
Assessment of the Genetic Diversity and Population Structure of Rhizophora mucronata along Coastal Areas in Thailand
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
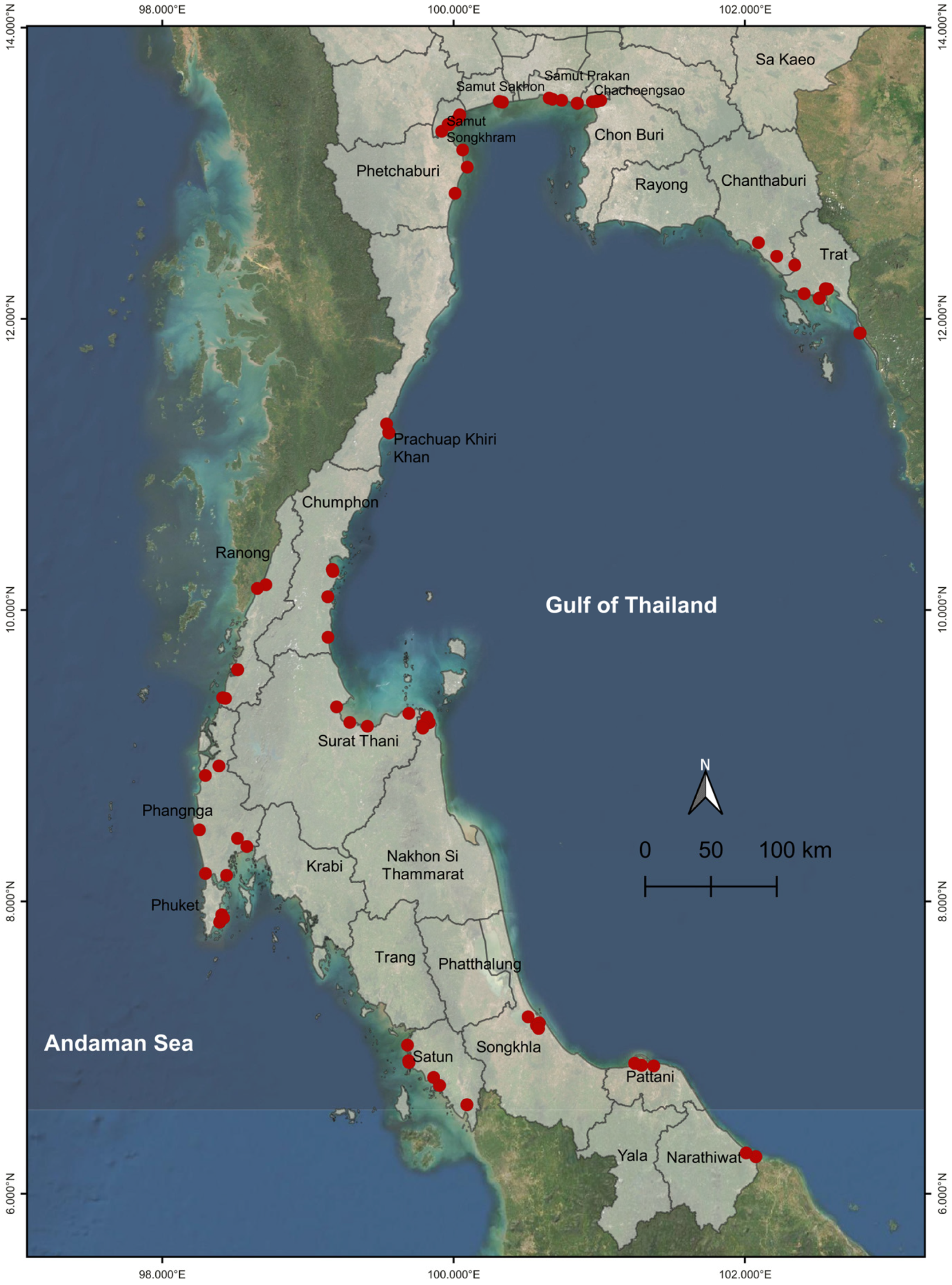
2.1. Plant Materials
2.2. DNA Extraction
2.3. RAD Library Preparation and Sequencing
2.4. Genome Assembly and Annotation
2.5. Repetitive Sequence Identification
2.6. Genome Assembly Quality Assessment
2.7. Single Nucleotide Polymorphisms Identification
2.8. Population Structure and Genetic Diversity
3. Results
3.1. Genome Assembly and Genome Annotation
3.2. Repeat Elements in the R. mucronata Genome
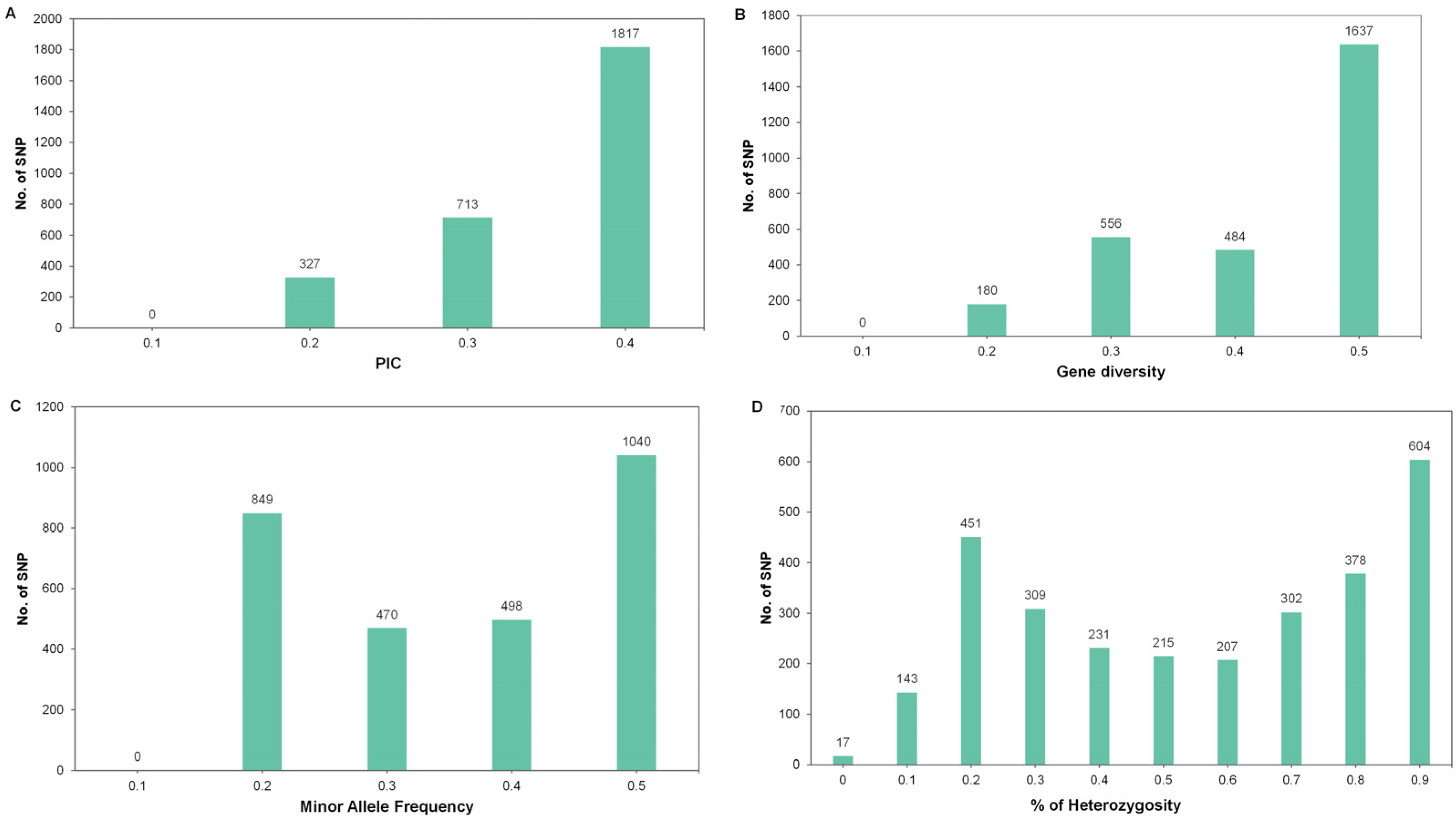
3.3. RAD-seq Data and SNP Identification
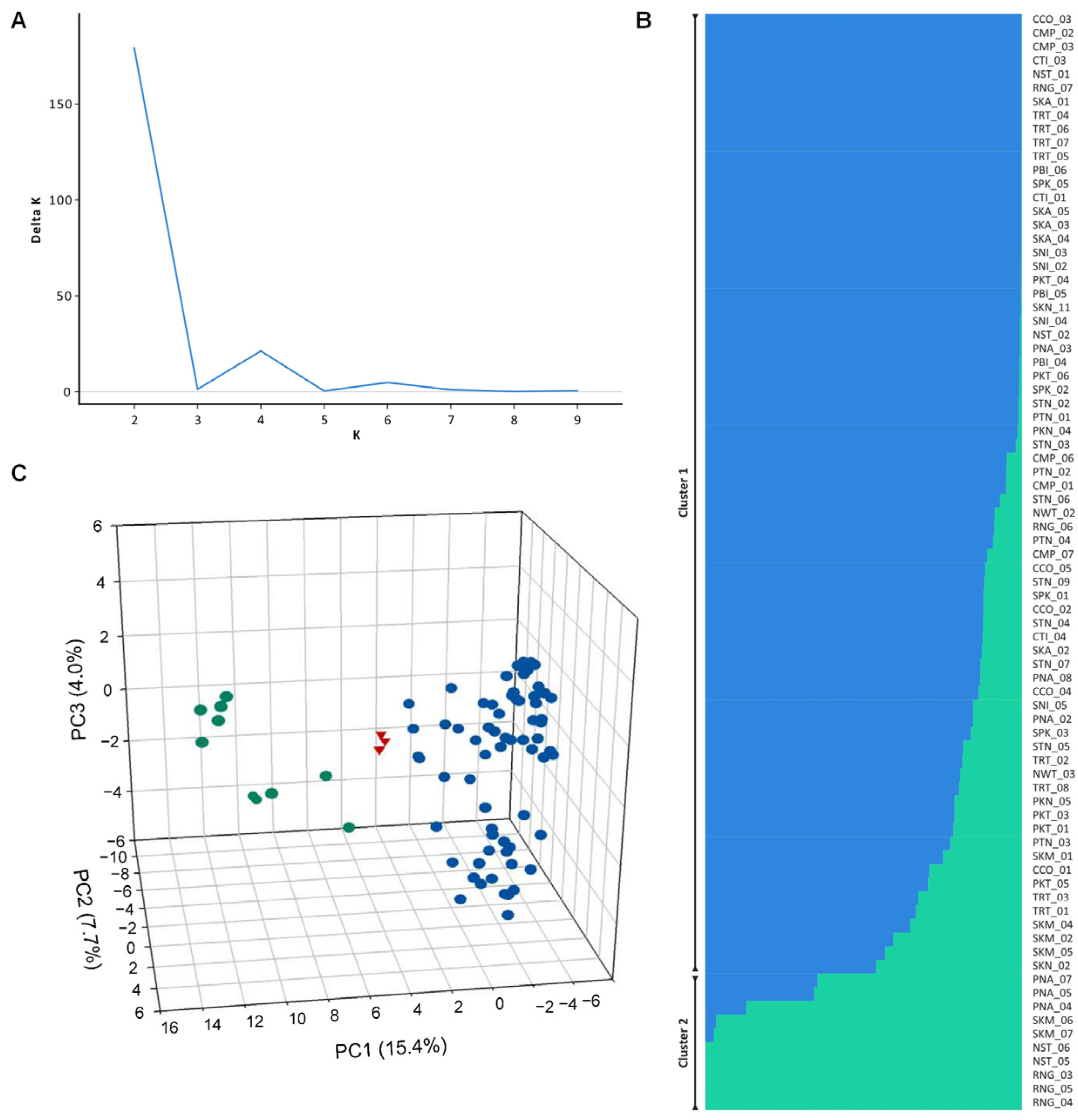
3.4. Population Genetic Structure
3.5. Genetic Differentiation of Populations
3.6. Genetic Diversity
4. Discussion
4.1. Genetic Structure and Genetic Differentiation
4.2. Genetic Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, S.Y.; Primavera, J.H.; Dahdouh-Guebas, F.; McKee, K.; Bosire, J.O.; Cannicci, S.; Diele, K.; Fromard, F.; Koedam, N.; Marchand, C.; et al. Ecological role and services of tropical mangrove ecosystems: A reassessment. Glob. Ecol. Biogeogr. 2014, 23, 726–743. [Google Scholar] [CrossRef]
- Walters, B.B.; Rönnbäck, P.; Kovacs, J.M.; Crona, B.; Hussain, S.A.; Badola, R.; Primavera, J.H.; Barbier, E.; Dahdouh-Guebas, F. Ethnobiology, socio-economics and management of mangrove forests: A review. Aquat. Bot. 2008, 89, 220–236. [Google Scholar] [CrossRef] [Green Version]
- Rippel, T.M.; Tomasula, J.; Murphy, S.M.; Wimp, G.M. Global change in marine coastal habitats impacts insect populations and communities. Curr. Opin. Insect Sci. 2021, 47, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gedan, K.B.; Kirwan, M.L.; Wolanski, E.; Barbier, E.B.; Silliman, B.R. The present and future role of coastal wetland vegetation in protecting shorelines: Answering recent challenges to the paradigm. Clim. Chang. 2011, 106, 7–29. [Google Scholar] [CrossRef]
- Bunting, P.; Rosenqvist, A.; Lucas, R.M.; Rebelo, L.-M.; Hilarides, L.; Thomas, N.; Hardy, A.; Itoh, T.; Shimada, M.; Finlayson, C.M. The Global Mangrove Watch—A New 2010 Global Baseline of Mangrove Extent. Remote Sens. 2018, 10, 1669. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global declines in human-driven mangrove loss. Glob. Change Biol. 2020, 26, 5844–5855. [Google Scholar] [CrossRef]
- Richards, D.R.; Friess, D.A. Rates and drivers of mangrove deforestation in Southeast Asia, 2000–2012. Proc. Natl. Acad. Sci. USA 2016, 113, 344–349. [Google Scholar] [CrossRef] [Green Version]
