
N20D/N116E Combined Mutant Downward Shifted the pH Optimum of Bacillus subtilis NADH Oxidase


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Site-Directed Mutagenesis
2.3. Enzyme Preparation, Purification, and Identification
2.4. Activity and Optimum pH Determination
2.5. Catalytic Performance of Enzymes Mixture
2.6. ROSETTA Supercharge, Modeling and Molecular Dynamics Simulation
3. Results
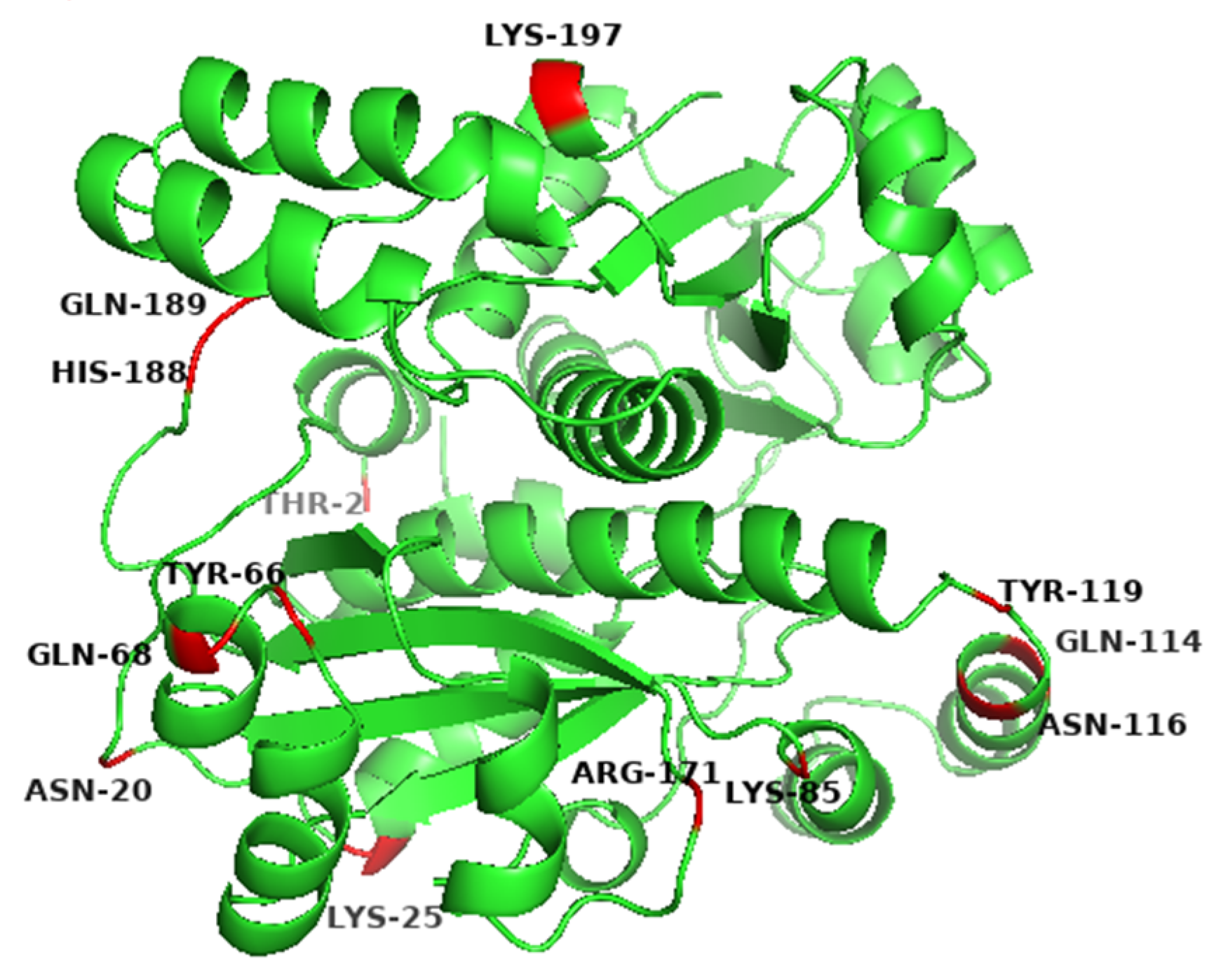
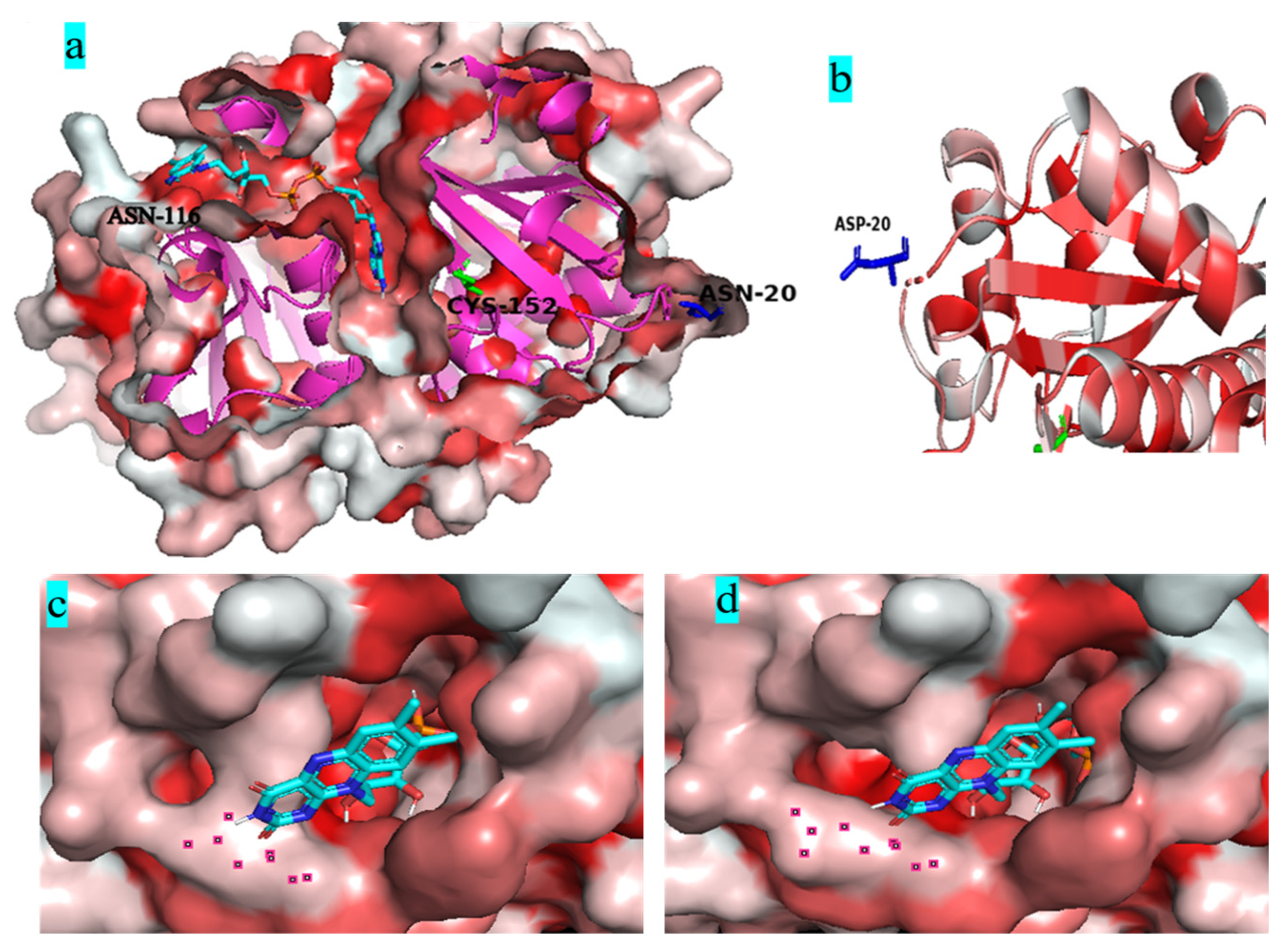
3.1. Screening of Residues on the Surface of BsNox for Site-Directed Mutagenesis
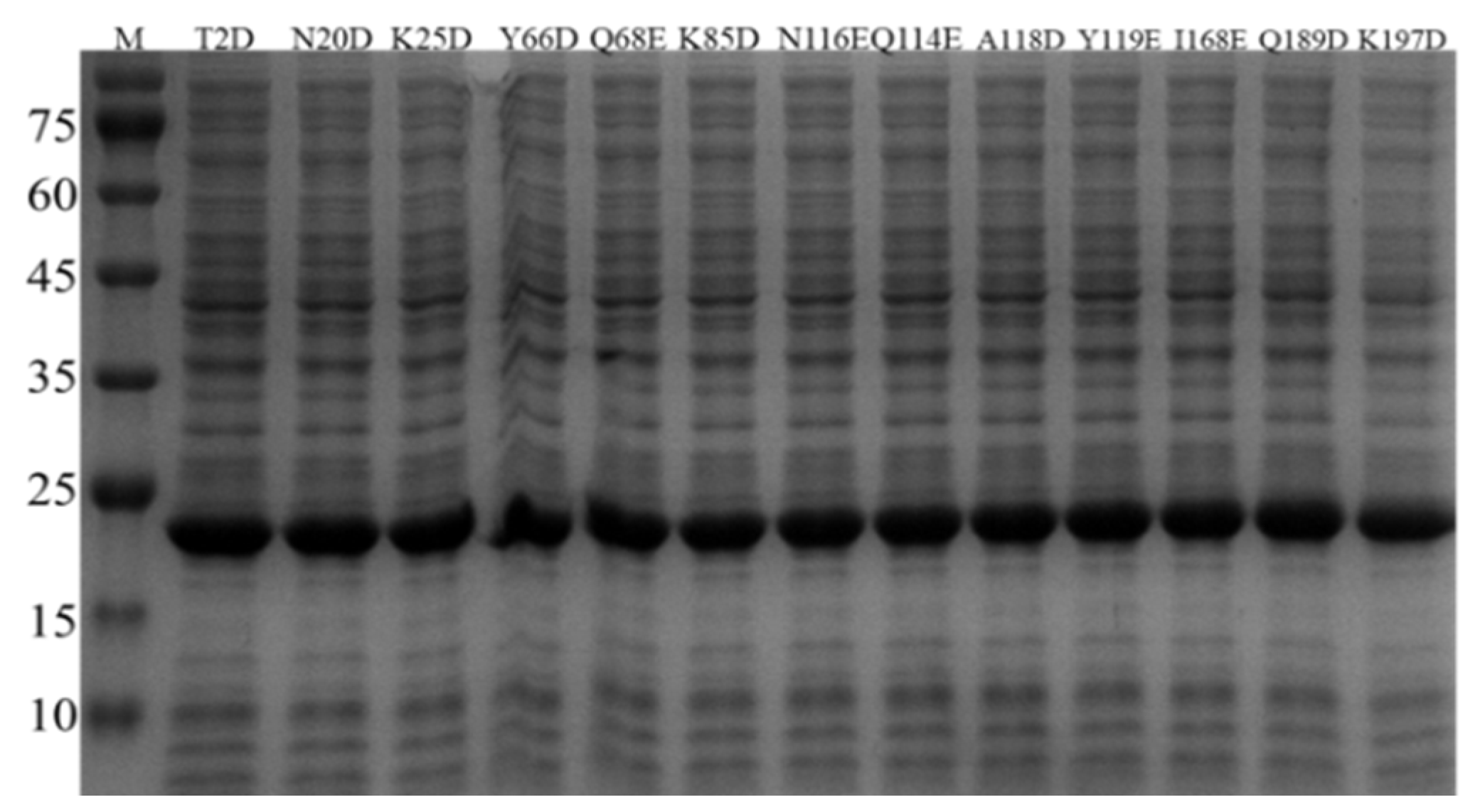
3.2. Expression of BsNox and Its Mutants
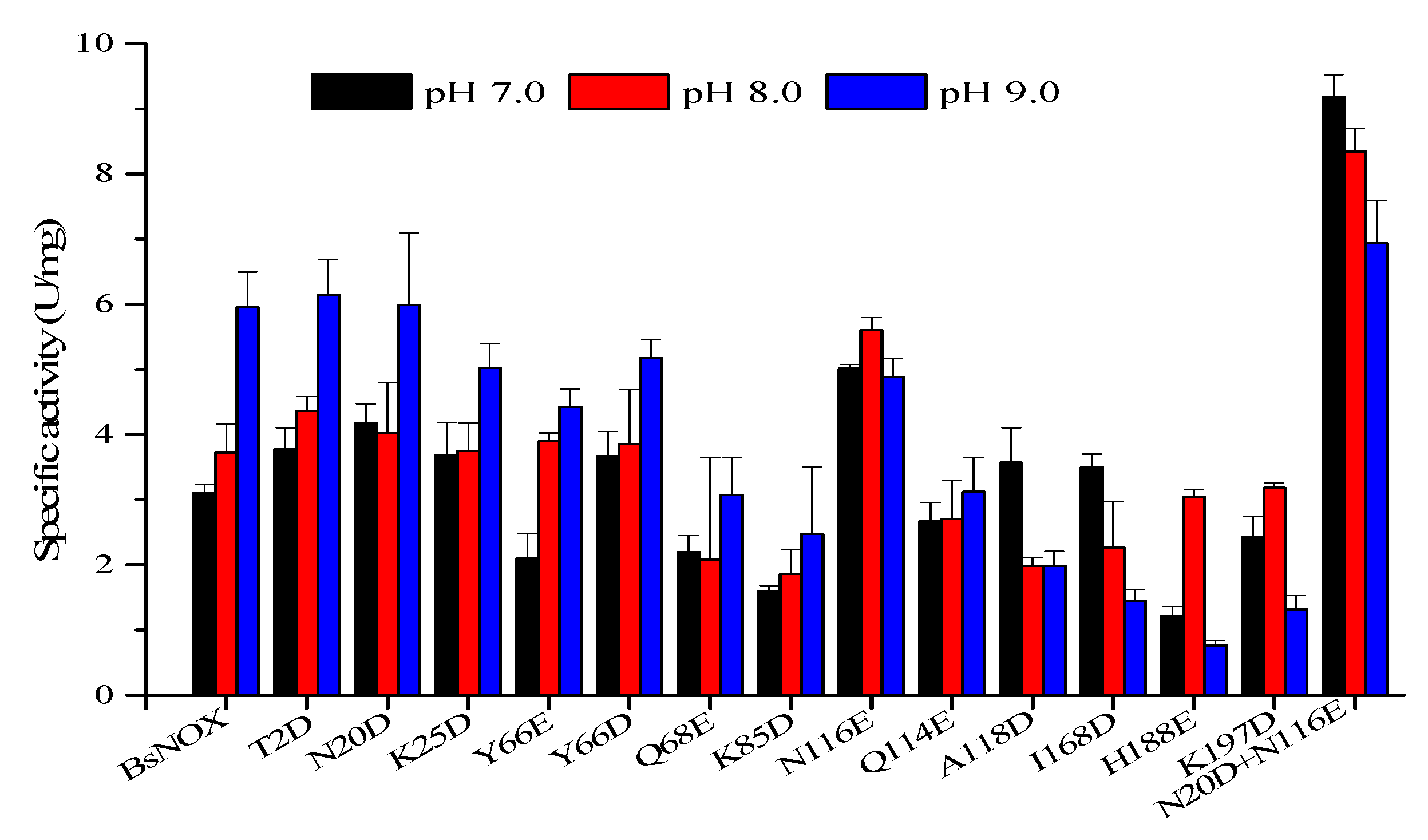
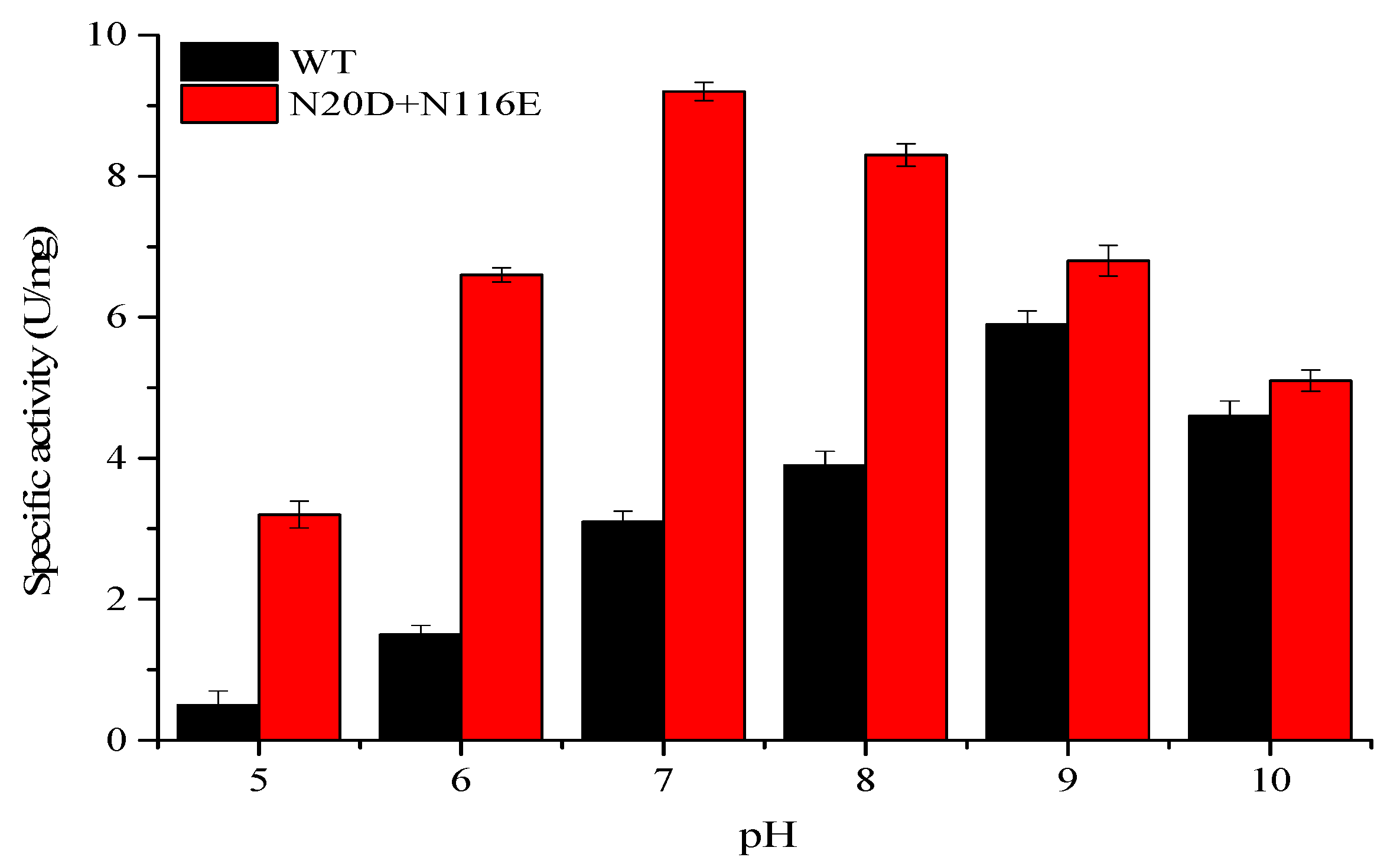
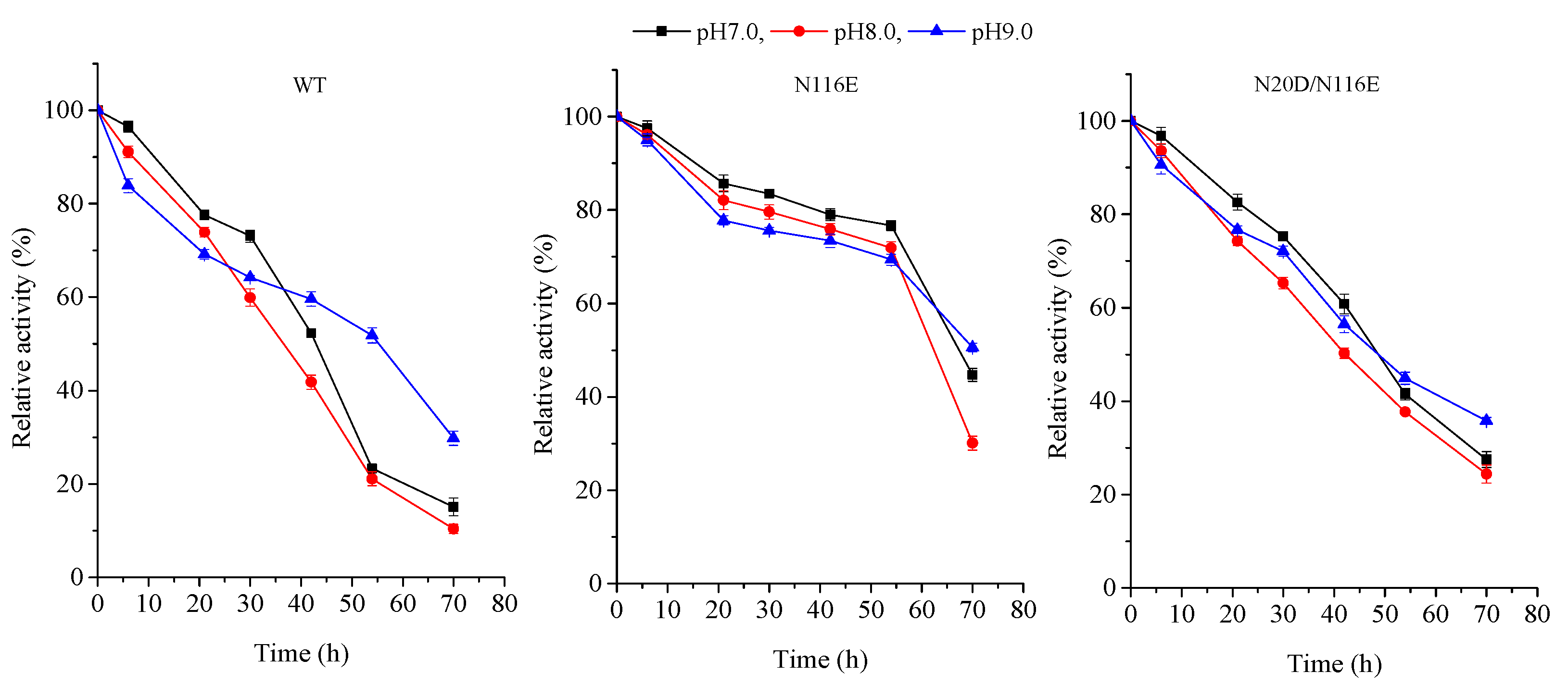
3.3. pH Profile and Stability
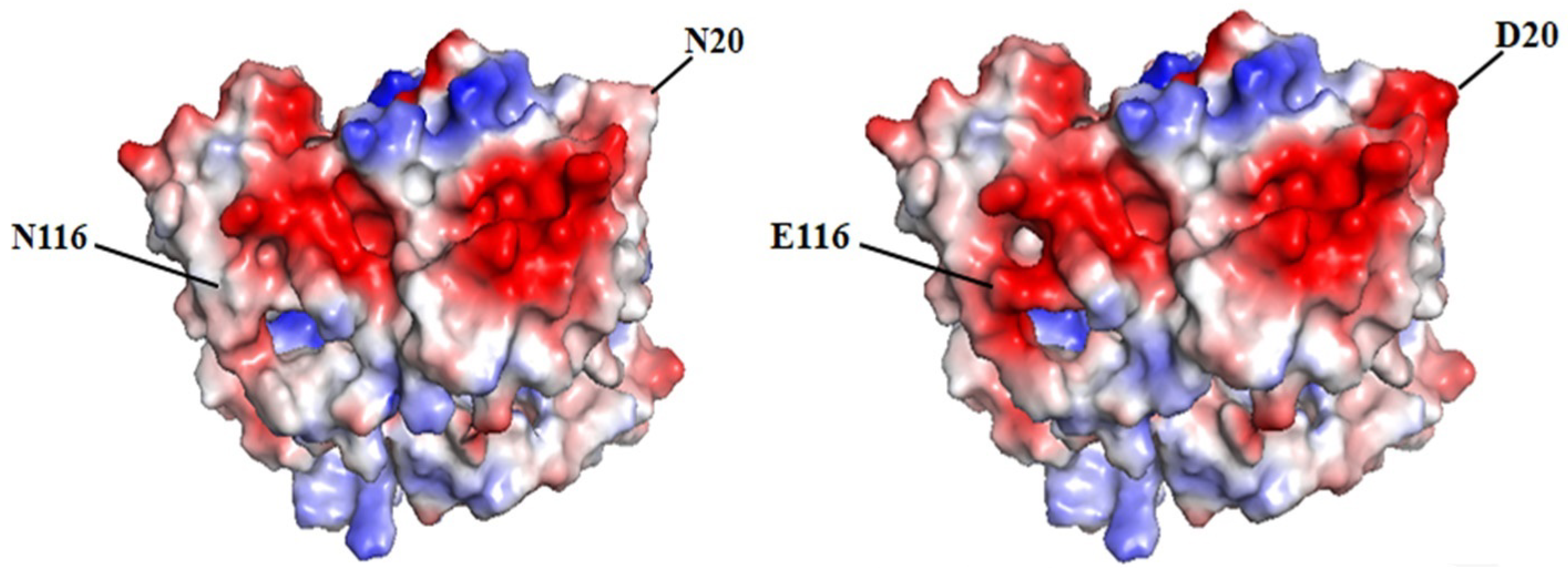
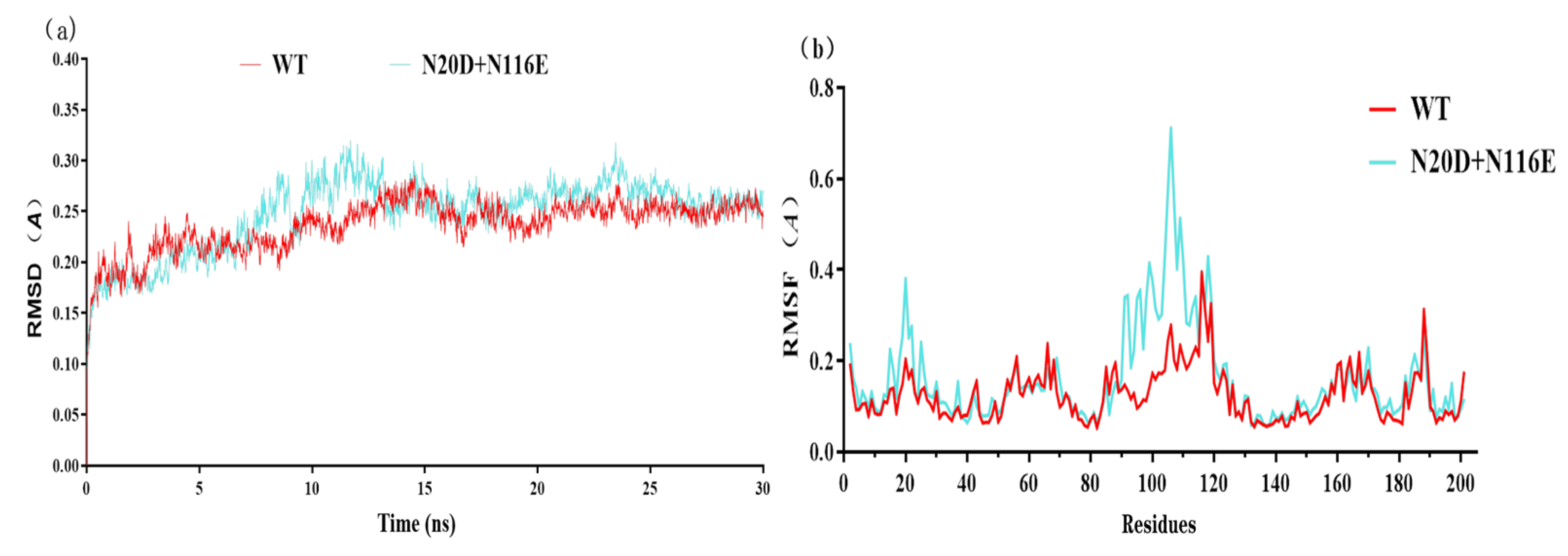
3.4. Molecular Dynamics Simulation
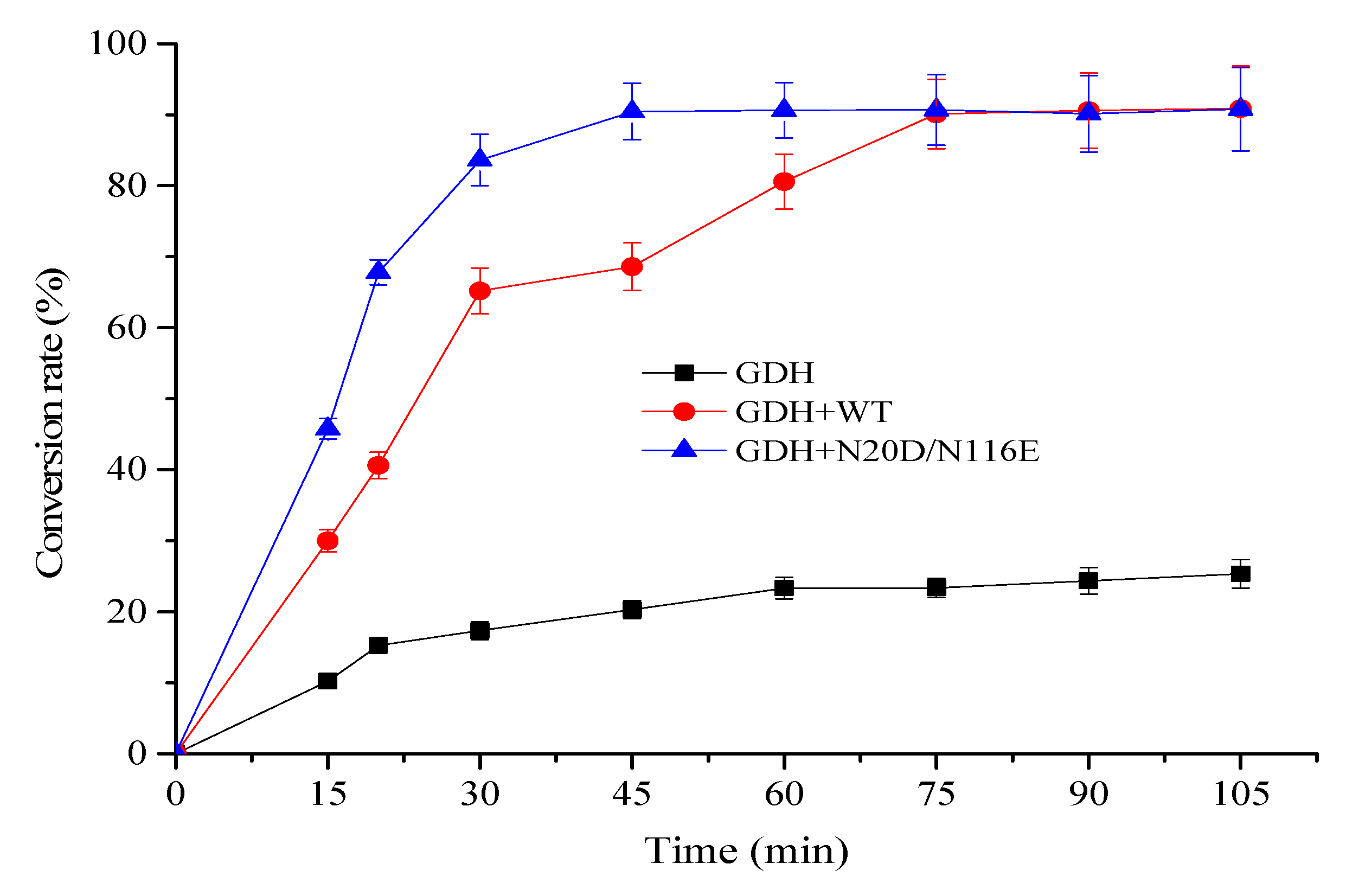
3.5. Using Mutant N20D/N116E for NAD+ Regeneration Enhanced α-Ketoglutarate Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miller, D.C.; Athavale, S.V.; Arnold, F.H. Combining chemistry and protein engineering for new-to-nature biocatalysis. Nat. Synth. 2022, 1, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, A.R.; Dominguez de Maria, P.; Littlechild, J.A.; Schurmann, M.; Sheldon, R.A.; Wohlgemuth, R. Biocatalysis as key to sustainable industrial chemistry. ChemSusChem 2022, 15, e202102709. [Google Scholar] [PubMed]
