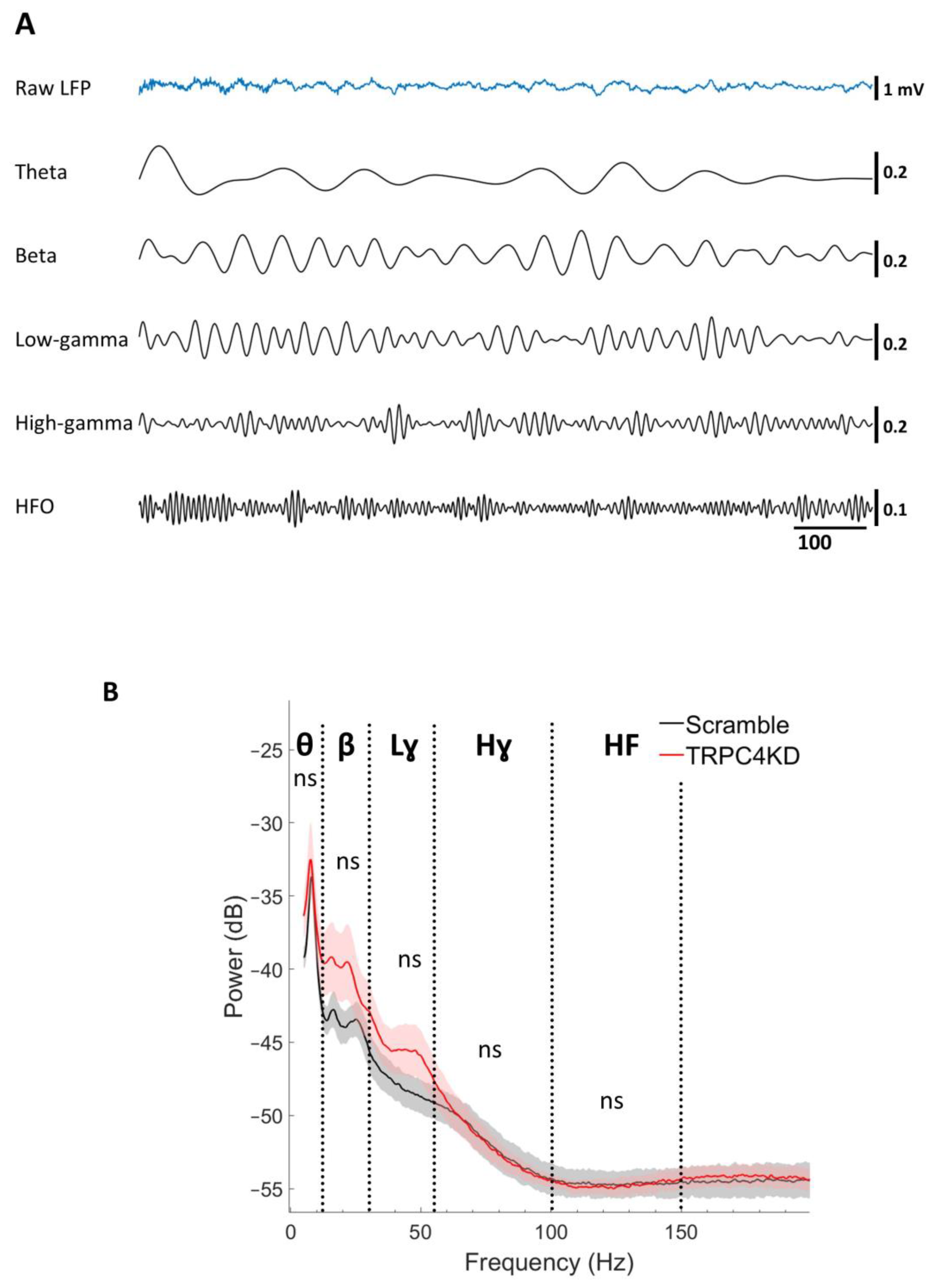
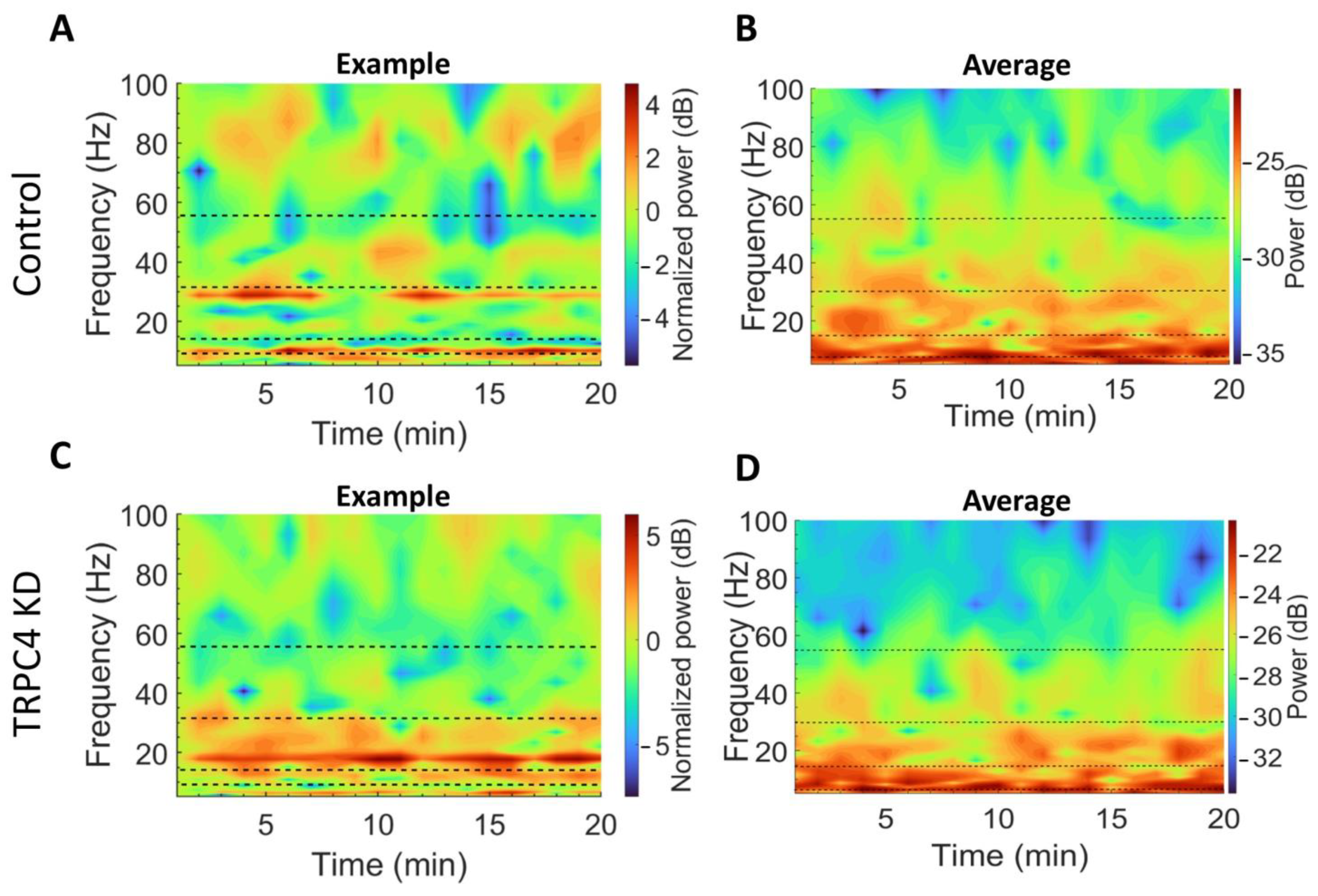
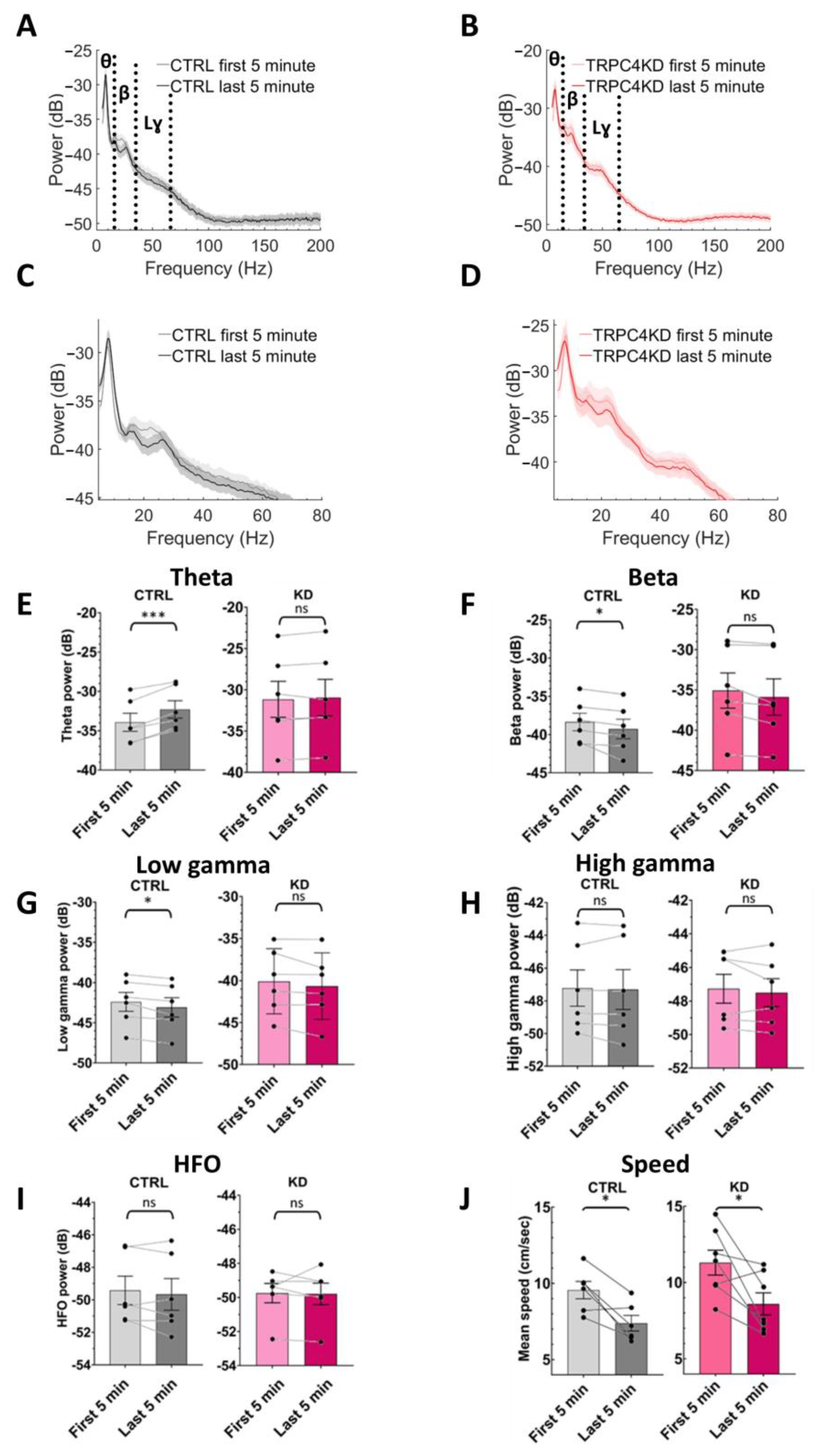
2.7. Data Analysis
3. Results
3.1. TRPC4 Knockdown
3.2. LFP Power Spectrum Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Preston, A.R.; Eichenbaum, H. Interplay of hippocampus and prefrontal cortex in memory. Curr. Biol. 2013, 23, R764–R773. [Google Scholar] [CrossRef]
- Kupferschmidt, D.A.; Gordon, J.A. The dynamics of disordered dialogue: Prefrontal, hippocampal and thalamic miscommunication underlying working memory deficits in schizophrenia. Brain Neurosci. Adv. 2018, 2, 239821281877182. [Google Scholar] [CrossRef] [PubMed]
- Golledge, R.G. Cognitive Maps. In Encyclopedia of Social Measurement; Elsevier Inc.: Amsterdam, The Netherlands, 2004; pp. 329–339. [Google Scholar]
- Eichenbaum, H. The role of the hippocampus in navigation is memory. J. Neurophysiol. Am. Physiol. Soc. 2017, 117, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, S.M.; Buzsáki, G. Gamma oscillations dynamically couple hippocampal CA3 and CA1 regions during memory task performance. Proc. Natl. Acad. Sci. USA 2007, 104, 14495–14500. [Google Scholar] [CrossRef]
- Larkin, M.C.; Lykken, C.; Tye, L.D.; Wickelgren, J.G.; Frank, L.M. Hippocampal output area CA1 broadcasts a generalized novelty signal during an object-place recognition task. Hippocampus 2014, 24, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Kafkas, A.; Montaldi, D. Expectation affects learning and modulates memory experience at retrieval. Cognition 2018, 180, 123–134. Available online: https://pubmed.ncbi.nlm.nih.gov/30053569/ (accessed on 17 March 2023). [CrossRef]
- Buzsáki, G.; Draguhn, A. Neuronal oscillations in cortical networks. Science 2004, 304, 1926–1929. Available online: https://pubmed.ncbi.nlm.nih.gov/15218136/ (accessed on 20 February 2023). [CrossRef]