- Feller, I.C.; Friess, D.A.; Krauss, K.W.; Lewis, R.R. The state of the world’s mangroves in the 21st century under climate change. Hydrobiologia 2017, 803, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ragavan, P.; Kathiresan, K.; Zimmer, M.; Zhou, R.; Amir, A.A.; Mohan, P.; Rana, T. Three decades of global mangrove conservation—An overview. Malay. Nat. J. 2020, 72, 551–576. [Google Scholar]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C.; Farnsworth, E.J.; Fernando, E.S.; Kathiresan, K.; Koedam, N.E.; et al. The Loss of Species: Mangrove Extinction Risk and Geographic Areas of Global Concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Young, A.; Boyle, T.; Brown, T. The population genetic consequences of habitat fragmentation for plants. Trends Ecol. Evol. 1996, 11, 413–418. [Google Scholar] [CrossRef]
- Nizam, A.; Meera, S.P.; Kumar, A. Genetic and molecular mechanisms underlying mangrove adaptations to intertidal environments. iScience 2022, 25, 103547. [Google Scholar] [CrossRef]
- Yan, Y.B.; Duke, N.C.; Sun, M. Comparative Analysis of the Pattern of Population Genetic Diversity in Three Indo-West Pacific Rhizophora Mangrove Species. Front. Plant Sci. 2016, 7, 1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nualla-ong, A.; Phongdara, A.; Buapet, P. Copper and zinc differentially affect root glutathione accumulation and phytochelatin synthase gene expression of Rhizophora mucronata seedlings: Implications for mechanisms underlying trace metal tolerance. Ecotoxicol. Environ. Saf. 2020, 205, 111175. [Google Scholar] [CrossRef]
- Dahdouh-Guebas, F.; Mathenge, C.; Kairo, J.G.; Koedam, N. Utilization of Mangrove Wood Products around Mida Creek (Kenya) Amongst Subsistence and Commercial Users. Econ. Bot. 2000, 54, 513–527. [Google Scholar] [CrossRef]
- Bandaranayake, W.M. Bioactivities, bioactive compounds and chemical constituents of mangrove plants. Wetl. Ecol. Manag. 2002, 10, 421–452. [Google Scholar] [CrossRef]
- Guo, Z.; Huang, Y.; Chen, Y.; Duke, N.C.; Zhong, C.; Shi, S. Genetic discontinuities in a dominant mangrove Rhizophora apiculata (Rhizophoraceae) in the Indo-Malesian region. J. Biogeogr. 2016, 43, 1856–1868. [Google Scholar] [CrossRef]
- Triest, L.; Hasan, S.; Mitro, P.R.; De Ryck, D.J.R.; Van der Stocken, T. Geographical Distance and Large Rivers Shape Genetic Structure of Avicennia officinalis in the Highly Dynamic Sundarbans Mangrove Forest and Ganges Delta Region. Estuaries Coasts 2018, 41, 908–920. [Google Scholar] [CrossRef]
- Do, B.T.N.; Koedam, N.; Triest, L. Avicennia marina maintains genetic structure whereas Rhizophora stylosa connects mangroves in a flooded, former inner sea (Vietnam). Estuar. Coast. Shelf Sci. 2019, 222, 195–204. [Google Scholar] [CrossRef]
- Ruang-areerate, P.; Naktang, C.; Kongkachana, W.; Sangsrakru, D.; Narong, N.; Maknual, C.; Pravinvongvuthi, T.; Promchoo, W.; Yamprasai, S.; Tangphatsornruang, S.; et al. Assessment of the Genetic Diversity and Population Structure of Rhizophora apiculata Blume (Rhizophoraceae) in Thailand. Biology 2022, 11, 1449. [Google Scholar] [CrossRef] [PubMed]