- Sheldon, R.A.; Brady, D. Green chemistry, biocatalysis, and the chemical industry of the future. ChemSusChem 2022, 15, e202102628. [Google Scholar] [CrossRef]
- Torrelo, G.; Hanefeld, U.; Hollmann, F. Biocatalysis. Catal. Lett. 2015, 145, 309–345. [Google Scholar] [CrossRef]
- Tassano, E.; Hall, M. Enzymatic self-sufficient hydride transfer processes. Chem. Soc. Rev. 2019, 48, 5596–5615. [Google Scholar] [CrossRef] [PubMed]
- Hollmann, F.; Opperman, D.J.; Paul, C.E. Biocatalytic reduction reactions from a chemist’s perspective. Angew. Chem. Int. Ed. Engl. 2021, 60, 5644–5665. [Google Scholar] [CrossRef]
- Ding, Y.; Li, X.; Horsman, G.P.; Li, P.; Wang, M.; Li, J.; Zhang, Z.; Liu, W.; Wu, B.; Tao, Y.; et al. Construction of an Alternative NAD(+) De Novo Biosynthesis Pathway. Adv. Sci. 2021, 8, 2004632. [Google Scholar] [CrossRef]
- Wang, M.; Chen, B.; Fang, Y.; Tan, T. Cofactor engineering for more efficient production of chemicals and biofuels. Biotechnol. Adv. 2017, 35, 1032–1039. [Google Scholar] [CrossRef]
- Zheng, Y.G.; Yin, H.H.; Yu, D.F.; Chen, X.; Tang, X.L.; Zhang, X.J.; Xue, Y.P.; Wang, Y.J.; Liu, Z.Q. Recent advances in biotechnological applications of alcohol dehydrogenases. Appl. Microbiol. Biotechnol. 2017, 101, 987–1001. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, S.; Wang, P. Nanoparticle-supported multi-enzyme biocatalysis with in situ cofactor regeneration. J. Biotechnol. 2009, 139, 102–107. [Google Scholar] [CrossRef]
