
Effect of PACAP/PAC1R on Follicle Development of Djungarian Hamster (Phodopus sungorus) with the Variation of Ambient Temperatures


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Tissue Preparation
2.2. Microstructure Observation of Ovarian Follicles
2.3. Total RNA Extraction and RT-PCR
2.4. Gene Cloning
2.5. Real-Time Fluorescence Quantitative PCR
2.6. FSH Hormone Content Determination
2.7. Statistical Analysis
3. Results
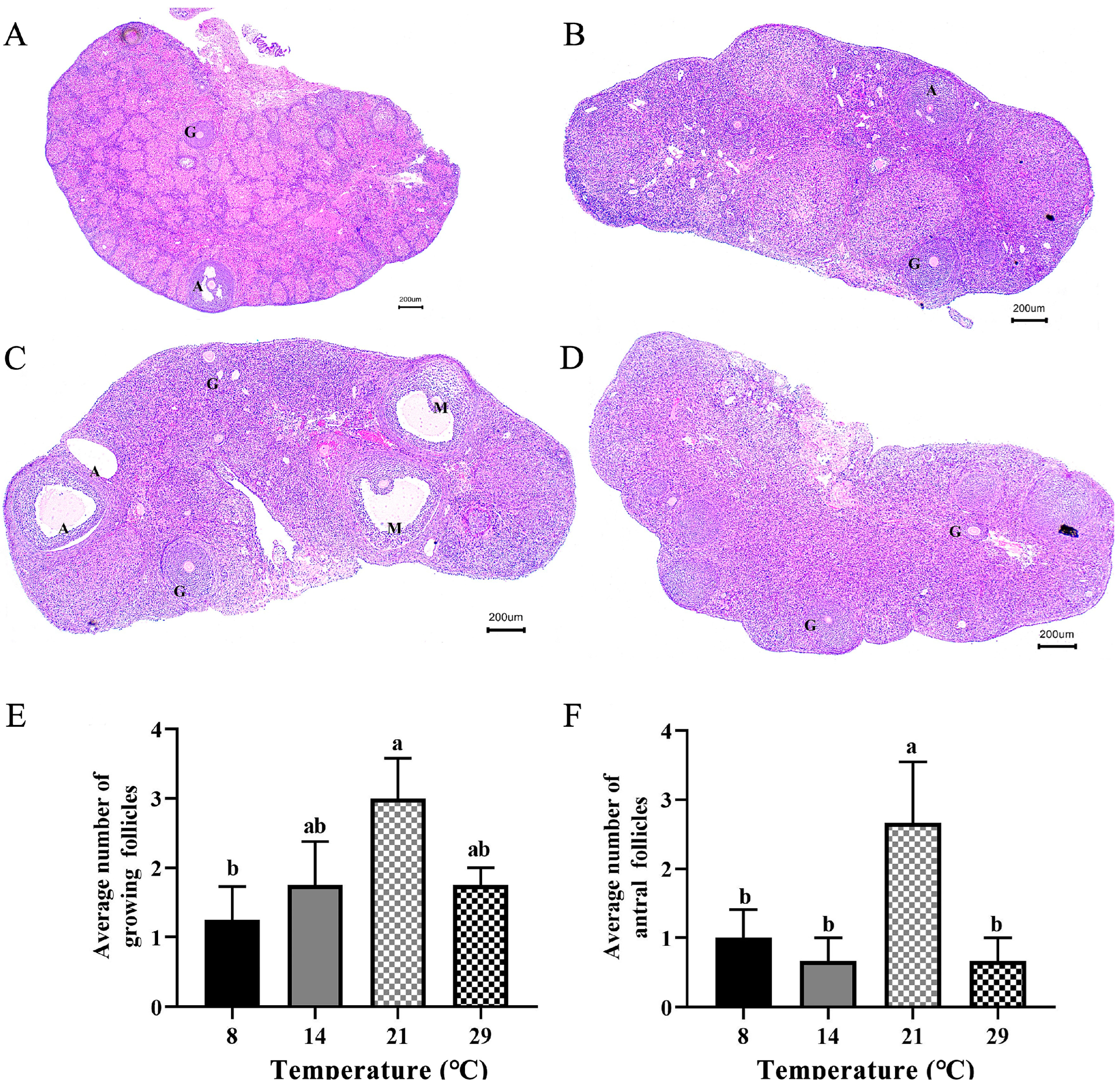
3.1. Differences in the Number of Growing Follicles and Antral Follicles at Different Temperatures
3.2. Characterization of PACAP and PAC1R
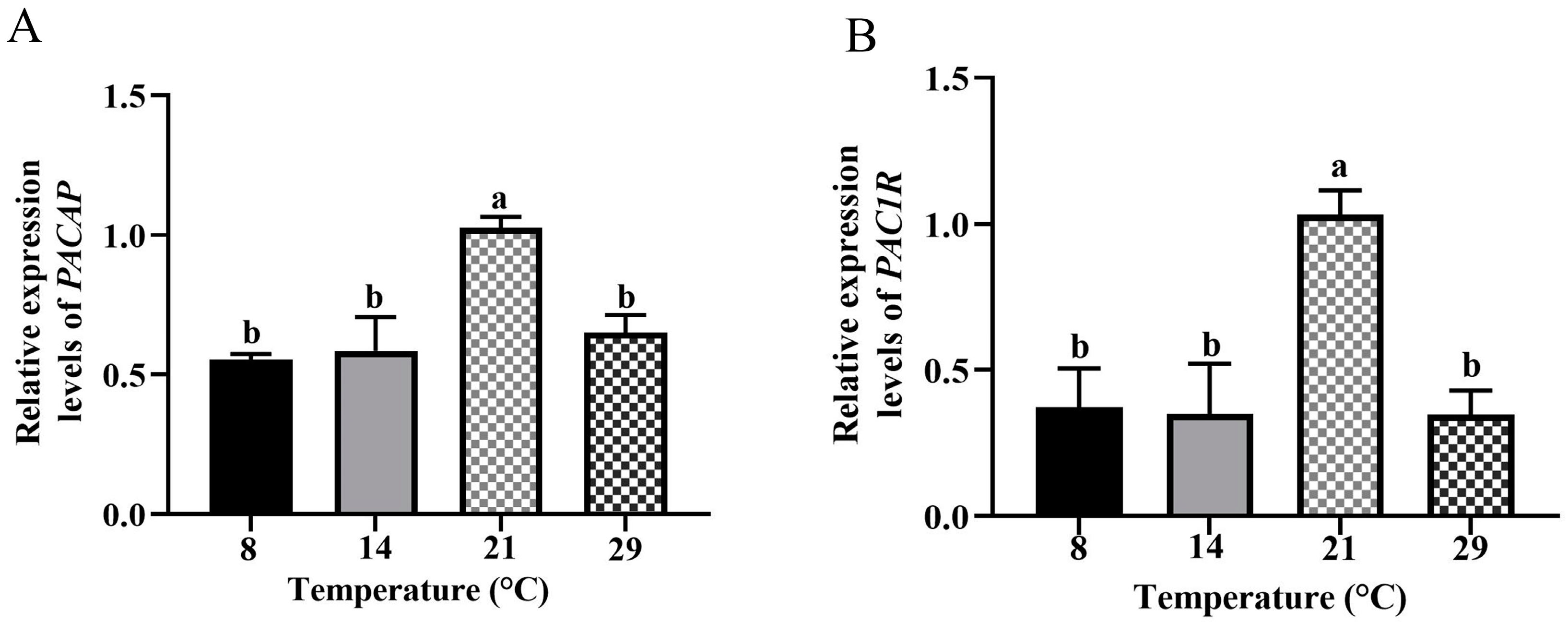
3.3. Differential mRNA Expression Levels of PACAP and PAC1R in the Hypothalamus of Female Phodopus Sungorus at Different Ambient Temperatures
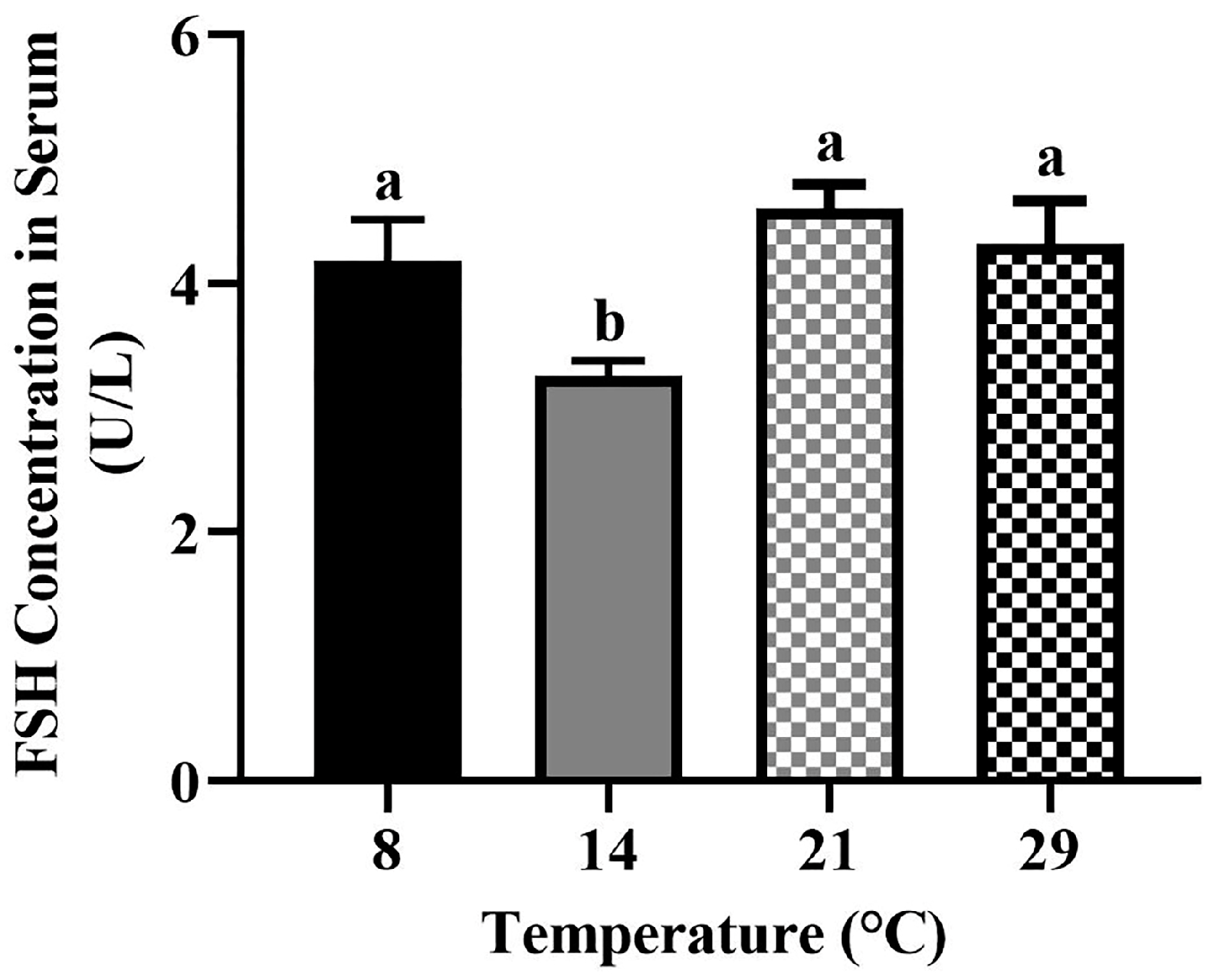
3.4. The Serum Concentration of FSH
3.5. Analysis of Correlation between the PACAP/PAC1R Expression and the Number of Growing Follicles and Antral Follicles
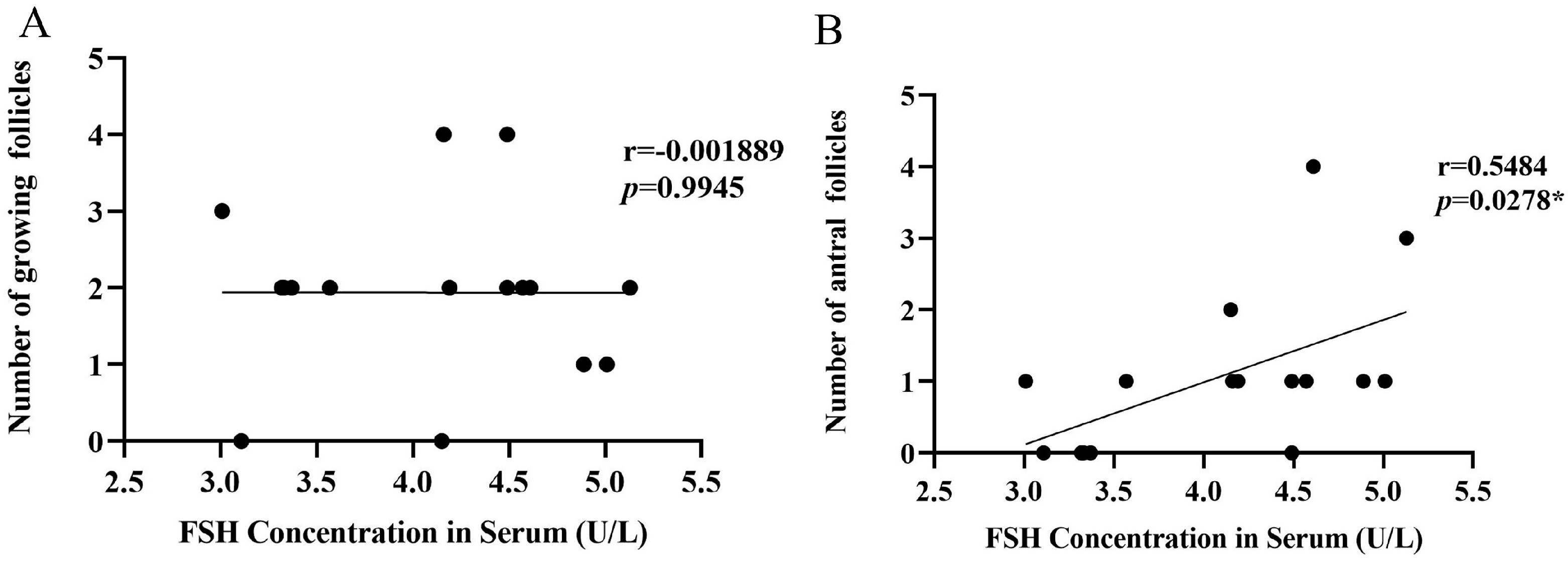
3.6. Analysis of Correlation between FSH Concentration in Serum and the Number of Growing Follicles and Antral Follicles
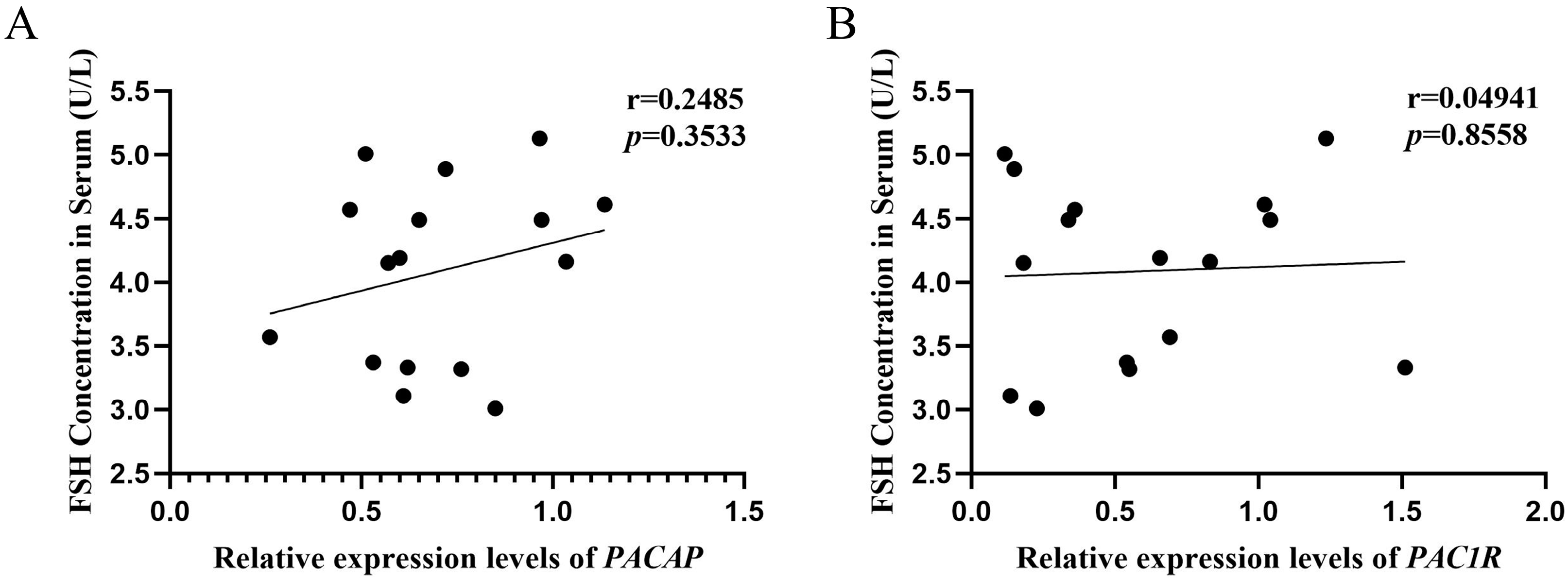
3.7. Analysis of Correlation between the PACAP/PAC1R Expression and FSH Concentration in Serum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moffatt-Blue, C.S.; Sury, J.J.; Young, K.A. Short photoperiod-induced ovarian regression is mediated by apoptosis in Siberian hamsters (Phodopus sungorus). Reproduction 2006, 131, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Donadeu, F.X.; Watson, E.D. Seasonal changes in ovarian activity: Lessons learnt from the horse. Anim. Reprod. Sci. 2007, 100, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.H.; Zhong, Y.Z.; Lin, L.L.; Zhang, G.L.; Su, W.H.; Ni, W.L.; Qu, M.J.; Chen, H.L. Effect of Photoperiod on Longevity, Food Consumption, and Reproduction of Holotrichia oblita (Coleoptera: Scarabaeidae). Environ. Entomol. 2021, 50, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Salverson, T.J.; McMichael, G.E.; Sury, J.J.; Shahed, A.; Young, K.A. Differential expression of matrix metalloproteinases during stimulated ovarian recrudescence in Siberian hamsters (Phodopus sungorus). Gen. Comp. Endocrinol. 2008, 155, 749–761. [Google Scholar] [CrossRef] [Green Version]
- Shahed, A.; McMichael, C.F.; Young, K.A. Rapid changes in ovarian mRNA induced by brief photostimulation in Siberian hamsters (Phodopus sungorus). J. Exp. Zool. A Ecol. Genet. Physiol. 2015, 323, 627–636. [Google Scholar] [CrossRef] [Green Version]
