
Investigating the Timing and Extent of Juvenile and Fetal Bone Diagenesis in a Temperate Environment

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Depositional Environment
2.2. Histological Methodology
2.3. Data Analysis
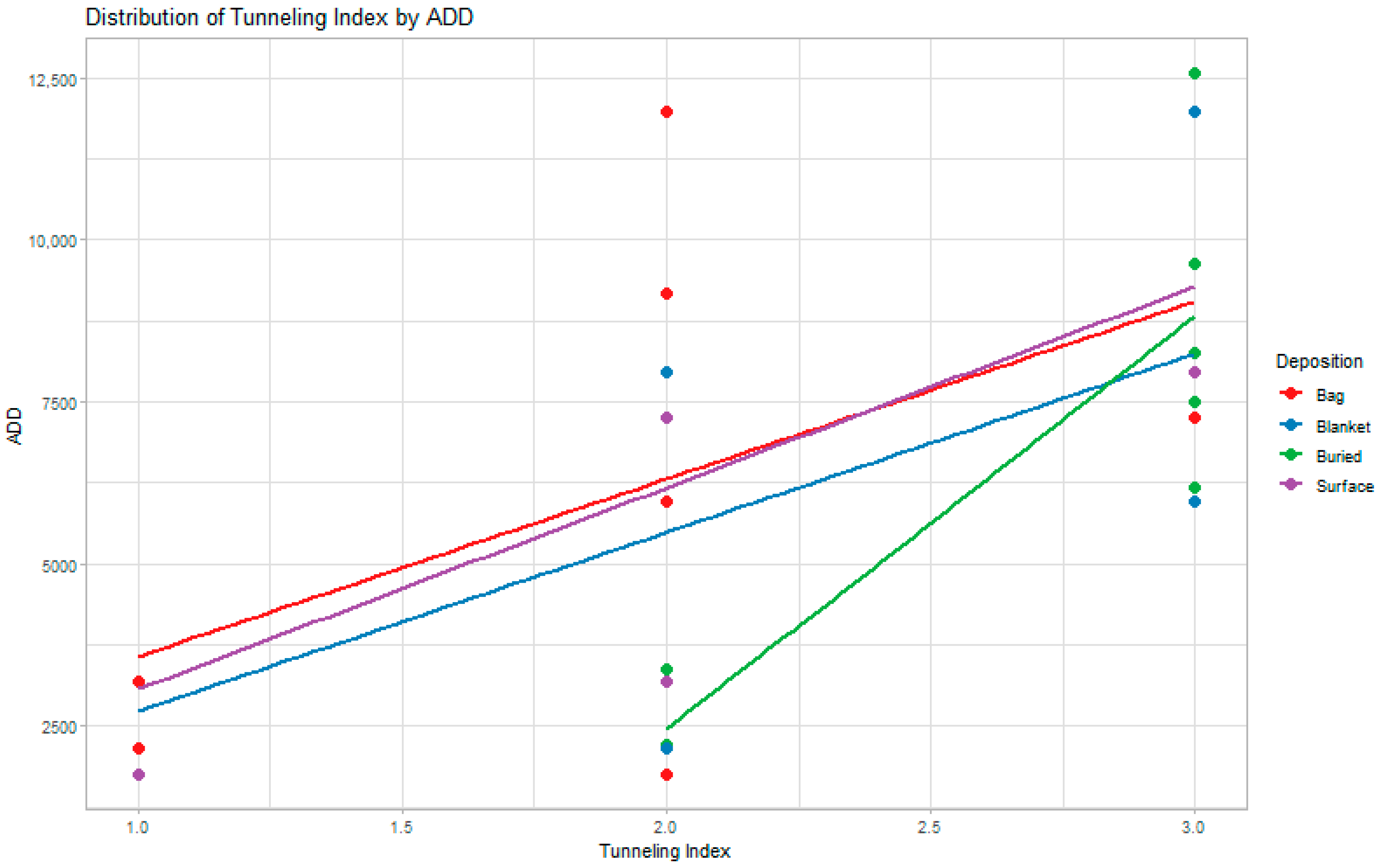
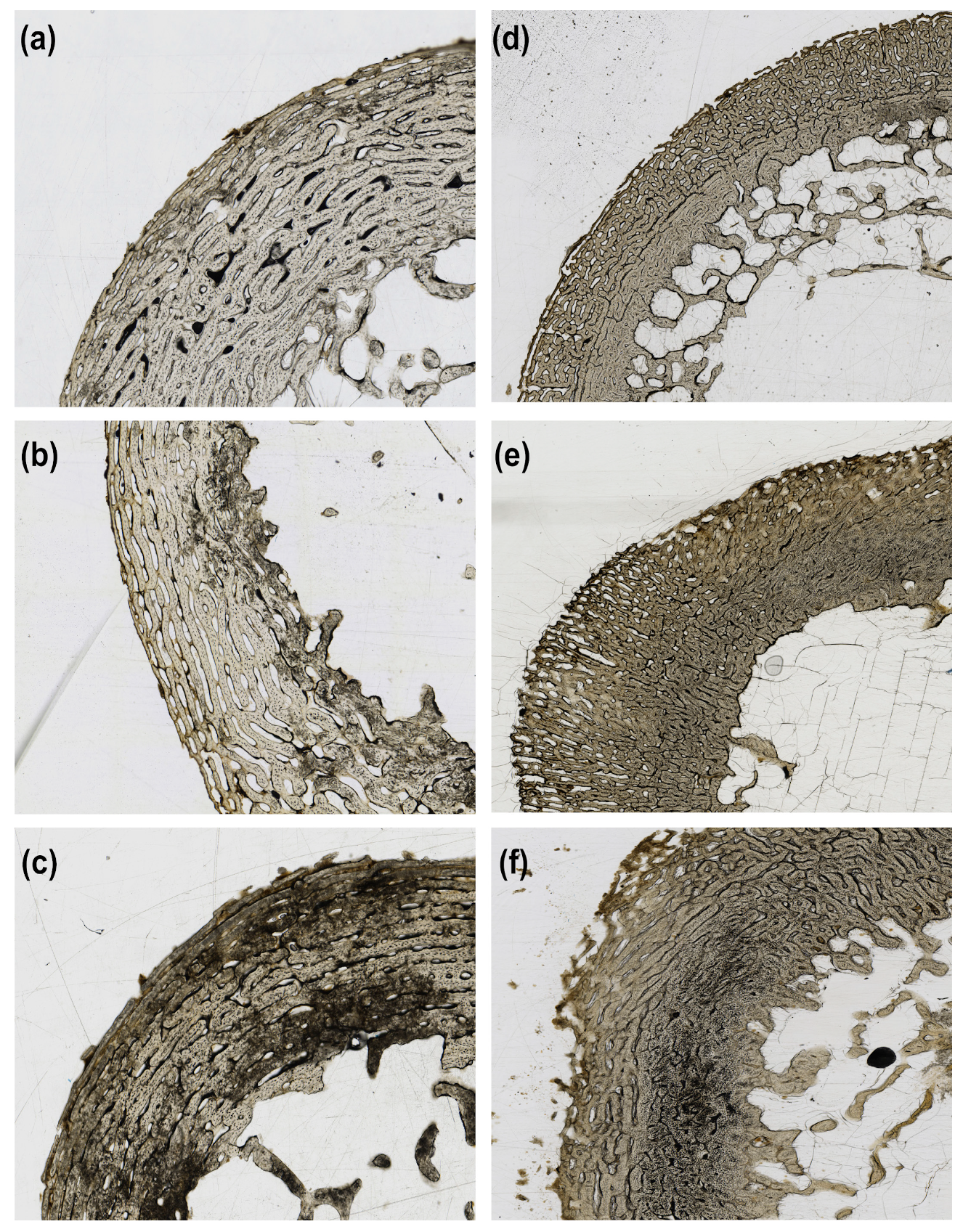
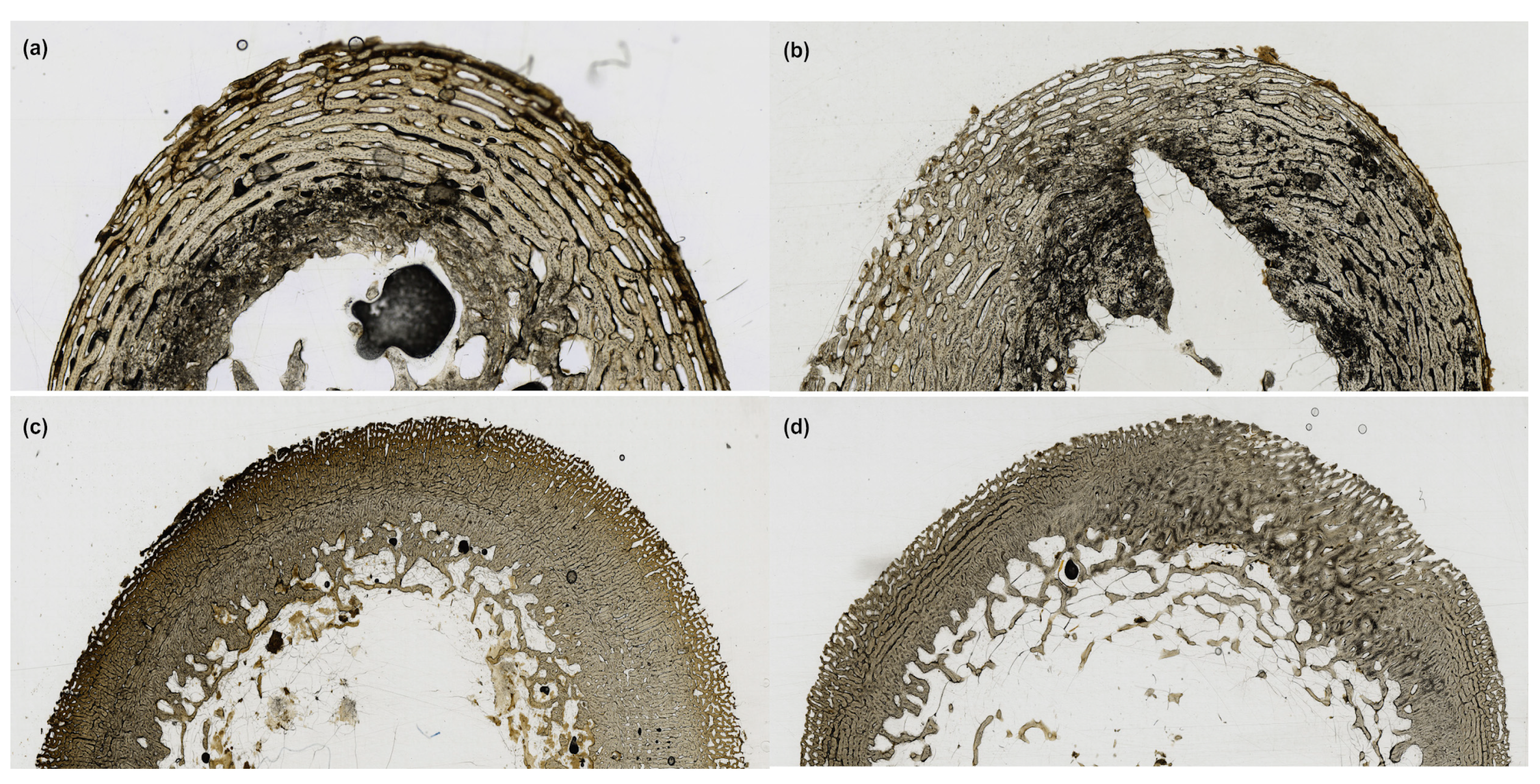
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Emmons, A.L.; Deel, H.; Davis, M.; Metcalf, J.L. Soft tissue decomposition in terrestrial ecosystems. In Manual of Forensic Taphonomy, 2nd ed.; Pokines, J.T., L’Abbe, E.N., Symes, S.A., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 41–78. [Google Scholar]
- Damann, F.E.; Jans, M.M.E. Microbes, anthropology, and bones. In Forensic Microbiology, 1st ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 312–327. [Google Scholar]
- Forbes, S.L.; Perrault, K.A.; Comstock, J.L. Microscopic post-mortem changes: The chemistry of decomposition. In Taphonomy of Human Remains: Forensic Analysis of the Dead and the Depositional Environment, 1st ed.; Schotsmans, E.M.J., Márquez-Grant, N., Forbes, S.L., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 26–38. [Google Scholar]
- Cockle, D.L.; Bell, L.S. Human decomposition and the reliability of a ‘universal’ model for postmortem interval estimations. Forensic Sci. Int. 2015, 253, 136.e1–136.e9. [Google Scholar] [CrossRef] [PubMed]
- Carter, D.O.; Yellowlees, D.; Tibbett, M. Cadaver decomposition in terrestrial ecosystems. Naturwissenschaften 2007, 94, 12–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megyesi, M.S.; Nawrocki, S.P.; Haskell, N.H. Using accumulated degree-days to estimate the postmortem interval from decomposed human remains. J. Forensic Sci. 2005, 50, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Micozzi, M.S. Postmortem Change in Human and Animal Remains: A Systematic Approach; Charles C. Thomas: Springfield, IL, USA, 1991. [Google Scholar]
- Simmons, T. Post-mortem interval estimation: An overview of techniques. In Taphonomy of Human Remains: Forensic Analysis of the Dead and the Depositional Environment; Schotsmans, E.M.J., Márquez-Grant, N., Forbes, S.L., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 134–142. [Google Scholar]