- El-Zahab, B.; Jia, H.; Wang, P. Enabling multienzyme biocatalysis using nanoporous materials. Biotechnol. Bioeng. 2004, 87, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yang, T.; Zhou, J.; Xu, M.; Zhang, X.; Rao, Z. Elimination of a free cysteine by creation of a disulfide bond increases the activity and stability of Candida boidinii formate dehydrogenase. Appl. Environ. Microbiol. 2017, 83, e02624-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, M.-Y.; Jiang, X.-P.; Ling, X.-M.; Xu, M.-Q.; Zhu, Y.-H.; Zhang, Y.-W. Immobilization of glycerol dehydrogenase and NADH oxidase for enzymatic synthesis of 1,3-dihydroxyacetone with in situ cofactor regeneration. J. Chem. Technol. Biotechnol. 2018, 93, 2351–2358. [Google Scholar] [CrossRef]
- Jia, B.; Park, S.C.; Lee, S.; Pham, B.P.; Yu, R.; Le, T.L.; Han, S.W.; Yang, J.K.; Choi, M.S.; Baumeister, W.; et al. Hexameric ring structure of a thermophilic archaeon NADH oxidase that produces predominantly H2O. FEBS J. 2008, 275, 5355–5366. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Li, J.; Sivakumar, D.; Kim, T.S.; Patel, S.K.S.; Kalia, V.C.; Kim, I.W.; Zhang, Y.W.; Lee, J.K. NADH oxidase from Lactobacillus reuteri: A versatile enzyme for oxidized cofactor regeneration. Int. J. Biol. Macromol. 2019, 123, 629–636. [Google Scholar] [CrossRef]
- Titov, D.V.; Cracan, V.; Goodman, R.P.; Peng, J.; Grabarek, Z.; Mootha, V.K. Complementation of mitochondrial electron transport chain by manipulation of the NAD+/NADH ratio. Science 2016, 352, 231–235. [Google Scholar] [CrossRef] [Green Version]