- Khanbo, S.; Kongkachana, W.; Jomchai, N.; Charoensri, S.; Maknual, C.; Maprasop, P.; Phormsin, N.; Tangphatsornruang, S.; Pootakham, W. Genetic diversity and population structure of Bruguiera cylindrica along coastal areas in Thailand. Aquat. Bot. 2022, 183, 103575. [Google Scholar] [CrossRef]
- Shearman, J.R.; Naktang, C.; Sonthirod, C.; Kongkachana, W.; U-thoomporn, S.; Jomchai, N.; Maknual, C.; Yamprasai, S.; Promchoo, W.; Ruang-Areerate, P.; et al. Assembly of a hybrid mangrove, Bruguiera hainesii, and its two ancestral contributors, Bruguiera cylindrica and Bruguiera gymnorhiza. Genomics 2022, 114, 110382. [Google Scholar] [CrossRef] [PubMed]
- Pootakham, W.; Naktang, C.; Sonthirod, C.; Kongkachana, W.; Narong, N.; Sangsrakru, D.; Maknual, C.; Jiumjamrassil, D.; Chumriang, P.; Tangphatsornruang, S. Chromosome-level genome assembly of Indian mangrove (Ceriops tagal) revealed a genome-wide duplication event predating the divergence of Rhizophoraceae mangrove species. Plant Genome 2022, 15, e20217. [Google Scholar] [CrossRef]
- Pootakham, W.; Naktang, C.; Sonthirod, C.; Kongkachana, W.; Yoocha, T.; Jomchai, N.; Maknual, C.; Chumriang, P.; Pravinvongvuthi, T.; Tangphatsornruang, S. De Novo Reference Assembly of the Upriver Orange Mangrove (Bruguiera sexangula) Genome. Genome Biol. Evol. 2022, 14, evac025. [Google Scholar] [CrossRef]
- Pootakham, W.; Sonthirod, C.; Naktang, C.; Kongkachana, W.; U-thoomporn, S.; Phetchawang, P.; Maknual, C.; Jiumjamrassil, D.; Pravinvongvuthi, T.; Tangphatsornruang, S. A de novo reference assembly of the yellow mangrove Ceriops zippeliana genome. G3 Genes Genomes Genet. 2022, 12, jkac025. [Google Scholar] [CrossRef]
- Xu, S.; He, Z.; Zhang, Z.; Guo, Z.; Guo, W.; Lyu, H.; Li, J.; Yang, M.; Du, Z.; Huang, Y.; et al. The origin, diversification and adaptation of a major mangrove clade (Rhizophoreae) revealed by whole-genome sequencing. Natl. Sci. Rev. 2017, 4, 721–734. [Google Scholar] [CrossRef] [Green Version]
- Miryeganeh, M.; Marlétaz, F.; Gavriouchkina, D.; Saze, H. De novo genome assembly and in natura epigenomics reveal salinity-induced DNA methylation in the mangrove tree Bruguiera gymnorhiza. New Phytol. 2022, 233, 2094–2110. [Google Scholar] [CrossRef]
- He, Z.; Feng, X.; Chen, Q.; Li, L.; Li, S.; Han, K.; Guo, Z.; Wang, J.; Liu, M.; Shi, C.; et al. Evolution of coastal forests based on a full set of mangrove genomes. Nat. Ecol. Evol. 2022, 6, 738–749. [Google Scholar] [CrossRef]
- Maguire, T.L.; Peakall, R.; Saenger, P. Comparative analysis of genetic diversity in the mangrove species Avicennia marina (Forsk.) Vierh. (Avicenniaceae) detected by AFLPs and SSRs. Appl. Genet. 2002, 104, 388–398. [Google Scholar] [CrossRef]
- Yahya, A.F.; Hyun, J.O.; Lee, J.H.; Kim, Y.Y.; Lee, K.M.; Hong, K.N.; Kim, S.-C. Genetic variation and population genetic structure of Rhizophora apiculata (Rhizophoraceae) in the greater Sunda Islands, Indonesia using microsatellite markers. J. Plant Res. 2014, 127, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Castro, E.; Muñiz-Salazar, R.; Enríquez-Paredes, L.M.; Riosmena-Rodríguez, R.; Dodd, R.S.; Tovilla-Hernández, C.; Arredondo-García, M.C. Genetic population structure of red mangrove (Rhizophora mangle L.) along the northwestern coast of Mexico. Aquat. Bot. 2012, 99, 20–26. [Google Scholar] [CrossRef]