- Matuszewski, S.; Konwerski, S.; Frątczak, K.; Szafałowicz, M. Effect of body mass and clothing on decomposition of pig carcasses. Int. J. Legal Med. 2014, 128, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Mann, R.W.; Bass, W.M.; Meadows, L. Time since death and decomposition of the human body: Variables and observations in case and experimental field studies. J. Forensic Sci. 1990, 35, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, W.C.; Bass, W.M. Decomposition of buried bodies and methods that may aid in their location. J. Forensic Sci. 1985, 30, 836–852. [Google Scholar] [CrossRef]
- Latham, K.E.; Madonna, M.E.; Hipp, J.L. DNA survivability in skeletal remains. In Manual of Forensic Taphonomy; Pokines, J.T., L’Abbe, E.N., Symes, S.A., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 555–580. [Google Scholar]
- Ottoni, C.; Bekaert, B.; Decorte, R. DNA degradation: Current knowledge and progress in DNA analysis. In Taphonomy of Human Remains: Forensic Analysis of the Dead and the Depositional Environment; Schotsmans, E.M.J., Márquez-Grant, N., Forbes, S.L., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 65–80. [Google Scholar]
- Bell, L.S.; Skinner, M.F.; Jones, S.J. The speed of postmortem change to the human skeleton and its taphonomic significance. Forensic Sci. Int. 1996, 82, 129–140. [Google Scholar] [CrossRef]
- Procopio, N.; Mein, C.A.; Starace, S.; Bonicelli, A.; Williams, A. Bone diagenesis in short timescales: Insights from an exploratory proteomic analysis. Biology 2021, 10, 460. [Google Scholar] [CrossRef]
- Ross, A.H.; Hale, A.R. Decomposition of juvenile-sized remains: A macro- and microscopic perspective. Forensic Sci. Res. 2018, 3, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Cockle, D.L.; Bell, L.S. The environmental variables that impact human decomposition in terrestrially exposed contexts within Canada. Sci. Justice 2017, 57, 107–117. [Google Scholar] [CrossRef]