- Riebel, B.R.; Gibbs, P.R.; Wellborn, W.B.; Bommarius, A.S. Cofactor regeneration of both NAD+ from NADH and NADP+ from NADPH:NADH oxidase from Lactobacillus sanfranciscensis. Adv. Synth. Catal. 2003, 345, 707–712. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Tiwari, M.K.; Gao, H.; Dhiman, S.S.; Jeya, M.; Lee, J.K. Cloning and characterization of a thermostable H2O-forming NADH oxidase from Lactobacillus rhamnosus. Enzyme Microb. Technol. 2012, 50, 255–262. [Google Scholar] [CrossRef]
- Gao, H.; Tiwari, M.K.; Kang, Y.C.; Lee, J.-K. Characterization of H2O-forming NADH oxidase from Streptococcus pyogenes and its application in l-rare sugar production. Bioorg. Med. Chem. Lett. 2012, 22, 1931–1935. [Google Scholar] [CrossRef]
- Li, F.-L.; Shi, Y.; Zhang, J.-X.; Gao, J.; Zhang, Y.-W. Cloning, expression, characterization and homology modeling of a novel water-forming NADH oxidase from Streptococcus mutans ATCC 25175. Int. J. Biol. Macromol. 2018, 113, 1073–1079. [Google Scholar] [CrossRef]
- Shi, X.C.; Zou, Y.N.; Chen, Y.; Zheng, C.; Li, B.B.; Xu, J.H.; Shen, X.N.; Ying, H.J. A water-forming NADH oxidase regulates metabolism in anaerobic fermentation. Biotechnol. Biofuels 2016, 9, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.Q.; Li, F.L.; Yu, W.Q.; Li, R.F.; Zhang, Y.W. Combined cross-linked enzyme aggregates of glycerol dehydrogenase and NADH oxidase for high efficiency in situ NAD(+) regeneration. Int. J. Biol. Macromol. 