
Stimulation of Nuclear Factor (Erythroid-Derived 2)-like 2 Signaling by Nucleus Targeted Irradiation with Proton Microbeam

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Antibodies and Chemicals
2.3. Nucleus Targeted Irradiation with Proton Microbeam
2.4. Immunofluorescence Staining
2.5. Measurement of Intracellular ROS
2.6. Statistical Analysis
3. Results
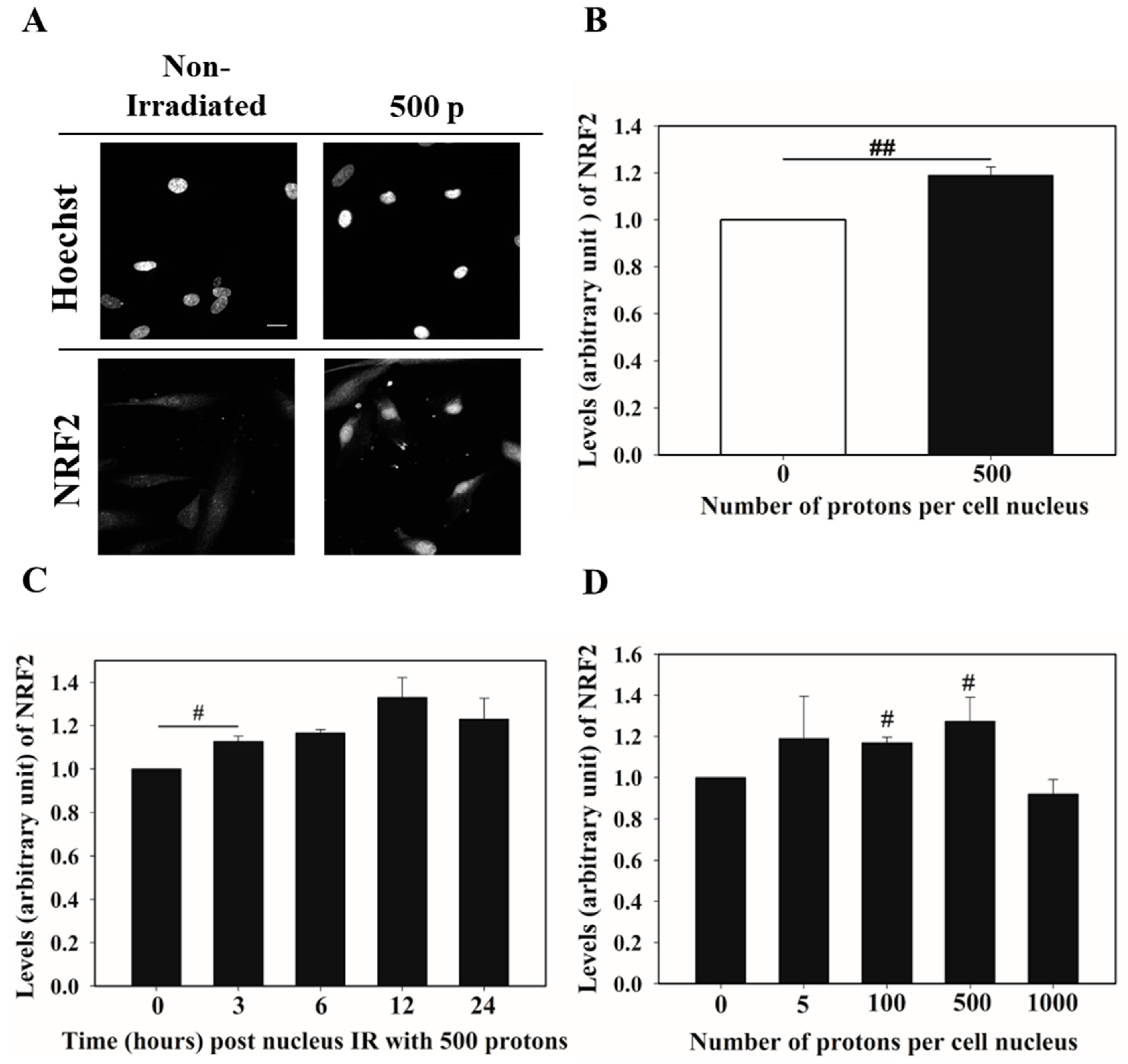
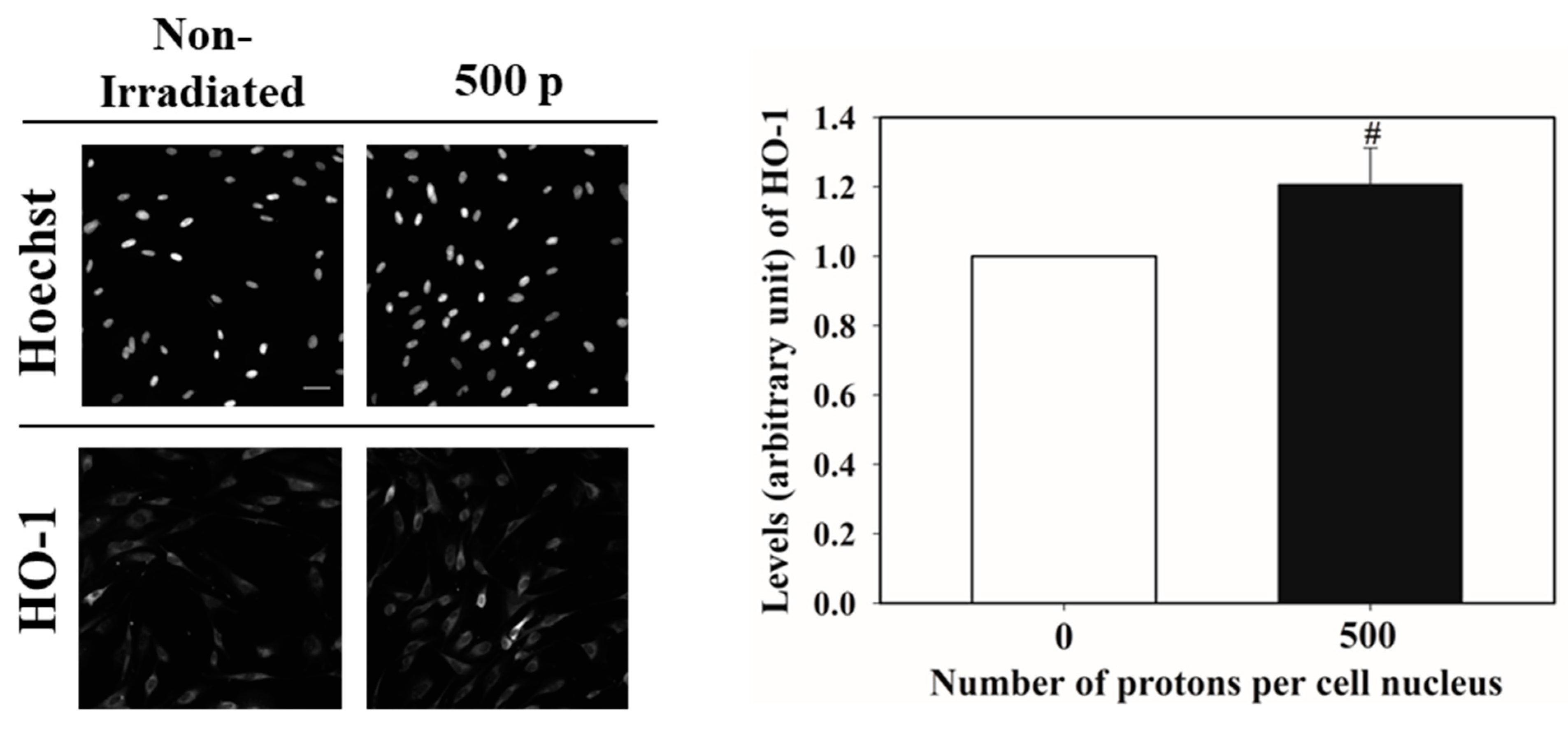
3.1. Nucleus Targeted Irradiation Promotes the Nucleus Accumulation of NRF2
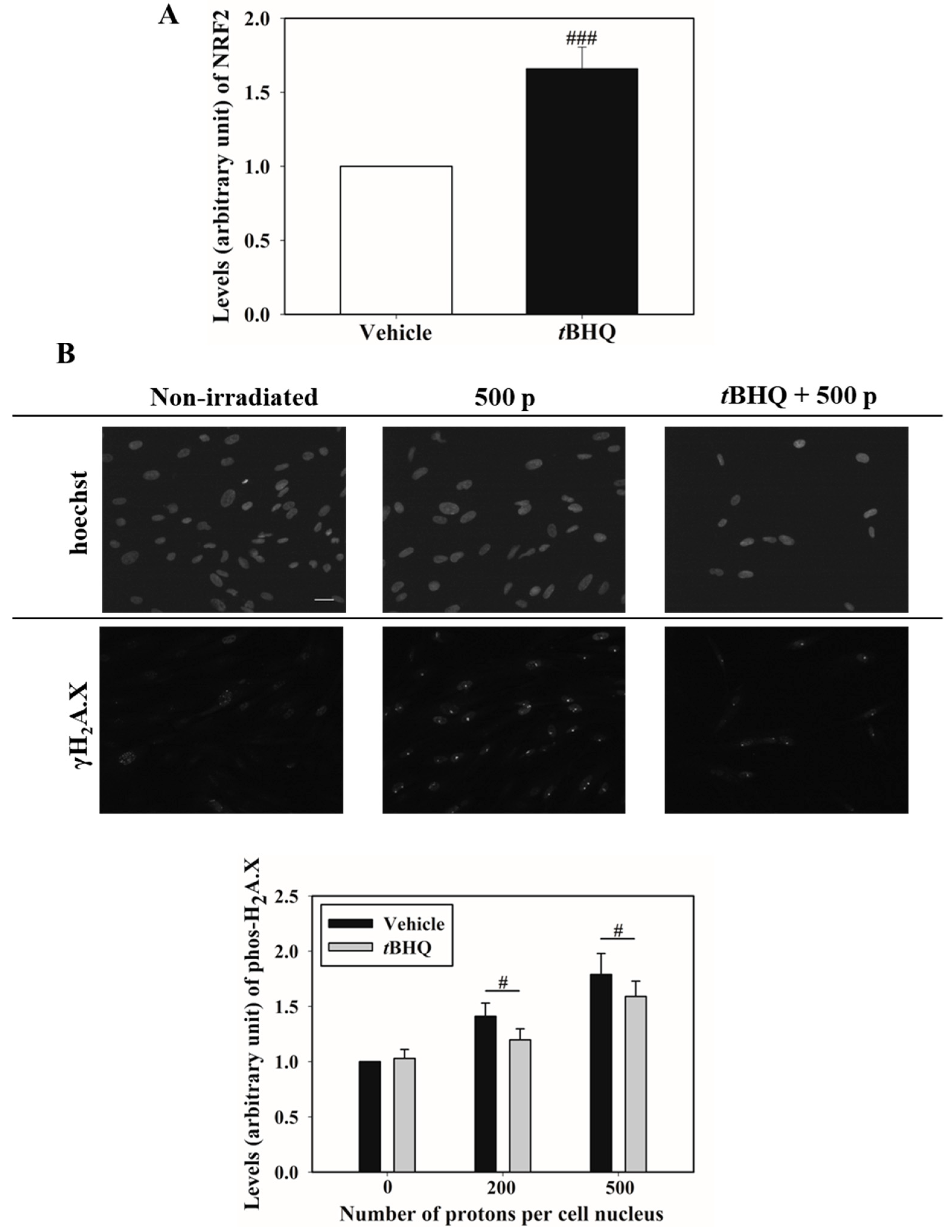
3.2. Activation of NRF2 Alleviates Nucleus Irradiation Induced DNA Double-Strand Breaks
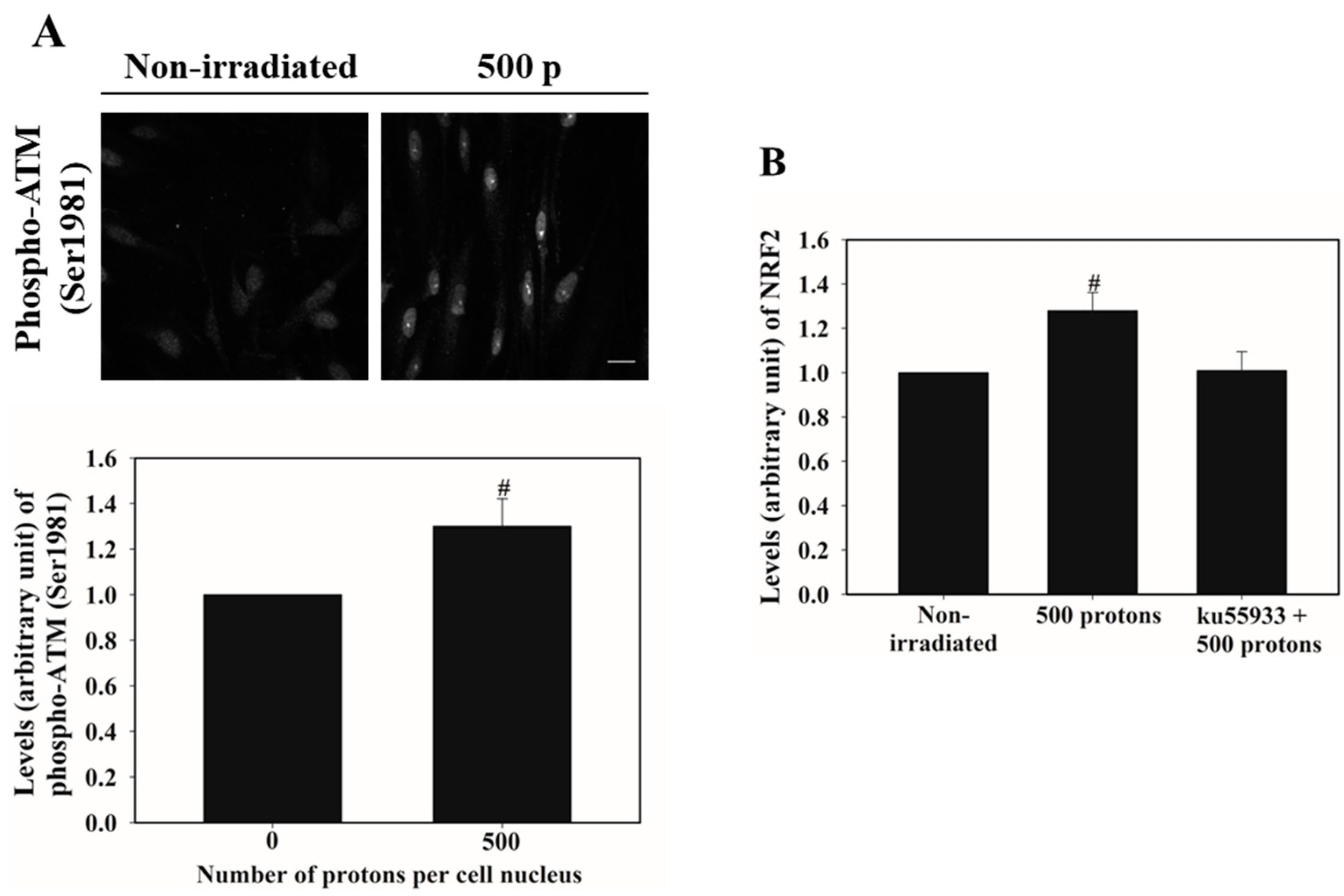
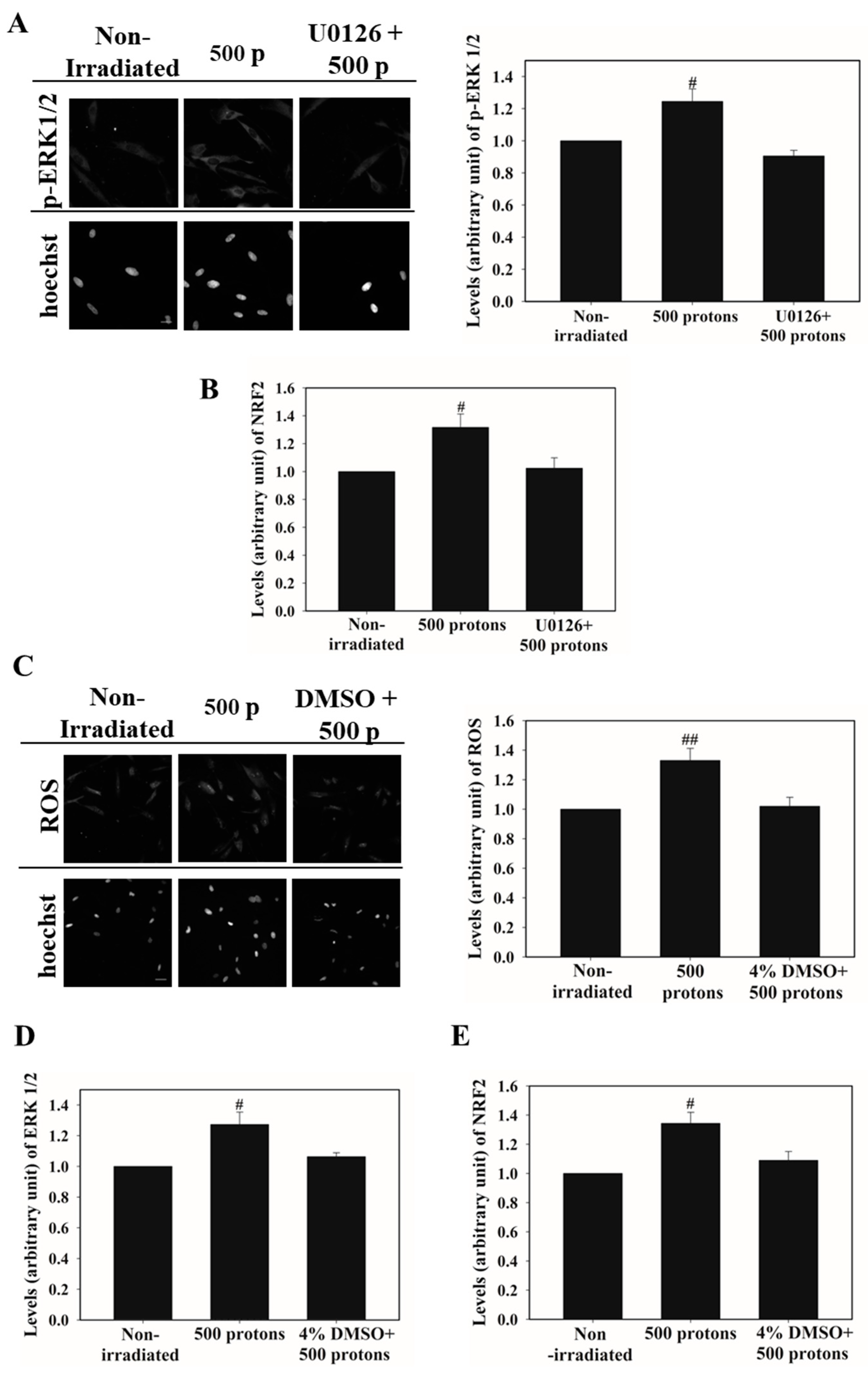
3.3. ATM Activation Is Involved in Stimulating NRF2 Nucleus Accumulation
3.4. ERK 1/2 Activation Promotes NRF2 Nucleus Accumulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prise, K.M.; Belyakov, O.V.; Folkard, M.; Michael, B.D. Studies of bystander effects in human fibroblasts using a charged particle microbeam. Int. J. Radiat. Biol. 1998, 74, 793–798. [Google Scholar] [CrossRef]
- Miller, R.C.; Randers-Pehrson, G.; Geard, C.R.; Hall, E.J.; Brenner, D.J. The oncogenic transforming potential of the passage of single alpha particles through mammalian cell nuclei. Proc. Natl. Acad. Sci. USA 1999, 96, 19–22. [Google Scholar] [CrossRef] [Green Version]