- Meyer, J.; Anderson, B.; Carter, D.O. Seasonal variation of carcass decomposition and gravesoil chemistry in a cold (Dfa) climate. J. Forensic Sci. 2013, 58, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Shirley, N.R.; Wilson, R.J.; Jantz, L.M. Cadaver use at the University of Tennessee’s anthropological research facility. Clin. Anat. 2011, 24, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Galloway, A.; Birkby, W.H.; Jones, A.M.; Henry, T.E.; Parks, B.O. Decay rates of human remains in an arid environment. J. Forensic Sci. 1989, 34, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Booth, T.J.; Redfern, R.C.; Gowland, R.L. Immaculate conceptions: Micro-CT analysis of diagenesis in Romano-British infant skeletons. J. Archaeol. Sci. 2016, 74, 124–134. [Google Scholar] [CrossRef]
- White, L.; Booth, T.J. The origin of bacteria responsible for bioerosion to the internal bone microstructure: Results from experimentally-deposited pig carcasses. Forensic Sci. Int. 2014, 239, 92–102. [Google Scholar] [CrossRef]
- Smith, C.I.; Nielsen-Marsh, C.M.; Jans, M.M.E.; Collins, M.J. Bone diagenesis in the European Holocene I: Patterns and mechanisms. J. Archaeol. Sci. 2007, 34, 1485–1493. [Google Scholar] [CrossRef]
- Jans, M.M.E.; Nielsen-Marsh, C.M.; Smith, C.I.; Collins, M.J.; Kars, H. Characterisation of microbial attack on archaeological bone. J. Archaeol. Sci. 2004, 31, 87–95. [Google Scholar] [CrossRef]
- Nielsen-Marsh, C.; Gernaey, A.; Turner-Walker, G.; Hedges, R.; Pike, A.; Collins, M. The chemical degradation of bone. In Human Osteology: In Archaeology and Forensic Science; Cambridge University Press: Cambridge, UK, 2000; pp. 439–454. [Google Scholar]
- Hedges, R.E.M.; Millard, A.R.; Pike, A.W.G. Measurements and relationships of diagenetic alteration of bone from three archaeological sites. J. Archaeol. Sci. 1995, 22, 201–209. [Google Scholar] [CrossRef]
- Pechal, J.L.; Crippen, T.L.; Tarone, A.M.; Lewis, A.J.; Tomberlin, J.K.; Benbow, M.E. Microbial community functional change during vertebrate carrion decomposition. PLoS ONE 2013, 8, e79035. [Google Scholar] [CrossRef]
- Keenan, S.W. From bone to fossil: A review of the diagenesis of bioapatite. Am. Mineral. 2016, 101, 1943–1951. [Google Scholar] [CrossRef]