- Dasgupta, N.; Nandy, P.; Sengupta, C.; Das, S. RAPD and ISSR marker mediated genetic polymorphism of two mangroves Bruguiera gymnorrhiza and Heritiera fomes from Indian Sundarbans in relation to their sustainability. Physiol. Mol. Biol. Plants 2015, 21, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Banks, T.W.; Cloutier, S. SNP Discovery through Next-Generation Sequencing and Its Applications. Int. J. Plant Genom. 2012, 2012, 831460. [Google Scholar] [CrossRef]
- Van Tassell, C.P.; Smith, T.P.; Matukumalli, L.K.; Taylor, J.F.; Schnabel, R.D.; Lawley, C.T.; Haudenschild, C.D.; Moore, S.S.; Warren, W.C.; Sonstegard, T.S. SNP discovery and allele frequency estimation by deep sequencing of reduced representation libraries. Nat. Methods 2008, 5, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhao, S.; Li, M.; Zhang, C.; Qu, H.; Li, Q.; Li, J.; Lin, Y.; Pu, Z. Genome-wide genetic diversity detection and population structure analysis in sweetpotato (Ipomoea batatas) using RAD-seq. Genomics 2020, 112, 1978–1987. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Yan, S.-Y.; Hui, W.-K.; Gong, W. SNP discovery for genetic diversity and population structure analysis coupled with restriction-associated DNA (RAD) sequencing in walnut cultivars of Sichuan Province, China. Biotechnol. Biotechnol. Equip. 2020, 34, 652–664. [Google Scholar] [CrossRef]
- Zhou, W.; Xiang, Q.-Y. Phylogenomics AND biogeography of Castanea (chestnut) and Hamamelis (witch-hazel)—Choosing between RAD-seq and Hyb-Seq approaches. Mol. Phylogenet. Evol. 2022, 176, 107592. [Google Scholar] [CrossRef]
- Pootakham, W.; Sonthirod, C.; Naktang, C.; Kongkachana, W.; Sangsrakru, D.; U-thoomporn, S.; Maknual, C.; Meepol, W.; Promchoo, W.; Maprasop, P.; et al. A chromosome-scale reference genome assembly of yellow mangrove (Bruguiera parviflora) reveals a whole genome duplication event associated with the Rhizophoraceae lineage. Mol. Ecol. Resour. 2022, 22, 1939–1953. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Salzberg, S.L.; Zhu, W.; Pertea, M.; Allen, J.E.; Orvis, J.; White, O.; Buell, C.R.; Wortman, J.R. Automated eukaryotic gene structure annotation using EVidenceModeler and the Program to Assemble Spliced Alignments. Genome Biol. 2008, 9, R7. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Adams, M.D.; Zhou, H.; Kerlavage, A.R. A tool for analyzing and annotating genomic sequences. Genomics 1997, 46, 37–45. [Google Scholar] [CrossRef]