- Autsavapromporn, N.; Kobayashi, A.; Liu, C.; Jaikang, C.; Tengku Ahmad, T.A.; Oikawa, M.; Konishi, T. Hypoxia and proton microbeam: Role of gap junction intercellular communication in inducing bystander responses on human lung cancer cells and normal cells. Radiat. Res. 2022, 197, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Autsavapromporn, N.; Ahmad, T.A.F.T.; Oikawa, M.; Homma-Takeda, S.; Furusawa, Y.; Wang, J.; Konishi, T. Bystander WI-38 cells modulate DNA double-STRAND break repair in microbeam-targeted A549 cells through gap junction intercellular communication. Radiat. Prot. Dosim. 2019, 183, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Kobayashi, A.; Ohsawa, D.; Oikawa, M.; Konishi, T. Cytoplasmic radiation induced radio-adaptive response in human lung fibroblast WI-38 cells. Radiat. Res. 2020, 194, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Hei, T.K.; Wu, L.J.; Liu, S.X.; Vannais, D.; Waldren, C.A.; Randers-Pehrson, G. Mutagenic effects of a single and an exact number of alpha particles in mammalian cells. Proc. Natl. Acad. Sci. USA 1997, 94, 3765–3770. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.J.; Randers-Pehrson, G.; Xu, A.; Waldren, C.A.; Geard, C.R.; Yu, Z.; Hei, T.K. Targeted cytoplasmic irradiation with alpha particles induces mutations in mammalian cells. Proc. Natl. Acad. Sci. USA 1999, 96, 4959–4964. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.; Folkard, M.; Michael, B.D.; Prise, K.M. Targeted cytoplasmic irradiation induces bystander responses. Proc. Natl. Acad. Sci. USA 2004, 101, 13495–13500. [Google Scholar] [CrossRef] [Green Version]
- Konishi, T.; Oikawa, M.; Suya, N.; Ishikawa, T.; Maeda, T.; Kobayashi, A.; Shiomi, N.; Kodama, K.; Hamano, T.; Homma-Takeda, S.; et al. SPICE-NIRS microbeam: A focused vertical system for proton irradiation of a single cell for radiobiological research. J. Radiat. Res. 2013, 54, 736–747. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, A.; Ahmad, T.T.A.F.; Autsavapromporn, N.; Oikawa, M.; Homma-Takeda, S.; Furusawa, Y.; Wang, J.; Konishi, T. Enhanced DNA double-strand break repair of microbeam targeted A549 lung carcinoma cells by adjacent WI38 normal lung fibroblast cells via bi-directional signaling. Mutat. Res. 2017, 803–805, 1–8. [Google Scholar] [CrossRef]
- Konishi, T.; Kobayashi, A.; Ahmad, T.T.A.F.; Wang, J. Enhanced DNA double strand break repair triggered by microbeam irradiation induced cytoplasmic damage. J. Radiat. Cancer Res. 2018, 9, 183–189. [Google Scholar] [CrossRef]
- Funayama, T. Heavy-ion microbeams for biological science: Development of system and utilization for biological experiments in QST-Takasaki. Quantum Beam Sci. 