- Bock, S.L.; Chow, M.I.; Forsgren, K.L.; Lema, S.C. Widespread alterations to hypothalamic-pituitary-gonadal (HPG) axis signaling underlie high temperature reproductive inhibition in the eurythermal sheepshead minnow (Cyprinodon variegatus). Mol. Cell. Endocrinol. 2021, 537, 111447. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, X.; Wang, W.; Zhao, L.; Gao, L.; Yang, W. Annual variation in the reproductive hormone and behavior rhythm in a population of the Asian short-toed lark: Can spring temperature influence activation of the HPG axis of wild birds? Horm. Behav. 2017, 95, 76–84. [Google Scholar] [CrossRef]
- Yom-Tov, Y. Global warming and body mass decline in Israeli passerine birds. Proc. R. Soc. Biol. Sci. Ser. B 2001, 268, 947–952. [Google Scholar] [CrossRef] [Green Version]
- Frehn, J.L.; Liu, C.C. Effects of temperature, photoperiod, and hibernation on the testes of golden hamsters. J. Exp. Zool. 1970, 174, 317–323. [Google Scholar] [CrossRef]
- Caro, S.P.; Schaper, S.V.; Hut, R.A.; Ball, G.F.; Visser, M.E. The case of the missing mechanism: How does temperature influence seasonal timing in endotherms? PLoS Biol. 2013, 11, e1001517. [Google Scholar] [CrossRef]
- Qiang, J.; Tao, Y.-F.; Zhu, J.-H.; Lu, S.-Q.; Cao, Z.-M.; Ma, J.-L.; He, J.; Xu, P. Effects of heat stress on follicular development and atresia in Nile tilapia (Oreochromis niloticus) during one reproductive cycle and its potential regulation by autophagy and apoptosis. Aquaculture 2022, 555, 738171. [Google Scholar] [CrossRef]
- Miyata, A.; Arimura, A.; Dahl, R.R.; Minamino, N.; Uehara, A.; Jiang, L.; Culler, M.D.; Coy, D.H. Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells. Biochem. Biophys. Res. Commun. 1989, 164, 567–574. [Google Scholar] [CrossRef]
- Levy, G.; Degani, G. The role of brain peptides in the reproduction of blue gourami males (Trichogaster trichopterus). J. Exp. Zool. Part A 2013, 319, 461–470. [Google Scholar] [CrossRef]
- Ai, N.; Liu, L.; Lau, E.S.; Tse, A.C.; Ge, W. Separation of Oocyte and Follicle Layer for Gene Expression Analysis in Zebrafish. Methods Mol. Biol. 2021, 2218, 1–9. [Google Scholar] [CrossRef]
- Pisegna, J.R.; Wank, S.A. Molecular cloning and functional expression of the pituitary adenylate cyclase-activating polypeptide type I receptor. Proc. Natl. Acad. Sci. USA 1993, 90, 6345–6349. [Google Scholar] [CrossRef] [Green Version]
- Velkeniers, B.; Zheng, L.; Kazemzadeh, M.; Robberecht, P.; Vanhaelst, L.; Hooghe-Peters, E.L. Effect of pituitary adenylate cyclase-activating polypeptide 38 on growth hormone and prolactin expression. J. Endocrinol. 1994, 143, 1–11. [Google Scholar] [CrossRef]
- Kanasaki, H.; Oride, A.; Kyo, S. Role of pituitary adenylate cyclase-activating polypeptide in modulating hypothalamus-pituitary neuroendocrine functions in mouse cell models. J. Neuroendocrinol. 2015, 27, 1–7. [Google Scholar] [CrossRef]