- Stanke, M.; Morgenstern, B. AUGUSTUS: A web server for gene prediction in eukaryotes that allows user-defined constraints. Nucleic Acids Res. 2005, 33, W465–W467. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO Update: Novel and Streamlined Workflows along with Broader and Deeper Phylogenetic Coverage for Scoring of Eukaryotic, Prokaryotic, and Viral Genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2007, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer Science+Business Media, LLC: New York, NY, USA, 2016; p. xvi. 260p. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.-J.; Sun, M. Population genetic structure of Ceriops tagal (Rhizophoraceae) in Thailand and China. Wetl. Ecol. Manag. 2001, 9, 213–219. [Google Scholar] [CrossRef]
- Liao, P.-C.; Havanond, S.; Huang, S. Phylogeography of Ceriops tagal (Rhizophoraceae) in Southeast Asia: The land barrier of the Malay Peninsula has caused population differentiation between the Indian Ocean and South China Sea. Conserv. Genet. 2007, 8, 89–98. [Google Scholar] [CrossRef]
- Minobe, S.; Fukui, S.; Saiki, R.; Kajita, T.; Changtragoon, S.; Ab Shukor, N.A.; Latiff, A.; Ramesh, B.R.; Koizumi, O.; Yamazaki, T. Highly differentiated population structure of a Mangrove species, Bruguiera gymnorhiza (Rhizophoraceae) revealed by one nuclear GapCp and one chloroplast intergenic spacer trnF–trnL. Conserv. Genet. 2010, 11, 301–310. [Google Scholar] [CrossRef]
- Wee, A.K.; Noreen, A.M.; Ono, J.; Takayama, K.; Kumar, P.P.; Tan, H.T.; Saleh, M.N.; Kajita, T.; Webb, E.L. Genetic structures across a biogeographical barrier reflect dispersal potential of four Southeast Asian mangrove plant species. J. Biogeogr. 2020, 47, 1258–1271. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, R.; Tang, T.; Huang, Y.; Zhong, Y.; Shi, S. Genetic variation in central and peripheral populations of Excoecaria agallocha from Indo-West Pacific. Aquat. Bot. 2008, 89, 57–62. [Google Scholar] [CrossRef]
- Yang, Y.; Li, J.; Yang, S.; Li, X.; Fang, L.; Zhong, C.; Duke, N.C.; Zhou, R.; Shi, S. Effects of Pleistocene sea-level fluctuations on mangrove population dynamics: A lesson from Sonneratia alba. BMC Evol. Biol. 2017, 17, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triest, L.; Satyanarayana, B.; Delange, O.; Sarker, K.K.; Sierens, T.; Dahdouh-Guebas, F. Barrier to Gene Flow of Grey Mangrove Avicennia marina Populations in the Malay Peninsula as Revealed From Nuclear Microsatellites and Chloroplast Haplotypes. Front. Conserv. Sci. 2021, 2, 727819. [Google Scholar] [CrossRef]
- Wee, A.K.S.; Takayama, K.; Asakawa, T.; Thompson, B.; Onrizal; Sungkaew, S.; Tung, N.X.; Nazre, M.; Soe, K.K.; Tan, H.T.W.; et al. Oceanic currents, not land masses, maintain the genetic structure of the mangrove Rhizophora mucronata Lam. (Rhizophoraceae) in Southeast Asia. J. Biogeogr. 2014, 41, 954–964. [Google Scholar] [CrossRef]
- Ng, W.L.; Onishi, Y.; Inomata, N.; Teshima, K.M.; Chan, H.T.; Baba, S.; Changtragoon, S.; Siregar, I.Z.; Szmidt, A.E. Closely related and sympatric but not all the same: Genetic variation of Indo-West Pacific Rhizophora mangroves across the Malay Peninsula. Conserv. Genet. 2015, 16, 137–150. [Google Scholar] [CrossRef]