- Hedges, R.E.M. Bone diagenesis: An overview of processes. Archaeometry 2002, 44, 319–328. [Google Scholar] [CrossRef]
- Turner-Walker, G. The chemical and microbial degradation of bones and teeth. In Advances in Human Palaeopathology; John Wiley & Sons, Ltd.: Chichester, UK, 2007; pp. 3–29. [Google Scholar]
- Child, A.M. Microbial taphonomy of archaeological bone. Stud. Conserv. 1995, 40, 19–30. [Google Scholar] [CrossRef]
- Collins, M.J.; Nielsen–Marsh, C.M.; Hiller, J.; Smith, C.I.; Roberts, J.P.; Prigodich, R.V.; Wess, T.J.; Csapò, J.; Millard, A.R.; Turner–Walker, G. The survival of organic matter in bone: A review. Archaeometry 2002, 44, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Berna, F.; Matthews, A.; Weiner, S. Solubilities of bone mineral from archaeological sites: The recrystallization window. J. Archaeol. Sci. 2004, 31, 867–882. [Google Scholar] [CrossRef]
- Grupe, G. Preservation of collagen in bone from dry, sandy soil. J. Archaeol. Sci. 1995, 22, 193–199. [Google Scholar] [CrossRef]
- Hall, B.K. Bones and Cartilage, 2nd ed.; Elsevier Academic Press: London, UK, 2015. [Google Scholar]
- Tripp, J.A.; Squire, M.E.; Hedges, R.E.M.; Stevens, R.E. Use of micro-computed tomography imaging and porosity measurements as indicators of collagen preservation in archaeological bone. Palaeogeogr. Palaeoclim. Palaeoecol. 2018, 511, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Trueman, C.N.; Palmer, M.R.; Field, J.; Privat, K.; Ludgate, N.; Chavagnac, V.; Eberth, D.A.; Cifelli, R.; Rogers, R.R. Comparing rates of recrystallisation and the potential for preservation of biomolecules from the distribution of trace elements in fossil bones. Comptes. Rendus. Palevol. 2008, 7, 145–158. [Google Scholar] [CrossRef]
- Jans, M.M.E. Microbial bioerosion of bone—A review. In Current Developments in Bioerosion; Wisshak, M., Tapanila, L., Eds.; Erlangen Earth Conference Series; Springer: Berlin/Heidelberg, Germany, 2008; pp. 397–413. [Google Scholar]
- Trueman, C.N.; Privat, K.; Field, J. Why do crystallinity values fail to predict the extent of diagenetic alteration of bone mineral? Palaeogeogr. Palaeoclim. Palaeoecol. 2008, 266, 160–167. [Google Scholar] [CrossRef]