2019, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Hamada, N.; Aoki-Nakano, M.; Funayama, T.; Sakashita, T.; Wada, S.; Kakizaki, T.; Kobayashi, Y.; Furusawa, Y. Dependence of the bystander effect for micronucleus formation on dose of heavy-ion radiation in normal human fibroblasts. Radiat. Prot. Dosim. 2015, 166, 152–156. [Google Scholar] [CrossRef]
- Autsavapromporn, N.; Suzuki, M.; Funayama, T.; Usami, N.; Plante, I.; Yokota, Y.; Mutou, Y.; Ikeda, H.; Kobayashi, K.; Kobayashi, Y.; et al. Gap junction communication and the propagation of bystander effects induced by microbeam irradiation in human fibroblast cultures: The impact of radiation quality. Radiat. Res. 2013, 180, 367–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jella, K.K.; Moriarty, R.; McClean, B.; Byrne, H.J.; Lyng, F.M. Reactive oxygen species and nitric oxide signaling in bystander cells. PLoS ONE 2018, 13, e0195371. [Google Scholar] [CrossRef] [PubMed]
- Sporn, M.; Liby, K. NRF2 and cancer: The good, the bad and the importance of context. Nat. Rev. Cancer 2012, 12, 564–571. [Google Scholar] [CrossRef] [Green Version]
- You, G.; Chang, J.; Li, Y.; Huang, C.; Tsai, Y.; Fan, K.; Kang, C.; Huang, S.; Chang, P.; Cheng, A. MYH9 facilitates cell invasion and radioresistance in head and neck cancer via modulation of cellular ROS levels by activating the MAPK-Nrf2-GCLC pathway. Cells 2022, 11, 2855. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Yoshida, R.; Kawahara, K.; Sakata, J.; Arita, H.; Nakashima, H.; Takahashi, N.; Hirayama, M.; Nagata, M.; Hirosue, A.; et al. The antioxidative stress regulator Nrf2 potentiates radioresistance of oral squamous cell carcinoma accompanied with metabolic modulation. Lab. Investig. 2022, 102, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Pandita, R.; Eskiocak, U.; Ly, P.; Kaisani, A.; Kumar, R.; Cornelius, C.; Wright, W.; Pandita, T.; Shay, J. Targeting of Nrf2 induces DNA damage signaling and protects colonic epithelial cells from ionizing radiation. Proc. Natl. Acad. Sci. USA 2012, 109, E2949–E2955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Konishi, T. Nuclear factor (erythroid-derived 2)-like 2 antioxidative response mitigates cytoplasmic radiation-induced DNA double-strand breaks. Cancer Sci. 2019, 110, 686–696. [Google Scholar] [CrossRef] [Green Version]
- Dinkova-Kostova, A.T.; Copple, I.M. Advances and challenges in therapeutic targeting of NRF2. Trends Pharm. Sci. 2023, 44, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Pan, C.; Zhang, X.; Yang, T.; Hu, T.; Zheng, L.; Cao, S.; Feng, C.; Hu, X.; Chai, X.; et al. Nuclear factor Nrf2 promotes glycosidase OGG1 expression by activating the AKT pathway to enhance leukemia cell resistance to cytarabine. J. Biol. Chem. 2022, 299, 102798. [Google Scholar] [CrossRef]