- Wright, S. The interpretation of population structure by F-statistics with special regard to systems of mating. Evolution 1965, 19, 395–420. [Google Scholar] [CrossRef]
- Shi, J.; Joshi, J.; Tielbörger, K.; Verhoeven, K.J.F.; Macel, M. Costs and benefits of admixture between foreign genotypes and local populations in the field. Ecol. Evol. 2018, 8, 3675–3684. [Google Scholar] [CrossRef] [PubMed]
- Van Hengstum, T.; Lachmuth, S.; Oostermeijer, J.G.B.; den Nijs, H.C.M.; Meirmans, P.G.; van Tienderen, P.H. Human-induced hybridization among congeneric endemic plants on Tenerife, Canary Islands. Plant Syst. Evol. 2012, 298, 1119–1131. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.H.D. Human Impact on Plant Gene Pools and Sampling for Their Conservation. Oikos 1992, 63, 109–118. [Google Scholar] [CrossRef]
- Duke, N.; Ball, M.; Ellison, J. Factors influencing biodiversity and distributional gradients in mangroves. Glob. Ecol. Biogeogr. Lett. 1998, 7, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Pumijumnong, N. Mangrove forests in Thailand. In Mangrove Ecosystems of Asia: Status, Challenges and Management Strategies; Faridah-Hanum, I., Latiff, A., Hakeem, K.R., Ozturk, M., Eds.; Springer: New York, NY, USA, 2014; pp. 61–79. [Google Scholar]
- Drexler, J.Z. Maximum longevities of Rhizophora apiculata and R. mucronata propagules. Pac. Sci. 2001, 55, 17–22. [Google Scholar] [CrossRef]
- Pil, M.W.; Boeger, M.R.; Muschner, V.C.; Pie, M.R.; Ostrensky, A.; Boeger, W.A. Postglacial north-south expansion of populations of Rhizophora mangle (Rhizophoraceae) along the Brazilian coast revealed by microsatellite analysis. Am. J. Bot. 2011, 98, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Millán-Aguilar, O.; Manzano-Sarabia, M.; Nettel-Hernanz, A.; Dodd, R.S.; Hurtado-Oliva, M.Á.; Velázquez-Velázquez, E. Genetic diversity of the black mangrove Avicennia germinans (L.) Stearn in Northwestern Mexico. Forests 2016, 7, 197. [Google Scholar] [CrossRef] [Green Version]
- Wee, A.K.S.; Teo, J.X.H.; Chua, J.L.; Takayama, K.; Asakawa, T.; Meenakshisundaram, S.H.; Onrizal; Adjie, B.; Ardli, E.R.; Sungkaew, S.; et al. Vicariance and Oceanic Barriers Drive Contemporary Genetic Structure of Widespread Mangrove Species Sonneratia alba J. Sm in the Indo-West Pacific. Forests 2017, 8, 483. [Google Scholar] [CrossRef] [Green Version]
- Arnaud-Haond, S.; Teixeira, S.; Massa, S.I.; Billot, C.; Saenger, P.; Coupland, G.; Duarte, C.M.; Serrão, E.A. Genetic structure at range edge: Low diversity and high inbreeding in Southeast Asian mangrove (Avicennia marina) populations. Mol. Ecol. 2006, 15, 3515–3525. [Google Scholar] [CrossRef] [PubMed]