- Gonzalez, B.J.; Basille, M.; Vaudry, D.; Fournier, A.; Vaudry, H. Pituitary adenylate cyclase-activating polypeptide. Ann. Endocrinol. 1998, 59, 364–405. [Google Scholar]
- Stroth, N.; Holighaus, Y.; Ait-Ali, D.; Eiden, L.E. PACAP: A master regulator of neuroendocrine stress circuits and the cellular stress response. Ann. N. Y. Acad. Sci. 2011, 1220, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Mercer, K.B.; Dias, B.; Shafer, D.; Maddox, S.A.; Mulle, J.G.; Hu, P.; Walton, J.; Ressler, K.J. Functional evaluation of a PTSD-associated genetic variant: Estradiol regulation and ADCYAP1R1. Transl. Psychiatr. 2016, 6, e978. [Google Scholar] [CrossRef] [Green Version]
- Szentleleky, E.; Szegeczki, V.; Karanyicz, E.; Hajdu, T.; Tamas, A.; Toth, G.; Zakany, R.; Reglodi, D.; Juhasz, T. Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) Reduces Oxidative and Mechanical Stress-Evoked Matrix Degradation in Chondrifying Cell Cultures. Int. J. Mol. Sci. 2019, 20, 168. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Piper, S.J.; Zhao, P.; Miller, L.J.; Wootten, D.; Sexton, P.M. Targeting VIP and PACAP Receptor Signaling: New Insights into Designing Drugs for the PACAP Subfamily of Receptors. Int. J. Mol. Sci. 2022, 23, 8069. [Google Scholar] [CrossRef] [PubMed]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef] [PubMed]
- Shabb, J.B. Physiological substrates of cAMP-dependent protein kinase. Chem. Rev. 2001, 101, 2381–2411. [Google Scholar] [CrossRef] [PubMed]
- Fowkes, R.C.; Burch, J.; Burrin, J.M. Stimulation of extracellular signal-regulated kinase by pituitary adenylate cyclase-activating polypeptide in alpha T3-1 gonadotrophs. J. Endocrinol. 2001, 171, R5–R10. [Google Scholar] [CrossRef] [Green Version]
- Yeh, D.M.; Coss, D. PACAP induces FSHbeta gene expression via EPAC. Mol. Cell. Endocrinol. 2019, 492, 110438. [Google Scholar] [CrossRef]
- Koppan, M.; Nagy, Z.; Bosnyak, I.; Reglodi, D. Female reproductive functions of the neuropeptide PACAP. Front. Endocrinol. 2022, 13, 982551. [Google Scholar] [CrossRef]
- Park, K.M.; Kim, K.J.; Jin, M.; Han, Y.; So, K.H.; Hyun, S.H. The use of pituitary adenylate cyclase-activating polypeptide in the pre-maturation system improves in vitro developmental competence from small follicles of porcine oocytes. Asian-Australas J. Anim. Sci. 2019, 32, 1844–1853. [Google Scholar] [CrossRef] [Green Version]
- Reglodi, D.; Tamas, A.; Koppan, M.; Szogyi, D.; Welke, L. Role of PACAP in Female Fertility and Reproduction at Gonadal Level - Recent Advances. Front. Endocrinol. 2012, 3, 155. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.P., Jr.; Yang, R.Q.; Winters, S.J. Targeted pituitary overexpression of pituitary adenylate-cyclase activating polypeptide alters postnatal sexual maturation in male mice. Endocrinology 2012, 153, 1421–1434. [Google Scholar] [CrossRef] [Green Version]