- Keenan, S.W.; Engel, A.S.; Roy, A.; Lisa Bovenkamp-Langlois, G. Evaluating the consequences of diagenesis and fossilization on bioapatite lattice structure and composition. Chem. Geol. 2015, 413, 18–27. [Google Scholar] [CrossRef]
- Hedges, R.E.M.; Millard, A.R. Bones and Groundwater: Towards the modelling of diagenetic processes. J. Archaeol. Sci. 1995, 22, 155–164. [Google Scholar] [CrossRef]
- Nielsen-Marsh, C.M.; Hedges, R.E.M. Bone porosity and the use of mercury intrusion porosimetry in bone diagenesis studies. Archaeometry 1999, 41, 165–174. [Google Scholar] [CrossRef]
- Boaks, A.; Siwek, D.; Mortazavi, F. The temporal degradation of bone collagen: A histochemical approach. Forensic Sci. Int. 2014, 240, 104–110. [Google Scholar] [CrossRef]
- Hackett, C.J. Microscopical focal destruction (tunnels) in exhumed human bones. Med. Sci. Law 1981, 21, 243–265. [Google Scholar] [CrossRef] [PubMed]
- Nielsen-Marsh, C.M.; Smith, C.I.; Jans, M.M.E.; Nord, A.; Kars, H.; Collins, M.J. Bone diagenesis in the European Holocene II: Taphonomic and environmental considerations. J. Archaeol. Sci. 2007, 34, 1523–1531. [Google Scholar] [CrossRef]
- Jackes, M.; Sherburne, R.; Lubell, D.; Barker, C.; Wayman, M. Destruction of microstructure in archaeological bone: A case study from Portugal. Int. J. Osteoarchaeol. 2001, 11, 415–432. [Google Scholar] [CrossRef]
- Garland, N.A. Microscopical analysis of fossil bone. Appl. Geochem. 1989, 4, 215–229. [Google Scholar] [CrossRef]
- Manifold, B.M. Intrinsic and extrinsic factors involved in the preservation of non-adult skeletal remains in archaeology and forensic science. Bull. Int. Assoc. Paleodontol. 2012, 6, 51–69. [Google Scholar]
- Manifold, B.M. Bone mineral density in children from anthropological and clinical sciences: A review. Anthropol. Rev. 2014, 77, 111–135. [Google Scholar] [CrossRef] [Green Version]
- Djuric, M.; Djukic, K.; Milovanovic, P.; Janovic, A.; Milenkovic, P. Representing children in excavated cemeteries: The intrinsic preservation factors. Antiquity 2011, 85, 250–262. [Google Scholar] [CrossRef]