- Buzśaki, G.; Wang, X.J. Mechanisms of Gamma Oscillations. Annu. Rev. Neurosci. 2012, 35, 203. [Google Scholar] [CrossRef]
- França, A.S.C.; Borgesius, N.Z.; Souza, B.C.; Cohen, M.X. Beta2 Oscillations in Hippocampal-Cortical Circuits during Novelty Detection. Front. Syst. Neurosci. 2021, 15, 8. [Google Scholar]
- Berke, J.D.; Hetrick, V.; Breck, J.; Greene, R.W. Transient 23–30 Hz oscillations in mouse hippocampus during exploration of novel environments. Hippocampus 2008, 18, 519–529. [Google Scholar] [CrossRef]
- Iwasaki, S.; Sasaki, T.; Ikegaya, Y. Hippocampal beta oscillations predict mouse object-location associative memory performance. Hippocampus 2021, 31, 503–511. [Google Scholar] [CrossRef]
- Jiang, H.; Bahramisharif, A.; van Gerven, M.A.J.; Jensen, O. Distinct directional couplings between slow and fast gamma power to the phase of theta oscillations in the rat hippocampus. Eur. J. Neurosci. 2020, 51, 2070–2081. [Google Scholar] [CrossRef] [PubMed]
- Kitanishi, T.; Ujita, S.; Fallahnezhad, M.; Kitanishi, N.; Ikegaya, Y.; Tashiro, A. Novelty-Induced Phase-Locked Firing to Slow Gamma Oscillations in the Hippocampus: Requirement of Synaptic Plasticity. Neuron 2015, 86, 1265–1276. [Google Scholar] [CrossRef]
- Gattas, S.; Elias, G.A.; Janecek, J.; Yassa, M.A.; Fortin, N.J. Proximal CA1 20-40 Hz power dynamics reflect trial-specific information processing supporting nonspatial sequence memory. Elife 2022, 11, e55528. [Google Scholar]
- Duszkiewicz, A.J.; McNamara, C.G.; Takeuchi, T.; Genzel, L. Novelty and Dopaminergic Modulation of Memory Persistence: A Tale of Two Systems. Trends Neurosci. 2019, 42, 102. [Google Scholar] [CrossRef] [PubMed]