- Gras, S.; Host, E.; Fahrenkrug, J. Role of pituitary adenylate cyclase-activating peptide (PACAP) in the cyclic recruitment of immature follicles in the rat ovary. Regul. Pept. 2005, 128, 69–74. [Google Scholar] [CrossRef]
- Merdan, H.; Duman, O.; Akin, O.; Celik, C. Allee effects on population dynamics in continuous (overlapping) case. Chaos Solitons Fractals 2009, 39, 1994–2001. [Google Scholar] [CrossRef]
- Figala, J.; Hoffmann, K.; Goldau, G. The annual cycle in the Djungarian Hamster Phodopus sungorus Pallas. Oecologia 1973, 12, 89–118. [Google Scholar] [CrossRef]
- Hoffmann, K. Photoperiod, pineal, melatonin and reproduction in hamsters. Prog. Brain Res. 1979, 52, 397–415. [Google Scholar] [CrossRef]
- Yue, C.; Guo, Q.W.; Zhang, Z.R.; Li, X.; Man, D.H.; Yuan, S.; Fu, H.P.; Wu, X.D.; Jin, G.; Liu, J.W.; et al. Trophic niche of Brandt’s voles (Lasiopodomys brandtii) and their interspecific relationships with other common rodents in a typical steppe, Inner Mongolia. Acta Theriol. Sin. 2020, 40, 424–434. [Google Scholar] [CrossRef]
- Ross, P.D. Phodopus sungorus. Poxford J. 1998, 595, 1–9. [Google Scholar] [CrossRef]
- Lerchl, A. Breeding of Djungarian hamsters (Phodopus sungorus): Influence of parity and litter size on weaning success and offspring sex ratio. Lab. Anim. 1995, 29, 172–176. [Google Scholar] [CrossRef]
- Liu, J.S.; Wang, D.H.; Sun, R.Y. Metabolism and thermoregulation in three species of rodent from Northeastern China. J. Therm. Biol. 2004, 29, 177–183. [Google Scholar] [CrossRef]
- Song, Z.G.; Wang, D.H. Metabolism and thermoregulation in the striped hamster Cricetulus barabensis. J. Therm. Biol. 2003, 28, 509–514. [Google Scholar] [CrossRef]
- Tsutsui, K.; Kawashima, S.; Masuda, A.; Oishi, T. Effects of photoperiod and temperature on the binding of follicle-stimulating hormone (FSH) to testicular preparations and plasma FSH concentration in the Djungarian hamster, Phodopus sungorus. Endocrinology 1988, 122, 1094–1102. [Google Scholar] [CrossRef]
- Masuda, A.; Oishi, T. Effects of photoperiod, temperature and testosterone-treatment on plasma T3 and T4 levels in the Djungarian hamster, Phodopus sungorus. Experientia 1989, 45, 102–103. [Google Scholar] [CrossRef] [PubMed]