- Kobayashi, E.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayakumar, S.; Pal, D.; Sandur, S. Nrf2 facilitates repair of radiation induced DNA damage through homologous recombination repair pathway in a ROS independent manner in cancer cells. Mutat. Res. 2015, 779, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Davidson, M.M.; Zhou, H.; Wang, C.; Walker, W.F.; Hei, T.K. Cytoplasmic irradiation results in mitochondrial dysfunction and DRP1-dependent mitochondrial fission. Cancer Res. 2013, 73, 6700–6710. [Google Scholar] [CrossRef] [Green Version]
- Owens, D.; Keyse, S. Differential regulation of MAP kinase signalling by dual-specificity protein phosphatases. Oncogene 2007, 26, 3203–3213. [Google Scholar] [CrossRef] [Green Version]
- Williams, T.M.; Flecha, A.R.; Keller, P.; Ram, A.; Karnak, D.; Galbán, S.; Galbán, C.J.; Ross, B.D.; Lawrence, T.S.; Rehemtulla, A.; et al. Cotargeting MAPK and PI3K signaling with concurrent radiotherapy as a strategy for the treatment of pancreatic cancer. Mol. Cancer Ther. 2012, 11, 1193–1202. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Ivanov, V.N.; Gillespie, J.; Geard, C.R.; Amundson, S.A.; Brenner, D.J.; Yu, Z.; Lieberman, H.B.; Hei, T.K. Mechanism of radiation-induced bystander effect: Role of the cyclooxygenase-2 signaling pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 14641–14646. [Google Scholar] [CrossRef] [Green Version]