- Barry, C.; Heys, J.G.; Hasselmo, M.E. Possible role of acetylcholine in regulating spatial novelty effects on theta rhythm and grid cells. Front. Neural Circuits 2012, 6, 5. [Google Scholar] [CrossRef]
- Marrosu, F.; Portas, C.; Mascia, M.S.; Casu, M.A.; Fà, M.; Giagheddu, M.; Imperato, A.; Gessa, G.L. Microdialysis measurement of cortical and hippocampal acetylcholine release during sleep-wake cycle in freely moving cats. Brain Res. 1995, 671, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Hasselmo, M.E. Neuromodulation and the hippocampus: Memory function and dysfunction in a network simulation. Prog. Brain Res. 1999, 121, 3–18. [Google Scholar] [PubMed]
- Meeter, M.; Murre, J.M.J.; Talamini, L.M. Mode shifting between storage and recall based on novelty detection in oscillating hippocampal circuits. Hippocampus 2004, 14, 722–741. [Google Scholar] [CrossRef]
- Arai, J.; Natsume, K. The properties of carbachol-induced beta oscillation in rat hippocampal slices. Neurosci. Res. 2006, 54, 95–103. [Google Scholar] [CrossRef]
- Fellous, J.M.; Sejnowski, T.J. Cholinergic Induction of Oscillations in the Hippocampal Slice in the Slow (0.5–2 Hz), Theta (5–12 Hz), and Gamma (35–70 Hz) Bands. Hippocampus 2000, 10, 187–197. [Google Scholar] [CrossRef]
- Gualdani, R.; Gailly, P. How TRPC channels modulate hippocampal function. Int. J. Mol. Sci. 2020, 21, 3915. Available online: https://pubmed.ncbi.nlm.nih.gov/32486187/ (accessed on 27 January 2022). [CrossRef]
- Valero-Aracama, M.J.; Reboreda, A.; Arboit, A.; Sauvage, M.; Yoshida, M. Noradrenergic Suppression of Persistent Firing in Hippocampal CA1 Pyramidal Cells through cAMP-PKA Pathway. eNeuro 2021, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, Y.; Oleinikov, K.; Schindeldecker, B.; Wyatt, A.; Weißgerber, P.; Flockerzi, V.; Boehm, U.; Freichel, M.; Bruns, D. TRPC channels regulate Ca2+-signaling and short-term plasticity of fast glutamatergic synapses. PLoS Biol. 2019, 17, e3000445. [Google Scholar] [CrossRef] [PubMed]