2020, 144, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cui, Z.; Chang, H.; Fan, X.; Zhao, Q.; Wei, W. Conversion of glycerol to 1,3-dihydroxyacetone by glycerol dehydrogenase co-expressed with an NADH oxidase for cofactor regeneration. Biotechnol. Lett. 2016, 38, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Bian, L.; Huang, P.; Zhao, L.; Chen, Y.; Wu, X. Efficient whole-cell biosynthesis of l-gulose by coupling mannitol-1-dehydrogenase with NADH oxidase. Enzyme Microb. Technol. 2021, 148, 109815. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.G.; Moon, Y.M.; Choi, T.R.; Jung, H.R.; Yang, S.Y.; Ahn, J.O.; Joo, J.C.; Park, K.; Kim, Y.G.; Bhatia, S.K.; et al. Enhanced production of glutaric acid by NADH oxidase and GabD-reinforced bioconversion from l-lysine. Biotechnol. Bioeng. 2019, 116, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Su, W.B.; Li, F.L.; Li, X.Y.; Fan, X.M.; Liu, R.J.; Zhang, Y.W. Using galactitol dehydrogenase coupled with water-forming NADH oxidase for efficient enzymatic synthesis of L-tagatose. New Biotechnol. 2021, 62, 18–25. [Google Scholar] [CrossRef]
- Cui, Z.; Zhao, Y.; Mao, Y.; Shi, T.; Lu, L.; Ma, H.; Wang, Z.; Chen, T. In vitro biosynthesis of optically pure d-(−)- acetoin from meso-2,3-butanediol using 2,3-butanediol dehydrogenase and NADH oxidase. J. Chem. Technol. Biotechnol. 2019, 94, 2547–2554. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Bao, T.; Rao, Z.; Yang, T.; Xu, M.; Xu, Z.; Li, H.; Yang, S. The rebalanced pathway significantly enhances acetoin production by disruption of acetoin reductase gene and moderate-expression of a new water-forming NADH oxidase in Bacillus subtilis. Metab. Eng. 2014, 23, 34–41. [Google Scholar] [CrossRef]