- Hong, M.; Xu, A.; Zhou, H.; Wu, L.; Randers-Pehrson, G.; Santella, R.M.; Yu, Z.; Hei, T.K. Mechanism of genotoxicity induced by targeted cytoplasmic irradiation. Br. J. Cancer 2010, 103, 1263–1268. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, B.; Wuu, Y.R.; Davidson, M.M.; Hei, T.K. Targeted cytoplasmic irradiation and autophagy. Mutat. Res. 2017, 806, 88–97. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Q.; Wuu, Y.R.; Zou, S.; Hei, T.K. Cytoplasmic irradiation induces metabolic shift in human small airway epithelial cells via activation of pim-1 kinase. Radiat. Res. 2017, 187, 441–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Oikawa, M.; Konishi, T. Stimulation of Nuclear Factor (Erythroid-Derived 2)-like 2 Signaling by Nucleus Targeted Irradiation with Proton Microbeam. Biology 2023, 12, 419. https://doi.org/10.3390/biology12030419
Wang J, Oikawa M, Konishi T. Stimulation of Nuclear Factor (Erythroid-Derived 2)-like 2 Signaling by Nucleus Targeted Irradiation with Proton Microbeam. Biology. 2023; 12(3):419. https://doi.org/10.3390/biology12030419
Chicago/Turabian StyleWang, Jun, Masakazu Oikawa, and Teruaki Konishi. 2023. "Stimulation of Nuclear Factor (Erythroid-Derived 2)-like 2 Signaling by Nucleus Targeted Irradiation with Proton Microbeam" Biology 12, no. 3: 419. https://doi.org/10.3390/biology12030419