- Bröker-Lai, J.; Kollewe, A.; Schindeldecker, B.; Pohle, J.; Nguyen Chi, V.; Mathar, I.; Guzman, R.; Schwarz, Y.; Lai, A.; Weißgerber, P.; et al. Heteromeric channels formed by TRPC1, TRPC4 and TRPC5 define hippocampal synaptic transmission and working memory. EMBO J. 2017, 36, 2770–2789. Available online: http://www.ncbi.nlm.nih.gov/pubmed/28790178 (accessed on 23 March 2018). [CrossRef] [PubMed]
- Chen, X.; Sooch, G.; Demaree, I.S.; White, F.A.; Obukhov, A.G. Transient Receptor Potential Canonical (TRPC) Channels: Then and Now. Cells 2020, 9, 1983. [Google Scholar] [CrossRef]
- França, A.S.C.; do Nascimento, G.C.; Lopes-dos-Santos, V.; Muratori, L.; Ribeiro, S.; Lobão-Soares, B.; Tort, A.B. Beta2 oscillations (23–30 Hz) in the mouse hippocampus during novel object recognition. Eur. J. Neurosci. 2014, 40, 3693–3703. [Google Scholar] [CrossRef]
- Lepannetier, S.; Gualdani, R.; Tempesta, S.; Schakman, O.; Seghers, F.; Kreis, A.; Yerna, X.; Slimi, A.; de Clippele, M.; Tajeddine, N.; et al. Activation of TRPC1 Channel by Metabotropic Glutamate Receptor mGluR5 Modulates Synaptic Plasticity and Spatial Working Memory. Front. Cell. Neurosci. 2018, 12, 318. [Google Scholar] [CrossRef]
- Mitlöhner, J.; Kaushik, R.; Niekisch, H.; Blondiaux, A.; Gee, C.E.; Happel, M.F.K.; Gundelfinger, E.; Dityatev, A.; Frischknecht, R.; Seidenbecher, C. Dopamine Receptor Activation Modulates the Integrity of the Perisynaptic Extracellular Matrix at Excitatory Synapses. Cells 2020, 9, 260. [Google Scholar] [CrossRef]