- Guy, H.; Masset, C.; Baud, C.-A. Infant taphonomy. Int. J. Osteoarchaeol. 1997, 7, 221–229. [Google Scholar] [CrossRef]
- Caruso, V.; Marinoni, N.; Diella, V.; Possenti, E.; Mancini, L.; Cantaluppi, M.; Berna, F.; Cattaneo, C.; Pavese, A. Diagenesis of juvenile skeletal remains: A multimodal and multiscale approach to examine the post-mortem decay of children’s bones. J. Archaeol. Sci. 2021, 135, 105477. [Google Scholar] [CrossRef]
- Buckberry, J. Missing, presumed buried? Bone diagenesis and the under-representation of Anglo-Saxon children. Assem. Univ. Sheff. Grad. Stud. J. Archaeol. 2000, 5, 1–18. [Google Scholar]
- Jans, M.M.E. Microscopic destruction of bone. In Manual of Forensic Taphonomy; Pokines, J.T., L’Abbe, E.N., Symes, S.A., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 23–39. [Google Scholar]
- Wedl, C. Über Einen Im Zahnbein Und Knochen Keimenden Pilz. Concerning a fungus germinating from dentine and bone. Sitz. Der Kais. Akad. Der Wiss. Math.-Nat. Cl. 1864, 50, 171–193. [Google Scholar]
- Stokes, K.L.; Forbes, S.L.; Benninger, L.A.; Carter, D.O.; Tibbett, M. Decomposition studies using animal models in contrasting environments: Evidence from temporal changes in soil chemistry and microbial activity. In Criminal and Environmental Soil Forensics; Ritz, K., Dawson, L., Miller, D., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 357–377. [Google Scholar]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. HESS 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Goldschlager, T.; Abdelkader, A.; Kerr, J.; Boundy, I.; Jenkin, G. Undecalcified bone preparation for histology, histomorphometry and fluorochrome analysis. J. Vis. Exp. 2010, 35, 1707. [Google Scholar] [CrossRef] [Green Version]
- JMP, version 16; SAS Institute Inc.: Cary, NC, USA, 2021.
- Turner-Walker, G.; Jans, M. Reconstructing taphonomic histories using histological analysis. Palaeogeogr. Palaeoclim. Palaeoecol. 2008, 266, 227–235. [Google Scholar] [CrossRef]
- Turner-Walker, G. Light at the end of the tunnels? the origins of microbial bioerosion in mineralised collagen. Palaeogeogr. Palaeoclim. Palaeoecol. 2019, 529, 24–38. [Google Scholar] [CrossRef]