- Canty, S.W.J.; Kennedy, J.P.; Fox, G.; Matterson, K.; González, V.L.; Núñez-Vallecillo, M.L.; Preziosi, R.F.; Rowntree, J.K. Mangrove diversity is more than fringe deep. Sci. Rep. 2022, 12, 1695. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Li, X.; Yang, M.; Wang, X.; Zhong, C.; Duke, N.C.; Wu, C.-I.; Shi, S. Speciation with gene flow via cycles of isolation and migration: Insights from multiple mangrove taxa. Natl. Sci. Rev. 2018, 6, 275–288. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Maguire, T.L.; Saenger, P.; Baverstock, P.; Henry, R. Microsatellite analysis of genetic structure in the mangrove species Avicennia marina (Forsk.) Vierh. (Avicenniaceae). Mol. Ecol. 2000, 9, 1853–1862. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.B.; Ding, W.Y.; Qiu, J.B.; Wang, G.; Zhou, Z.M.; Chen, J.F.; Ai, W.M.; Wang, C.Y.; Xie, Q.L. The genetic diversity of the mangrove Kandelia obovata in China revealed by issr analysis. Pak. J. Bot. 2010, 42, 3755–3764. [Google Scholar]
- Azman, A.; Ng, K.-K.-S.; Ng, C.-H.; Lee, C.-T.; Tnah, L.-H.; Zakaria, N.-F.; Mahruji, S.; Perdan, K.; Abdul-Kadir, M.-Z.; Cheng, A. Low genetic diversity indicating the threatened status of Rhizophora apiculata (Rhizophoraceae) in Malaysia: Declined evolution meets habitat destruction. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Islam, M.S.; Lian, C.; Kameyama, N.; Hogetsu, T. Low genetic diversity and limited gene flow in a dominant mangrove tree species (Rhizophora stylosa) at its northern biogeographical limit across the chain of three Sakishima islands of the Japanese archipelago as revealed by chloroplast and nuclear SSR analysis. Plant Syst. Evol. 2014, 300, 1123–1136. [Google Scholar] [CrossRef]
- Senjaya, S.K. Genetic diversity of Rhizophora apiculata blume in Banggai Kepulauan inferred from sequence-related amplified polymorphism (SRAP) marker. In Proceedings of the International Conference on Science and Engineering (ICSE-UIN-SUKA 2021), Yogyakarta, Indonesia, 27 October 2021; pp. 154–158. [Google Scholar]
- Lu, W.; Zou, Z.; Hu, X.; Yang, S. Genetic Diversity and Mating System of Two Mangrove Species (Rhizophora apiculata and Avicennia marina) in a Heavily Disturbed Area of China. Diversity 2022, 14, 115. [Google Scholar] [CrossRef]
- Ge, J.P.; Cai, B.; Ping, W.; Song, G.; Ling, H.; Lin, P. Mating system and population genetic structure of Bruguiera gymnorrhiza (Rhizophoraceae), a viviparous mangrove species in China. J. Exp. Mar. Biol. Ecol. 2005, 326, 48–55. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W. Effects of Life History Traits on Genetic Diversity in Plant Species. Philos. Trans. Biol. Sci. 1996, 351, 1291–1298. [Google Scholar]
- Mori, G.M.; Zucchi, M.I.; Souza, A.P. Multiple-geographic-scale genetic structure of two mangrove tree species: The roles of mating system, hybridization, limited dispersal and extrinsic factors. PLoS ONE 2015, 10, e0118710. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 10× Genomics | |
|---|---|
| N50 scaffold size (bases) | 542,540 |
| L50 scaffold number | 108 |
| N75 scaffold size (bases) | 157,341 |
| L75 scaffold number | 287 |
| N90 scaffold size (bases) | 6738 |