- Paxinos, G.; Keith, B.J.; Franklin, M.A.P. The Mouse Brain in Stereotaxic Coordinates, 3rd ed.; Elsevier Science: Amsterdam, The Netherlands, 2008; 360p. [Google Scholar]
- Kadir, S.N.; Goodman, D.F.M.; Harris, K.D. High-dimensional cluster analysis with the masked EM algorithm. Neural Comput. 2014, 26, 2379–2394. Available online: https://pubmed.ncbi.nlm.nih.gov/25149694/ (accessed on 21 March 2023). [CrossRef]
- Buzsáki, G.; Buhl, D.L.; Harris, K.D.; Csicsvari, J.; Czéh, B.; Morozov, A. Hippocampal network patterns of activity in the mouse. Neuroscience 2003, 116, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.J.; Sweet, T.B.; Clapham, D.E. International Union of Basic and Clinical Pharmacology. LXXVI. Current progress in the Mammalian TRP ion channel family. Pharmacol. Reviews Am. Soc. Pharmacol. Exp. Ther. 2010, 62, 381–404. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, M.G.; Rakovska, A.; Benton, R.S.; Pazzagli, M.; Bianchi, L.; Pepeu, G. Effects of novelty and habituation on acetylcholine, GABA, and glutamate release from the frontal cortex and hippocampus of freely moving rats. Neuroscience 2001, 106, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Acquas, E.; Wilson, C.; Fibiger, H.C. Conditioned and Unconditioned Stimuli Increase Frontal Cortical and Hippocampal Acetylcholine Release: Effects of Novelty, Habituation, and Fear. J. Neurosci. 1996, 16, 3089–3096. Available online: https://www.jneurosci.org/content/16/9/3089 (accessed on 28 February 2023). [CrossRef]
- Shimono, K.; Brucher, F.; Granger, R.; Lynch, G.; Taketani, M. Origins and Distribution of Cholinergically Induced β Rhythms in Hippocampal Slices. J. Neurosci. 2000, 20, 8462–8473. Available online: https://www.jneurosci.org/content/20/22/8462 (accessed on 27 February 2023). [CrossRef]