- Crooks, P.V.; O’Reilly, C.B.; Owens, P.D. Microscopy of the dentine of enamel-free areas of rat molar teeth. Arch. Oral Biol. 1983, 28, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Ledevin, R.; Quere, J.P.; Renaud, S. Morphometrics as an insight into processes beyond tooth shape variation in a bank vole population. PLoS ONE 2010, 5, e15470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pevet, P.; Vivien-Roels, B.; Masson-Pevet, M. Low temperature in the golden hamster accelerates the gonadal atrophy induced by short photoperiod but does not affect the daily pattern of melatonin secretion. J. Neural Transm. 1989, 76, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, J.H.; Mou, J.J.; Kong, X.T.; Wu, M.; Xue, H.L.; Xu, L.X. Photoperiod Affects Harderian Gland Morphology and Secretion in Female Cricetulus barabensis: Autophagy, Apoptosis, and Mitochondria. Front. Physiol. 2020, 11, 408. [Google Scholar] [CrossRef]
- Tian, M.Y. Microstructure Observation on the Follicular Development of Blue Fox during the Estrous Period. J. Anhui Agric. Sci. 2011, 33, 221. [Google Scholar] [CrossRef]
- Rutledge, R.G.; Stewart, D. A kinetic-based sigmoidal model for the polymerase chain reaction and its application to high-capacity absolute quantitative real-time PCR. BMC Biotechnol. 2008, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; Heijne, G.V.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Rost, B.; Yachdav, G.; Liu, J. The PredictProtein server. Nucleic Acids Res. 2004, 32, W321–W326. [Google Scholar] [CrossRef] [Green Version]
- Bairoch, A.; Bucher, P. PROSITE: Recent developments. Nucleic Acids Res. 1994, 22, 3583–3589. [Google Scholar]
- Dejda, A.; Sokolowska, P.; Nowak, J.Z. Neuroprotective potential of three neuropeptides PACAP, VIP and PHI. Pharm. Rep. 2005, 57, 307–320. [Google Scholar]
- Egbert, J.R.; Yee, S.P.; Jaffe, L.A. Luteinizing hormone signaling phosphorylates and activates the cyclic GMP phosphodiesterase PDE5 in mouse ovarian follicles, contributing an additional component to the hormonally induced decrease in cyclic GMP that reinitiates meiosis. Dev. Biol. 2018, 435, 6–14. [Google Scholar] [CrossRef]
- Vaudry, D.; Gonzalez, B.J.; Basille, M.; Yon, L.; Fournier, A.; Vaudry, H. Pituitary adenylate cyclase-activating polypeptide and its receptors: From structure to functions. Pharmacol. Rev. 2000, 52, 269–324. [Google Scholar]
- Bronson, F.H. Climate change and seasonal reproduction in mammals. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 3331–3340. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Fu, X.; Ge, S.; Sun, T.; Sheng, Z. Differential expression of matrix metalloproteinases and tissue-derived inhibitors of metalloproteinase in fetal and adult skins. Int. J. Biochem. Cell Biol. 2007, 39, 997–1005. [Google Scholar] [CrossRef]
- Xu, X.; Liu, X.; Ma, S.; Xu, Y.; Xu, Y.; Guo, X.; Li, D. Association of Melatonin Production with Seasonal Changes, Low Temperature, and Immuno-Responses in Hamsters. Molecules 2018, 23, 703. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; He, C.J.; Ji, P.Y.; Zhuo, Z.Y.; Tian, X.Z.; Wang, F.; Tan, D.X.; Liu, G.S. Effects of melatonin on the proliferation and apoptosis of sheep granulosa cells under thermal stress. Int. J. Mol. Sci. 2014, 15, 21090–21104. [Google Scholar] [CrossRef] [Green Version]