- Fernández-Jalvo, Y.; Andrews, P.; Pesquero, D.; Smith, C.; Marín-Monfort, D.; Sánchez, B.; Geigl, E.-M.; Alonso, A. Early bone diagenesis in temperate environments part I: Surface features and histology. Palaeogeogr. Palaeoclim. Palaeoecol. 2010, 288, 62–81. [Google Scholar] [CrossRef]
- Moore, R.E.; Townsend, S.D. Temporal development of the infant gut microbiome. Open Biol. 2019, 9, 190128. [Google Scholar] [CrossRef] [Green Version]
- Trueman, C.N.; Martill, D.M. The long–term survival of bone: The role of bioerosion. Archaeometry 2002, 44, 371–382. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample | Start Date | ADD | ADD (Buried) | Mean Soil Temperature | Mean Soil Moisture |
|---|---|---|---|---|---|
| Summer 2013 | 22 May 2013 | 11,966.61 | 12,558.61 | 61.24 | 0.26 |
| Fall 2013 | 13 September 2013 | 9163.86 | 9630.06 | 59.64 | 0.27 |
| Winter 2013 | 5 December 2013 | 7965.17 | 8252.11 | 58.06 | 0.28 |
| Spring 2014 | 30 March 2014 | 7246.89 | 7494.78 | 61.17 | 0.31 |
| Summer 2014 | 6 June 2014 | 5969.67 | 6171.67 | 60.48 | 0.30 |
| Fall 2014 | 27 September 2014 | 3184.39 | 3378.39 | 55.96 | 0.33 |
| Winter 2014 | 20 December 2014 | 2153.72 | 2221.56 | 52.76 | 0.33 |
| Spring 2015 | 13 March 2015 | 1753.69 | 1748.94 | 59.35 | 0.33 |
| Score | Category | Definition |
|---|---|---|
| 0 | No damage | Microstructure appears intact, with no enlarged osteocyte lacunae apparent. |
| 1 | Minor damage | Microstructure shows enlarged osteocyte lacunae not coincident with exogenous staining but no amalgamations. |
| 2 | Minor damage with amalgamation | Microstructure shows amalgamations of enlarged osteocyte lacunae not coincident with exogenous staining, but no coalescence or damage to more than one region of the microstructure. |
| 3 | Major damage | Microstructure shows amalgamations of enlarged osteocyte lacunae not coincident with exogenous staining, and coalescence of amalgamations across more than one region of the microstructure. |
| Deposition | Probability | TI | ADD | Time Equivalent | BIC |
|---|---|---|---|---|---|
| Bag fetal | 0.63 | 2 | 6860.15 | 355 days | 18.050 |