- Bibbig, A.; Middleton, S.; Racca, C.; Gillies, M.J.; Garner, H.; LeBeau, F.E.N.; Davies, C.H.; Whittington, M.A. Beta rhythms (15–20 Hz) generated by nonreciprocal communication in hippocampus. J. Neurophysiol. 2007, 97, 2812–2823. [Google Scholar] [CrossRef]
- Jeewajee, A.; Lever, C.; Burton, S.; O’Keefe, J.; Burgess, N. Environmental novelty is signaled by reduction of the hippocampal theta frequency. Hippocampus 2008, 18, 340–348. [Google Scholar] [CrossRef]
- Grossberg, S. Beta oscillations and hippocampal place cell learning during exploration of novel environments. Hippocampus 2009, 19, 881–885. [Google Scholar] [CrossRef]
- Hasselmo, M.E. Neuromodulation: Acetylcholine and memory consolidation. Trends Cogn. Sci. 1999, 3, 351–359. Available online: https://pubmed.ncbi.nlm.nih.gov/10461198/ (accessed on 17 March 2023). [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saber Marouf, B.; Reboreda, A.; Theissen, F.; Kaushik, R.; Sauvage, M.; Dityatev, A.; Yoshida, M. TRPC4 Channel Knockdown in the Hippocampal CA1 Region Impairs Modulation of Beta Oscillations in Novel Context. Biology 2023, 12, 629. https://doi.org/10.3390/biology12040629
Saber Marouf B, Reboreda A, Theissen F, Kaushik R, Sauvage M, Dityatev A, Yoshida M. TRPC4 Channel Knockdown in the Hippocampal CA1 Region Impairs Modulation of Beta Oscillations in Novel Context. Biology. 2023; 12(4):629. https://doi.org/10.3390/biology12040629
Chicago/Turabian StyleSaber Marouf, Babak, Antonio Reboreda, Frederik Theissen, Rahul Kaushik, Magdalena Sauvage, Alexander Dityatev, and Motoharu Yoshida. 2023. "TRPC4 Channel Knockdown in the Hippocampal CA1 Region Impairs Modulation of Beta Oscillations in Novel Context" Biology 12, no. 4: 629. https://doi.org/10.3390/biology12040629