- Karck, U.; Keck, C. Physiology of ovarian function. Ther. Umsch. 2002, 59, 153–158. [Google Scholar] [CrossRef]
- Sherwood, N.M.; Adams, B.A.; Isaac, E.R.; Wu, S.; Fradinger, E.A. Knocked down and out: PACAP in development, reproduction and feeding. Peptides 2007, 28, 1680–1687. [Google Scholar] [CrossRef] [PubMed]
- Levy, G.; David, D.; Degani, G. Effect of environmental temperature on growth- and reproduction-related hormones gene expression in the female blue gourami (Trichogaster trichopterus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 160, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Lazarovici, P.; Fink, D., Jr. Heterologous upregulation of nerve growth factor-TrkA receptors in PC12 cells by pituitary adenylate cyclase-activating polypeptide (PACAP). Mol. Cell Biol. Res. Commun. 1999, 2, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Jiang, X.M.; Ganea, D. The neuropeptides VIP and PACAP inhibit IL-2 transcription by decreasing c-Jun and increasing JunB expression in T cells. J. Neuroimmunol. 2000, 104, 68–78. [Google Scholar] [CrossRef]
- Srinivasula, S.M.; Gupta, S.; Datta, P.; Zhang, Z.; Hegde, R.; Cheong, N.; Fernandes-Alnemri, T.; Alnemri, E.S. Inhibitor of apoptosis proteins are substrates for the mitochondrial serine protease Omi/HtrA2. J. Biol. Chem. 2003, 278, 31469–31472. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Nakayama, K.; Shuto, Y.; Somogyvari-Vigh, A.; Arimura, A. Testis-specific prohormone convertase PC4 processes the precursor of pituitary adenylate cyclase-activating polypeptide (PACAP). Peptides 1998, 19, 259–268. [Google Scholar] [CrossRef]
- Nilsson, S.F. PACAP-27 and PACAP-38: Vascular effects in the eye and some other tissues in the rabbit. Eur. J. Pharmacol. 1994, 253, 17–25. [Google Scholar] [CrossRef]
- van Landeghem, F.K.; Weiss, T.; Oehmichen, M.; von Deimling, A. Cellular localization of pituitary adenylate cyclase-activating peptide (PACAP) following traumatic brain injury in humans. Acta Neuropathol. 2007, 113, 683–693. [Google Scholar] [CrossRef]
- Johnson, L.N. The regulation of protein phosphorylation. Biochem. Soc. Trans. 2009, 37, 627–641. [Google Scholar] [CrossRef]
- Kim, C.; Cheng, C.Y.; Saldanha, S.A.; Taylor, S.S. PKA-I holoenzyme structure reveals a mechanism for cAMP-dependent activation. Cell 2007, 130, 1032–1043. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.S.; Kim, C.; Cheng, C.Y.; Brown, S.H.; Wu, J.; Kannan, N. Signaling through cAMP and cAMP-dependent protein kinase: Diverse strategies for drug design. Biochim. Biophys. Acta 2008, 1784, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Nishizuka, Y. Intracellular signaling by hydrolysis of phospholipids and activation of protein kinase C. Science 1992, 258, 607–614. [Google Scholar] [CrossRef]