| Blanket fetal | 0.31 | 2 | 6860.15 | 355 days | 21.539 |
| Buried juvenile | 0.25 | 2.5 | 7153.78 | 394 days | 7.483 |
| Surface juvenile | 0.50 | 2 | 6860.15 | 355 days | 16.740 |
| Deposition | Model | Standard Deviation | Log-Likelihood |
|---|---|---|---|
| Bag fetal | TI = 0.222 + (5.407 × 10−5) × ADD | 0.280 | 6.106 |
| Blanket fetal | TI = 0.536 + (4.164 × 10−5) × ADD | 0.199 | 7.650 |
| Buried juvenile | TI = 0.621 + (5.032 × 10−5) × ADD | 0.069 | 0.622 |
| Surface juvenile | TI = 0.530 + (2.420 × 10−5) × ADD | 0.176 | 5.251 |
| Deposition | Model Parameter | Log Worth | p-Value (α = 0.05) |
|---|---|---|---|
| Bag fetal (R2 = 0.83) | Mean Soil Temperature | 1.251 | 0.06 |
| Mean Soil Moisture | 0.538 | 0.29 | |
| ADD | 0.432 | 0.37 | |
| Blanket fetal (R2 = 0.54) | Mean Soil Temperature | 0.544 | 0.29 |
| Mean Soil Moisture | 0.312 | 0.44 | |
| ADD | 0.356 | 0.49 | |
| Buried juvenile (R2 = 0.81) | Mean Soil Temperature | 0.425 | 0.38 |
| Mean Soil Moisture | 0.127 | 0.75 | |
| ADD | 0.495 | 0.32 | |
| Surface juvenile (R2 = 0.51) | Mean Soil Temperature | 0.708 | 0.20 |
| Mean Soil Moisture | 0.069 | 0.85 | |
| ADD | 0.359 | 0.44 |
| Variable | Bag Fetal | Blanket Fetal | Buried Juvenile | Surface Juvenile |
|---|---|---|---|---|
| Mean Soil Temperature | 0.85 (0.016) | 0.67 (0.067) | 0.72 (0.043) | −0.12 (0.782) |
| Mean Soil Moisture | −0.42 (0.350) | −0.45 (0.267) | −0.81 (0.014) | −0.43 (0.290) |
| ADD | 0.49 (0.260) | 0.54 (0.168) | 0.87 (0.005) | 0.50 (0.253) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hale, A.R.; Ross, A.H. Investigating the Timing and Extent of Juvenile and Fetal Bone Diagenesis in a Temperate Environment. Biology 2023, 12, 403. https://doi.org/10.3390/biology12030403
Hale AR, Ross AH. Investigating the Timing and Extent of Juvenile and Fetal Bone Diagenesis in a Temperate Environment. Biology. 2023; 12(3):403. https://doi.org/10.3390/biology12030403
Chicago/Turabian StyleHale, Amanda R., and Ann H. Ross. 2023. "Investigating the Timing and Extent of Juvenile and Fetal Bone Diagenesis in a Temperate Environment" Biology 12, no. 3: 403. https://doi.org/10.3390/biology12030403