- Yang, J.H.; Park, J.Y.; Kim, S.H.; Yoo, Y.J. Shifting pH optimum of Bacillus circulans xylanase based on molecular modeling. J. Biotechnol. 2008, 133, 294–300. [Google Scholar] [CrossRef]
- Pedersen, J.N.; Zhou, Y.; Guo, Z.; Perez, B. Genetic and chemical approaches for surface charge engineering of enzymes and their applicability in biocatalysis: A review. Biotechnol. Bioeng. 2019, 116, 1795–1812. [Google Scholar] [CrossRef]
- Der, B.S.; Kluwe, C.; Miklos, A.E.; Jacak, R.; Lyskov, S.; Gray, J.J.; Georgiou, G.; Ellington, A.D.; Kuhlman, B. Alternative computational protocols for supercharging protein surfaces for reversible unfolding and retention of stability. PLoS ONE 2013, 8, e64363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.; Mullaney, E.J.; Porres, J.M.; Roneker, K.R.; Crowe, S.; Rice, S.; Ko, T.; Ullah, A.H.J.; Daly, C.B.; Welch, R.; et al. Shifting the pH profile of Aspergillus niger PhyA Phytase to match the stomach pH enhances its effectiveness as an animal feed additive. Appl. Environ. Microbiol. 2006, 72, 4397–4403. [Google Scholar] [CrossRef] [Green Version]
- Pokhrel, S.; Joo, J.C.; Kim, Y.H.; Yoo, Y.J. Rational design of a Bacillus circulans xylanase by introducing charged residue to shift the pH optimum. Process Biochem. 2012, 47, 2487–2493. [Google Scholar] [CrossRef]