- Grafer, C.M.; Thomas, R.; Lambrakos, L.; Montoya, I.; White, S.; Halvorson, L.M. GnRH stimulates expression of PACAP in the pituitary gonadotropes via both the PKA and PKC signaling systems. Mol. Endocrinol. 2009, 23, 1022–1032. [Google Scholar] [CrossRef] [Green Version]
- Buteau, H.; Pezet, A.; Ferrag, F.; Perrot-Applanat, M.; Kelly, P.A.; Edery, M. N-Glycosylation of the Prolactin Receptor Is Not Required for Activation of Gene Transcription but Is Crucial for Its Cell Surface Targeting. Mol. Endocrinol. 1998, 12, 544–555. [Google Scholar] [CrossRef]
- Li, L.B.; Chen, N.; Ramamoorthy, S.; Chi, L.; Cui, X.N.; Wang, L.C.; Reith, M.E. The role of N-glycosylation in function and surface trafficking of the human dopamine transporter. J. Biol. Chem. 2004, 279, 21012–21020. [Google Scholar] [CrossRef] [Green Version]
- Langer, I.; Jeandriens, J.; Couvineau, A.; Sanmukh, S.; Latek, D. Signal Transduction by VIP and PACAP Receptors. Biomedicines 2022, 10, 406. [Google Scholar] [CrossRef] [PubMed]
- Kanasaki, H.; Purwana, I.N.; Miyazaki, K. Possible role of PACAP and its PAC1 receptor in the differential regulation of pituitary LHbeta- and FSHbeta-subunit gene expression by pulsatile GnRH stimulation. Biol. Reprod. 2013, 88, 35. [Google Scholar] [CrossRef] [Green Version]
- McGee, E.A.; Perlas, E.; LaPolt, P.S.; Tsafriri, A.; Hsueh, A.J. Follicle-stimulating hormone enhances the development of preantral follicles in juvenile rats. Biol. Reprod. 1997, 57, 990–998. [Google Scholar] [CrossRef] [Green Version]
- Hartshorne, G.M.; Sargent, I.L.; Barlow, D.H. Meiotic progression of mouse oocytes throughout follicle growth and ovulation in vitro. Hum. Reprod. 1994, 9, 352–359. [Google Scholar] [CrossRef]
- Kol, S.; Adashi, E.Y. Intraovarian factors regulating ovarian function. Curr. Opin. Obstet. Gynecol. 1995, 7, 209–213. [Google Scholar] [CrossRef]
- Hillier, S.G. Current concepts of the roles of follicle stimulating hormone and luteinizing hormone in folliculogenesis. Hum. Reprod. 1994, 9, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J.; Downs, S.M. The effect of hypoxanthine on mouse oocyte growth and development in vitro: Maintenance of meiotic arrest and gonadotropin-induced oocyte maturation. Dev. Biol. 1987, 119, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Sirard, M.A. Protein kinases influence bovine oocyte competence during short-term treatment with recombinant human follicle stimulating hormone. Reproduction 2005, 130, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Salomon, A.K.; Leon, K.; Campbell, M.M.; Young, K.A. Folliculogenic factors in photoregressed ovaries: Differences in mRNA expression in early compared to late follicle development. Gen. Comp. Endocrinol. 2018, 260, 90–99. [Google Scholar] [CrossRef]