| L90 scaffold number | 1569 |
| Total (bases) | 219,246,407 |
| Number of scaffolds | 12,956 |
| Number of scaffolds ≥ 100 kb | 340 |
| Number of scaffolds ≥ 1 Mb | 37 |
| Number of scaffolds ≥ 10 Mb | - |
| Longest scaffold (bases) | 4,666,015 |
| GC content (%) | 35 |
| BUSCO evaluation (% completeness) | 96.4 |
| R. mucronata | |
|---|---|
| Number of predicted gene models | 28,500 |
| Total gene length (Mb) | 69.73 |
| Average gene size (nt) | 2447 |
| Average number of exons/gene | 4.64 |
| Total exon length (Mb) | 31.21 |
| Average exon length (nt) | 235.60 |
| GC content of exons (%) | 45.08 |
| Average number of introns/gene | 3.64 |
| Total intron length (Mb) | 38.55 |
| Average intron length (nt) | 370.80 |
| GC content of introns (%) | 34.55 |
| Types of Repeats | Bases (Mb) | % of the Assembly | % of Total Repeats |
|---|---|---|---|
| DNA transposons | 3.61 | 1.65 | 5.91 |
| Retrotransposons: | |||
| LINE | 1.93 | 0.88 | 3.16 |
| SINE | 0.20 | 0.09 | 0.32 |
| LTR: Copia | 17.50 | 7.98 | 28.65 |
| LTR: Gypsy | 11.94 | 5.44 | 19.55 |
| LTR: Others | 4.65 | 2.12 | 7.61 |
| Simple sequence repeats | 4.46 | 2.03 | 7.30 |
| Others | 16.77 | 7.66 | 27.5 |
| Total | 61.06 | 27.85 |
| Source of Variation | df | Sum of Squares | Variance Components | Percentage of Variation | F-Statistic |
|---|---|---|---|---|---|
| Among populations | 1 | 2425.68 | 53.92 | 9.10 | FST = 0.09 ** |
| Within populations | 158 | 85,072.54 | 538.43 | 90.90 | |
| Total | 159 | 87,498.22 | 592.35 |
| Population | N | Ne | I | Ho | He | PPL | FIS |
|---|---|---|---|---|---|---|---|
| Cluster 1 | 70 | 1.701 | 0.578 | 0.604 | 0.394 | 100% | −0.451 |
| Cluster 2 | 10 | 1.702 | 0.567 | 0.633 | 0.389 | 98.19% | −0.538 |
| Overall | 80 | 1.701 | 0.573 | 0.619 | 0.391 | 99.09% | −0.494 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naktang, C.; Khanbo, S.; Yundaeng, C.; U-thoomporn, S.; Kongkachana, W.; Jiumjamrassil, D.; Maknual, C.; Wanthongchai, P.; Tangphatsornruang, S.; Pootakham, W. Assessment of the Genetic Diversity and Population Structure of Rhizophora mucronata along Coastal Areas in Thailand. Biology 2023, 12, 484. https://doi.org/10.3390/biology12030484
Naktang C, Khanbo S, Yundaeng C, U-thoomporn S, Kongkachana W, Jiumjamrassil D, Maknual C, Wanthongchai P, Tangphatsornruang S, Pootakham W. Assessment of the Genetic Diversity and Population Structure of Rhizophora mucronata along Coastal Areas in Thailand. Biology. 2023; 12(3):484. https://doi.org/10.3390/biology12030484
Chicago/Turabian StyleNaktang, Chaiwat, Supaporn Khanbo, Chutintorn Yundaeng, Sonicha U-thoomporn, Wasitthee Kongkachana, Darunee Jiumjamrassil, Chatree Maknual, Poonsri Wanthongchai, Sithichoke Tangphatsornruang, and Wirulda Pootakham. 2023. "Assessment of the Genetic Diversity and Population Structure of Rhizophora mucronata along Coastal Areas in Thailand" Biology 12, no. 3: 484. https://doi.org/10.3390/biology12030484