- Ma, F.; Xie, Y.; Luo, M.; Wang, S.; Hu, Y.; Liu, Y.; Feng, Y.; Yang, G.-Y. Sequence homolog-based molecular engineering for shifting the enzymatic pH optimum. Synth. Syst. Biotechnol. 2016, 1, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, M.; Yang, T.; Zhang, X.; Rao, Z. Surface charge-based rational design of aspartase modifies the optimal pH for efficient beta-aminobutyric acid production. Int. J. Biol. Macromol. 2020, 164, 4165–4172. [Google Scholar] [CrossRef]
- Sanchis, J.; Fernandez, L.; Carballeira, J.D.; Drone, J.; Gumulya, Y.; Hobenreich, H.; Kahakeaw, D.; Kille, S.; Lohmer, R.; Peyralans, J.J.; et al. Improved PCR method for the creation of saturation mutagenesis libraries in directed evolution: Application to difficult-to-amplify templates. Appl. Microbiol. Biotechnol. 2008, 81, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Z.; Wang, Y.M.; Li, X.; Zhu, M.C.; Zhang, H.W.; Xu, M.J.; Yang, T.W.; Rao, Z.M. Heterologous expression and rational design of l-asparaginase from Rhizomucor miehei to improve thermostability. Biology 2021, 10, 1346. [Google Scholar] [CrossRef]
- Kaufmann, K.W.; Lemmon, G.H.; DeLuca, S.L.; Sheehan, J.H.; Meiler, J. Practically useful: What the ROSETTA protein modeling suite can do for you. Biochemistry 2010, 49, 2987–2998. [Google Scholar] [CrossRef]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, R.; Haas, J.; Schwede, T. QMEANDisCo-distance constraints applied on model quality estimation. Bioinformatics 2020, 36, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Tiwari, M.K.; Singh, R.K.; Sung, B.H.; Kim, S.C.; Lee, J.K. Role of surface residue 184 in the catalytic activity of NADH oxidase from Streptococcus pyogenes. Appl. Microbiol. Biotechnol. 