- Abel, M.H.; Wootton, A.N.; Wilkins, V.; Huhtaniemi, I.; Knight, P.G.; Charlton, H.M. The effect of a null mutation in the follicle-stimulating hormone receptor gene on mouse reproduction. Endocrinology 2000, 141, 1795–1803. [Google Scholar] [CrossRef]
- Latini, S.; Chiarpotto, M.; Muciaccia, B.; Vaccari, S.; Barberi, M.; Guglielmo, M.C.; Stefanini, M.; Cecconi, S.; Canipari, R. Inhibitory effect of pituitary adenylate cyclase activating polypeptide on the initial stages of rat follicle development. Mol. Cell. Endocrinol. 2010, 320, 34–44. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Post-Translational Modification Site | Modified Position | Amino Acid Sequence |
|---|---|---|
| PKA/PKG phosphorylation sites | 131–134 | KRHS |
| PKC phosphorylation sites | 101–103 | STR |
| 130–132 | TKR | |
| N-cardamom acylation site | 6–11 | GARLAL |
| 63–68 | GSPASA | |
| 113–118 | GLGENL | |
| 119–124 | GAGAAD | |
| amidation sites | 159–162 | LGKR |
| 170–173 | KGRR |
| Post-Translational Modification Site | Modified Position | Amino Acid Sequence |
|---|---|---|
| N-glycosylation site | 47–50 | NESS |
| 59–62 | NITC | |
| 116–119 | NCTE | |
| 299–302 | NDST | |
| 342–345 | NESS | |
| 374–377 | NVSK | |
| PKC phosphorylation sites | 151–153 | SVK |
| 376–378 | SKR | |
| 416–418 | SWK | |
| Tyrosine kinase II phosphorylation site | 75–78 | SCPE |
| 93–96 | TIGD | |
| 221–224 | STVE | |
| 376–379 | SKRE | |
| N-cardamom acylation site | 45–50 | GLNESS |
| 55–60 | GMWDNI | |
| 340–345 | GGNESS | |
| 362–367 | GIHYTV | |
| 388–393 | GSFQGF | |
| 438–443 | GVNGGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Y.; Xue, H.; Xu, J.; Wu, M.; Chen, L.; Xu, L. Effect of PACAP/PAC1R on Follicle Development of Djungarian Hamster (Phodopus sungorus) with the Variation of Ambient Temperatures. Biology 2023, 12, 315. https://doi.org/10.3390/biology12020315
Qi Y, Xue H, Xu J, Wu M, Chen L, Xu L. Effect of PACAP/PAC1R on Follicle Development of Djungarian Hamster (Phodopus sungorus) with the Variation of Ambient Temperatures. Biology. 2023; 12(2):315. https://doi.org/10.3390/biology12020315
Chicago/Turabian StyleQi, Yan, Huiliang Xue, Jinhui Xu, Ming Wu, Lei Chen, and Laixiang Xu. 2023. "Effect of PACAP/PAC1R on Follicle Development of Djungarian Hamster (Phodopus sungorus) with the Variation of Ambient Temperatures" Biology 12, no. 2: 315. https://doi.org/10.3390/biology12020315