2014, 98, 7081–7088. [Google Scholar] [CrossRef]
- Wu, W.; Liu, F.; Wang, Y.; Yang, T.; Rao, Z. Surface charge engineering-based rational design of glutamate dehydrogenase from Bacillus subtilis modifies the optimal pH. Chin. J. Appl. Environ. Biol. 2022, 29, 7–14. (In Chinese) [Google Scholar]
- del Castillo-Olivares, A.; Yantiri, F.; Chueh, P.J.; Wang, S.; Sweeting, M.; Sedlak, D.; Morré, D.M.; Burgess, J.; Morré, D.J. A drug-responsive and protease-resistant peripheral NADH oxidase complex from the surface of HeLa S cells. Arch. Biochem. Biophys. 1998, 358, 125–140. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.; Pan, L.; Wu, W.; Pan, X.; Xu, M.; Zhang, X.; Rao, Z. N20D/N116E Combined Mutant Downward Shifted the pH Optimum of Bacillus subtilis NADH Oxidase. Biology 2023, 12, 522. https://doi.org/10.3390/biology12040522
Yang T, Pan L, Wu W, Pan X, Xu M, Zhang X, Rao Z. N20D/N116E Combined Mutant Downward Shifted the pH Optimum of Bacillus subtilis NADH Oxidase. Biology. 2023; 12(4):522. https://doi.org/10.3390/biology12040522
Chicago/Turabian StyleYang, Taowei, Longze Pan, Wenhui Wu, Xuewei Pan, Meijuan Xu, Xian Zhang, and Zhiming Rao. 2023. "N20D/N116E Combined Mutant Downward Shifted the pH Optimum of Bacillus subtilis NADH Oxidase" Biology 12, no. 4: 522. https://doi.org/10.3390/biology12040522